
Zebrafish HERC7c Acts as an Inhibitor of Fish IFN Response

Abstract
:1. Introduction
2. Results
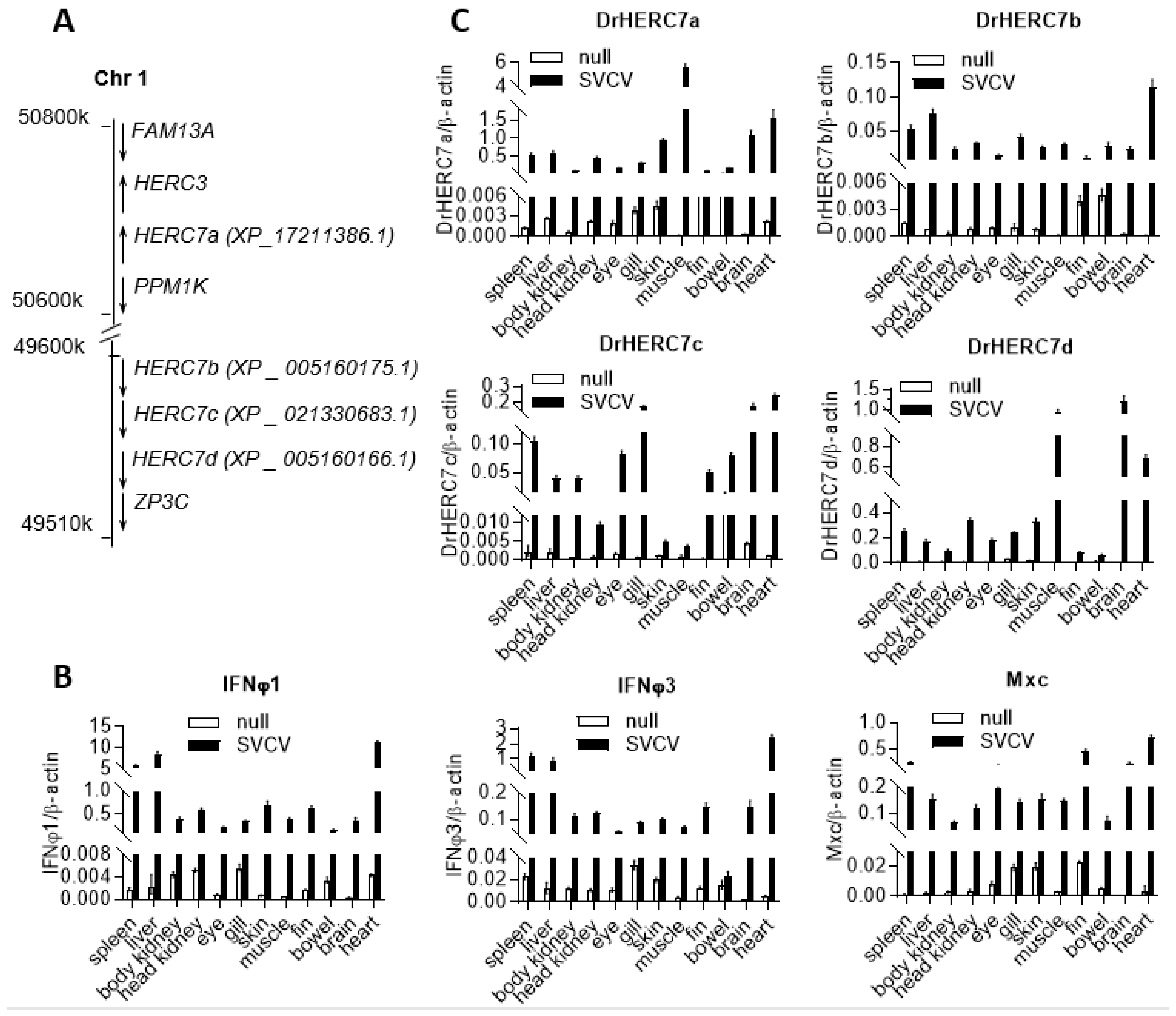
2.1. Zebrafish Has Four herc7 Genes That Are Induced by SVCV Infection
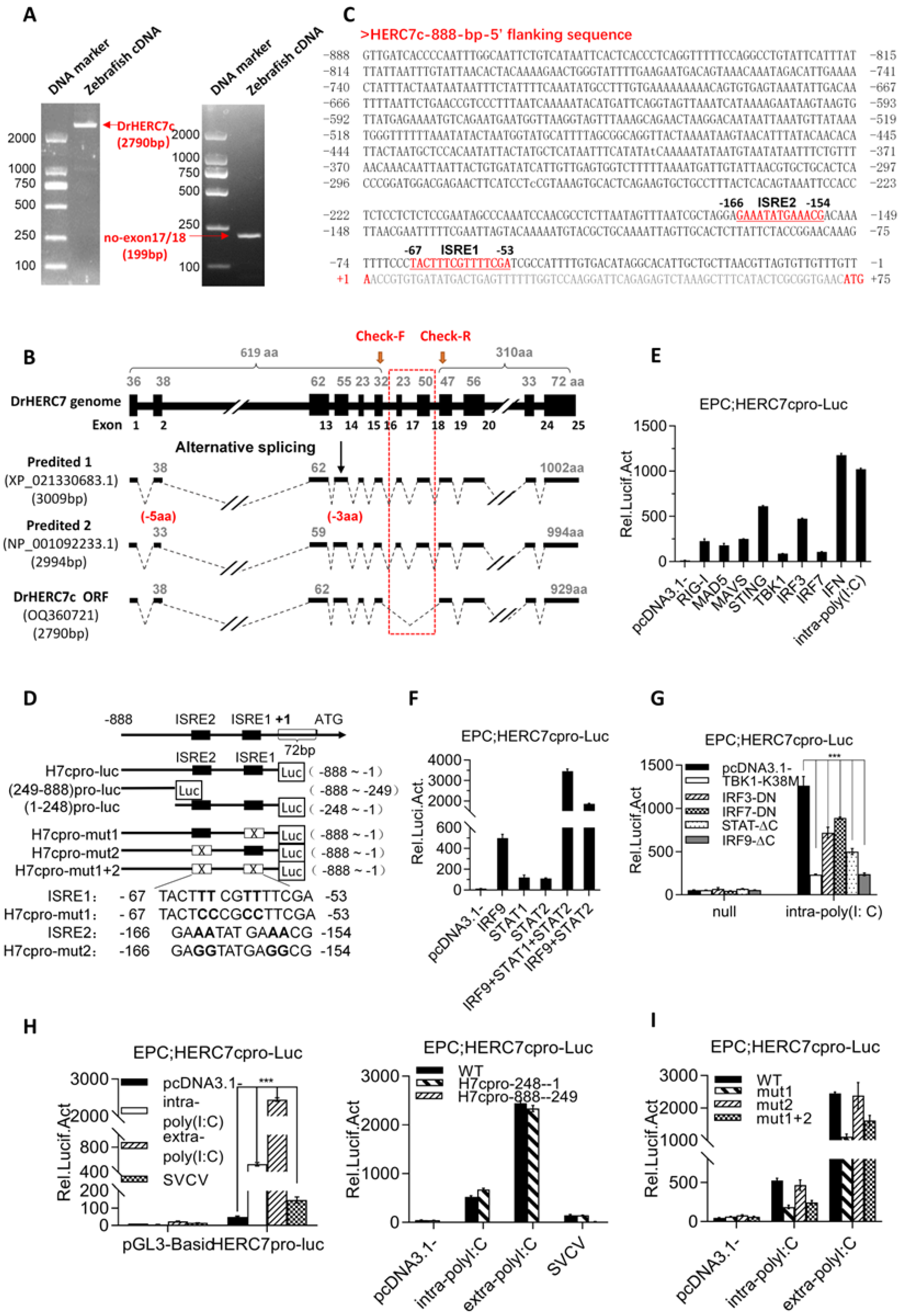
2.2. Zebrafish herc7c Is a Typical IFN-Stimulated Gene
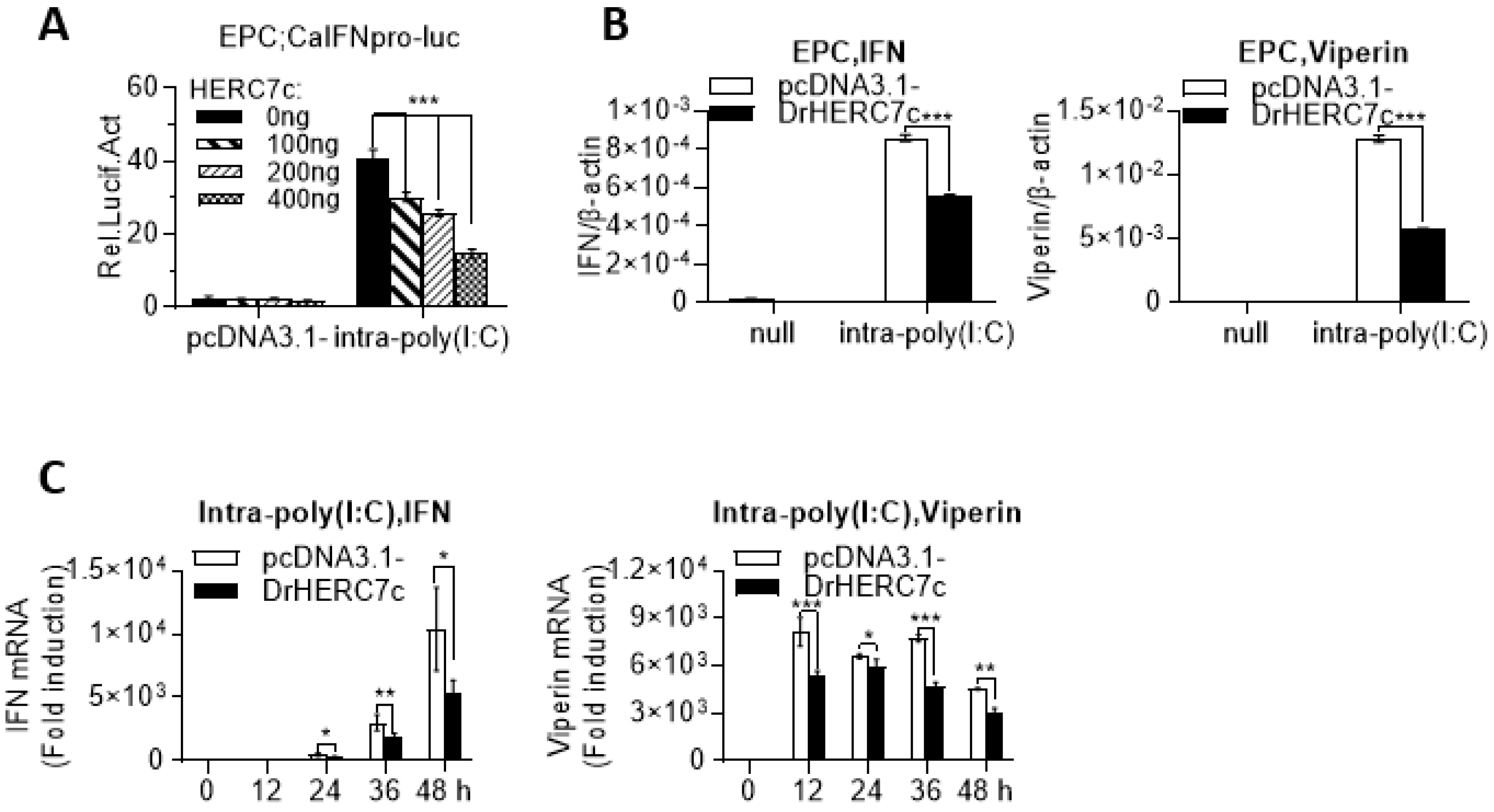
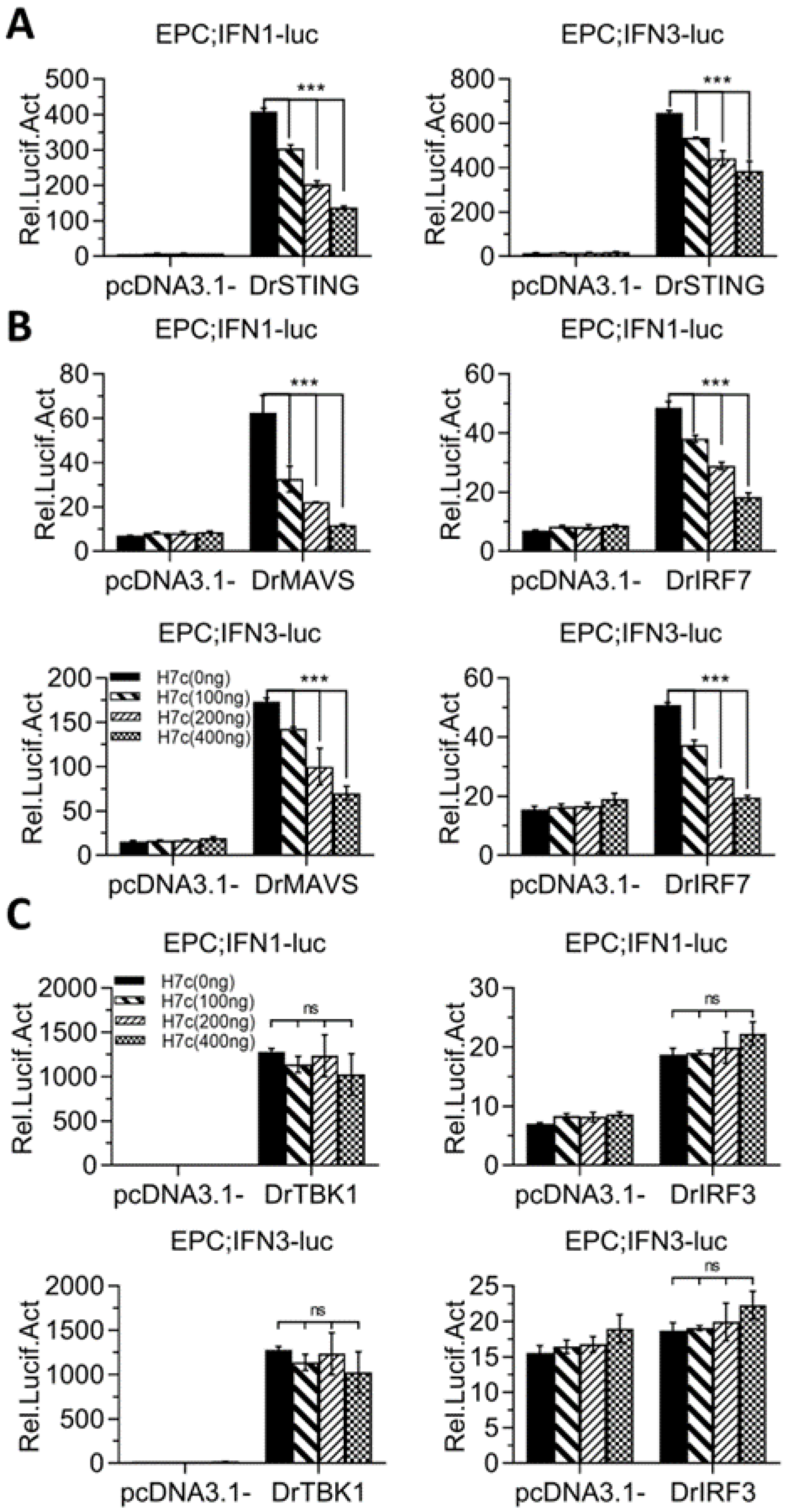
2.3. Zebrafish HERC7c Inhibits Fish IFN Response
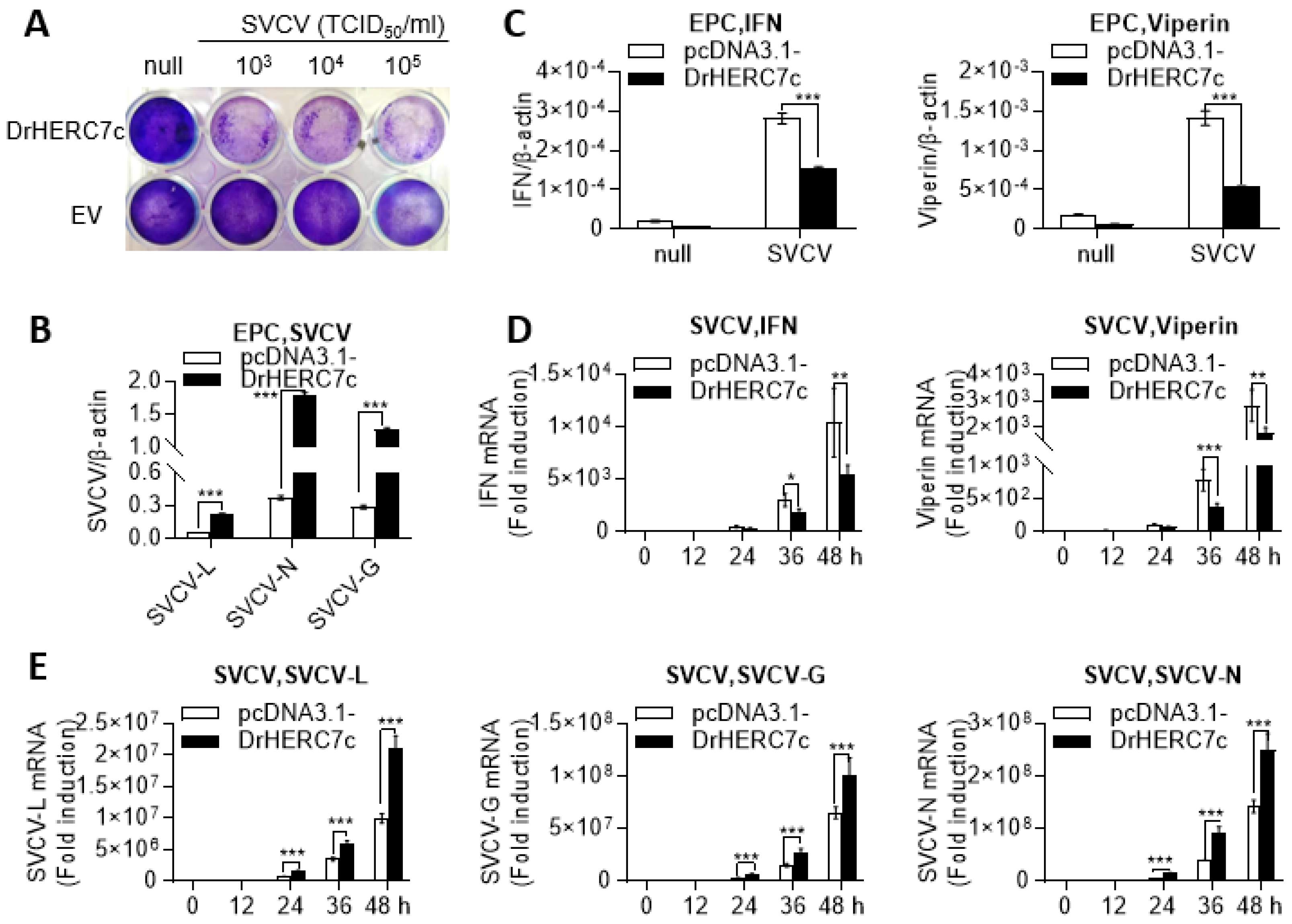
2.4. Zebrafish HERC7c Promotes Virus Replication in Fish Cells
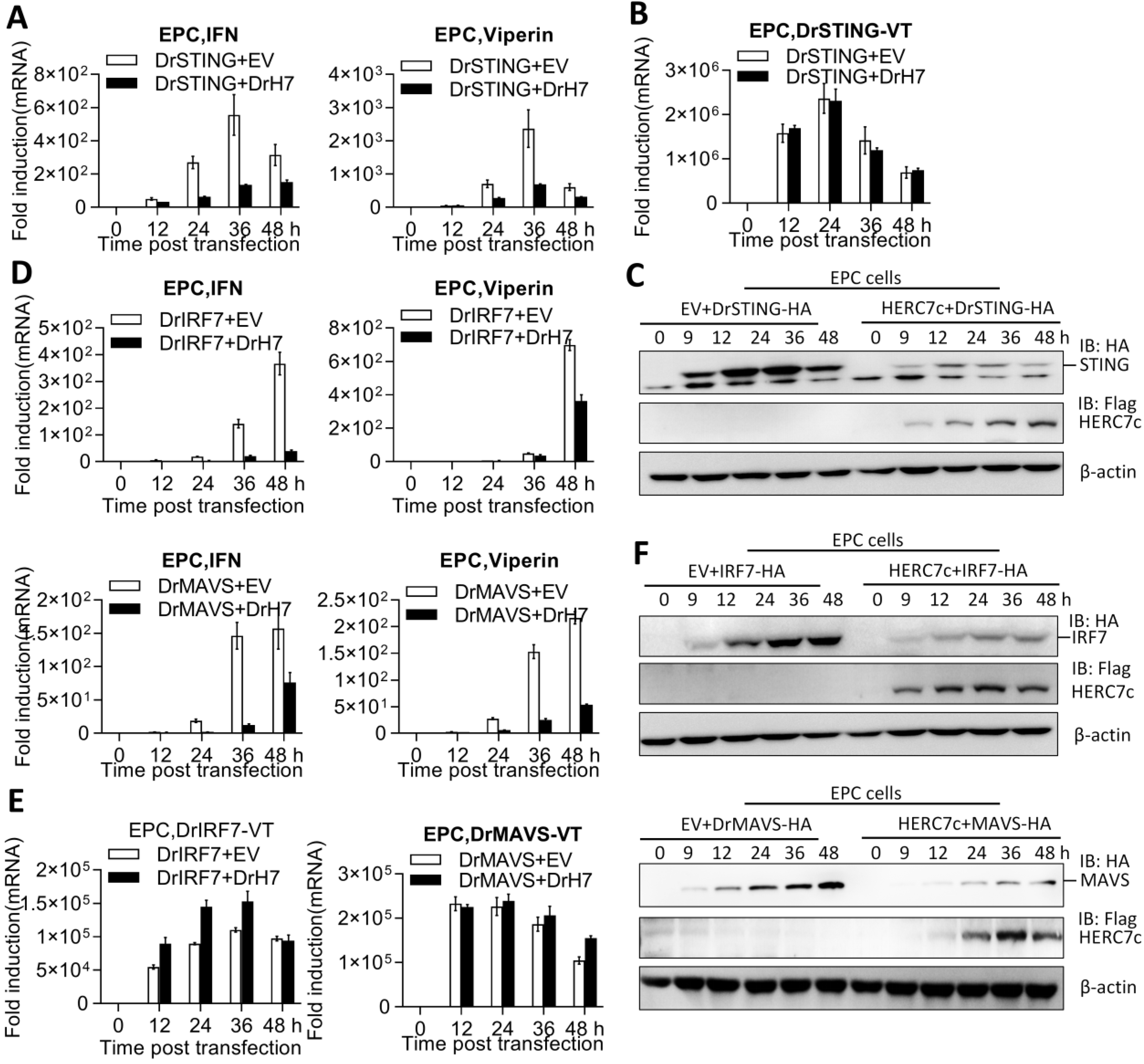
2.5. Zebrafish HERC7c Downregulates Fish IFN Response by Targeting STING, MAVS, and IRF7
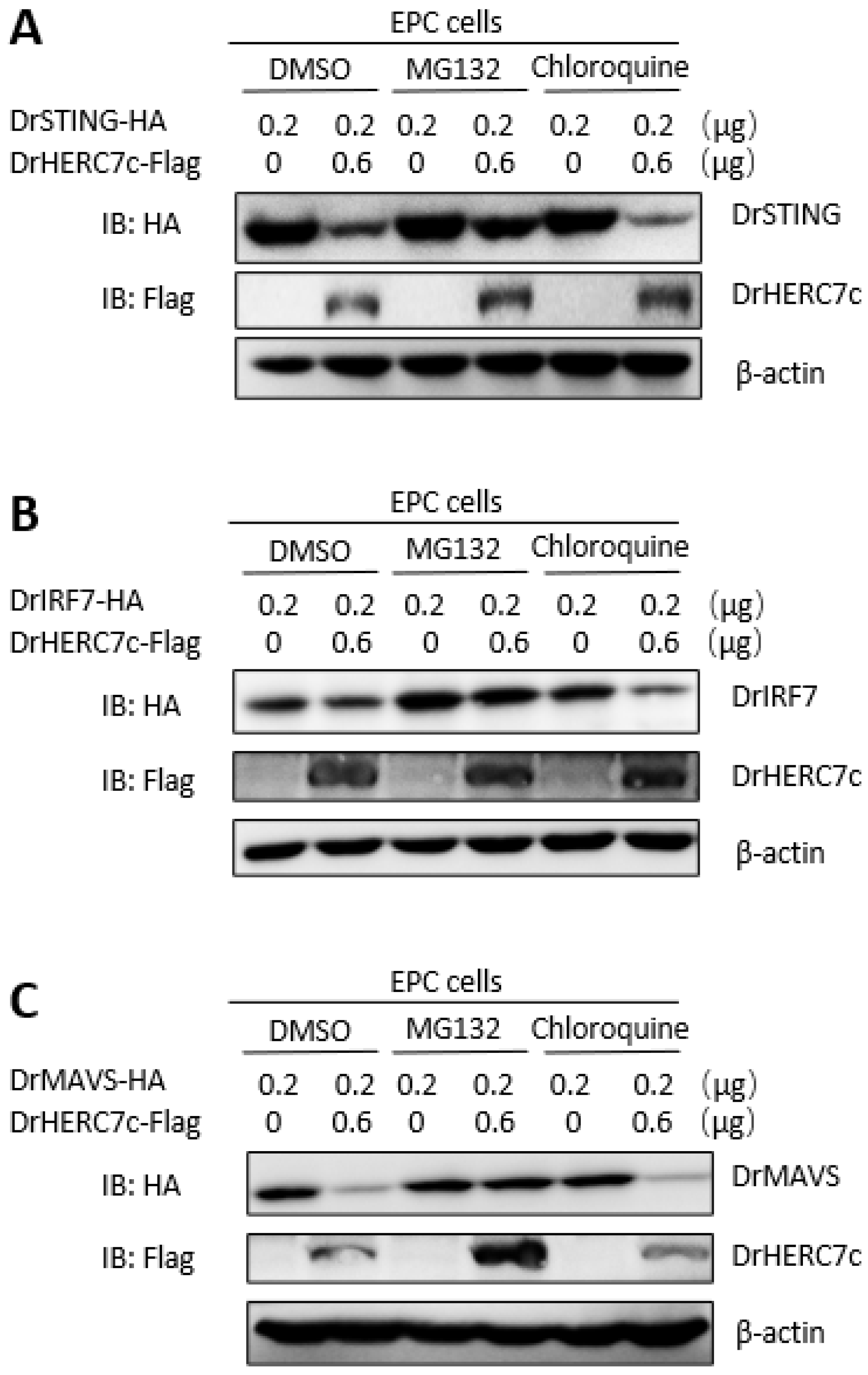
2.6. Zebrafish HERC7c Facilitates STING, MAVS, and IRF7 Protein Degradation to Downregulate IFN Response
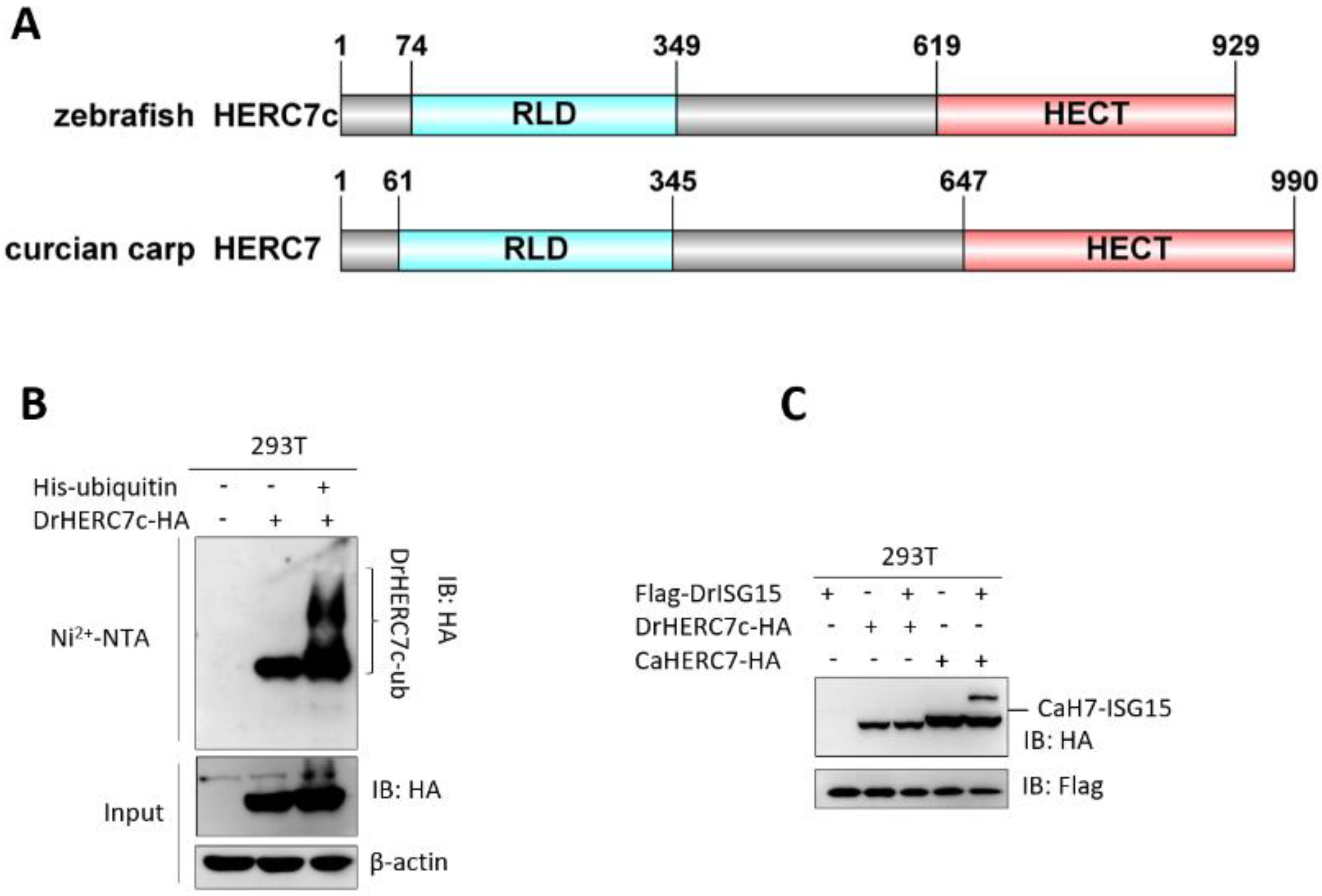
2.7. Zebrafish HERC7c Is an E3 Ligase for the Conjugation of Ubiquitin but Not ISG15
2.8. Zebrafish HERC7c Is a Species-Specific Gene
3. Discussion
4. Materials and Methods
4.1. Cells, Virus, and Zebrafish
4.2. Gene Cloning, Database Mining, and Sequence Analysis
4.3. Plasmid Construction
4.4. Transfection and Luciferase Activity Assays
4.5. RNA Extraction, cDNA Synthesis, and Real Time-PCR
4.6. Ubiquitination Assays and ISGylation Assays
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paparisto, E.; Woods, M.W.; Coleman, M.D.; Moghadasi, S.A.; Kochar, D.S.; Tom, S.K.; Kohio, H.P.; Gibson, R.M.; Rohringer, T.J.; Hunt, N.R.; et al. Evolution-Guided Structural and Functional Analyses of the HERC Family Reveal an Ancient Marine Origin and Determinants of Antiviral Activity. J. Virol. 2018, 92, e00528-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hochrainer, K.; Mayer, H.; Baranyi, U.; Binder, B.; Lipp, J.; Kroismayr, R. The human HERC family of ubiquitin ligases: Novel members, genomic organization, expression profiling, and evolutionary aspects. Genomics 2005, 85, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Marin, I. Animal HECT ubiquitin ligases: Evolution and functional implications. BMC Evol. Biol. 2010, 10, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, S.E.; Rosa, J.L.; Scheffner, M. Characterization of human hect domain family members and their interaction with UbcH5 and UbcH7. J. Biol. Chem. 1998, 273, 12148–12154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, C.; Ventura, F.; Bartrons, R.; Rosa, J.L. HERC3 binding to and regulation by ubiquitin. FEBS Lett. 2001, 488, 74–80. [Google Scholar] [CrossRef]
- Kroismayr, R.; Baranyi, U.; Stehlik, C.; Dorfleutner, A.; Binder, B.R.; Lipp, J. HERC5, a HECT E3 ubiquitin ligase tightly regulated in LPS activated endothelial cells. J. Cell. Sci. 2004, 117 Pt 20, 4749–4756. [Google Scholar] [CrossRef] [Green Version]
- Dastur, A.; Beaudenon, S.; Kelley, M.; Krug, R.M.; Huibregtse, J.M. Herc5, an interferon-induced HECT E3 enzyme, is required for conjugation of ISG15 in human cells. J. Biol. Chem. 2006, 281, 4334–4338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, J.J.Y.; Pung, Y.F.; Sze, N.S.K.; Chin, K.C. HERC5 is an IFN-induced HECT-type E3 protein ligase that mediates type I IFN-induced ISGylation of protein targets. Proc. Natl. Acad. Sci. USA 2006, 103, 10735–10740. [Google Scholar] [CrossRef] [Green Version]
- Sala-Gaston, J.; Martinez-Martinez, A.; Pedrazza, L.; Lorenzo-Martin, L.F.; Caloto, R.; Bustelo, X.R.; Ventura, F.; Rosa, J.L. HERC Ubiquitin Ligases in Cancer. Cancers 2020, 12, 1653. [Google Scholar] [CrossRef]
- Garcia-Gonzalo, F.R.; Rosa, J.L. The HERC proteins: Functional and evolutionary insights. Cell. Mol. Life Sci. 2005, 62, 1826–1838. [Google Scholar] [CrossRef]
- Oudshoorn, D.; van Boheemen, S.; Sanchez-Aparicio, M.T.; Rajsbaum, R.; Garcia-Sastre, A.; Versteeg, G.A. HERC6 Is the Main E3 Ligase for Global ISG15 Conjugation in Mouse Cells. PLoS ONE 2012, 7, e29870. [Google Scholar] [CrossRef] [Green Version]
- Rehwinkel, J.; Gack, M.U. RIG-I-like receptors: Their regulation and roles in RNA sensing. Nat. Rev. Immunol. 2020, 20, 537–551. [Google Scholar] [CrossRef]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O’Garra, A. Type I interferons in infectious disease. Nat. Rev. Immunol. 2015, 15, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Zhao, X.; Gong, X.Y.; Wang, Y.; Gui, J.F.; Zhang, Y.B. FTRCA1, a Species-Specific Member of finTRIM Family, Negatively Regulates Fish IFN Response through Autophage-Lysosomal Degradation of TBK1. J. Immunol. 2019, 202, 2407–2420. [Google Scholar] [CrossRef] [PubMed]
- Qu, Z.L.; Li, Y.L.; Gong, X.Y.; Zhao, X.; Sun, H.Y.; Dan, C.; Gui, J.F.; Zhang, Y.B. A finTRIM Family Protein Acquires RNA-Binding Activity and E3 Ligase Activity to Shape the IFN Response in Fish. J. Immunol. 2022, 209, 1335–1347. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.Y.; Zhang, Q.M.; Zhao, X.; Li, Y.L.; Qu, Z.L.; Li, Z.; Dan, C.; Gui, J.F.; Zhang, Y.B. LGP2 is essential for zebrafish survival through dual regulation of IFN antiviral response. iScience 2022, 25, 104821. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.X.; Yang, K.; Liu, X.; Liu, X.Y.; Wei, B.; Shan, Y.F.; Zhu, L.H.; Wang, C. Positive regulation of interferon regulatory factor 3 activation by Herc5 via ISG15 modification. Mol. Cell Biol. 2010, 30, 2424–2436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacquet, S.; Pontier, D.; Etienne, L. Rapid Evolution of HERC6 and Duplication of a Chimeric HERC5/6 Gene in Rodents and Bats Suggest an Overlooked Role of HERCs in Mammalian Immunity. Front. Immunol. 2020, 11, 605270. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.L.; Gong, X.Y.; Qu, Z.L.; Zhao, X.; Dan, C.; Gui, J.F.; Zhang, Y.B. A Novel Non-Mammalian-Specific HERC7 Negatively Regulates IFN Response through Degrading RLR Signaling Factors. J. Immunol. 2022, 208, 1189–1203. [Google Scholar] [CrossRef]
- Zhang, Y.-B.; Li, Q.; Gui, J.-F. Differential expression of two Carassius auratus Mx genes in cultured CAB cells induced by grass carp hemorrhage virus and interferon. Immunogenetics 2004, 56, 68–75. [Google Scholar] [CrossRef]
- Yu, F.-F.; Zhang, Y.-B.; Liu, T.-K.; Liu, Y.; Sun, F.; Jiang, J.; Gui, J.-F. Fish virus-induced interferon exerts antiviral function through Stat1 pathway. Mol. Immunol. 2010, 47, 2330–2341. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhang, Y.B.; Liu, T.K.; Sun, F.; Gui, J.F. Subcellular localization and functional characterization of a fish IRF9 from crucian carp Carassius auratus. Fish Shellfish. Immunol. 2012, 33, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.B.; Gui, J.F. Molecular regulation of interferon antiviral response in fish. Dev. Comp. Immunol. 2012, 38, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Zhang, Y.B.; Liu, T.K.; Shi, J.; Wang, B.; Gui, J.F. Fish MITA serves as a mediator for distinct fish IFN gene activation dependent on IRF3 or IRF7. J. Immunol. 2011, 187, 2531–2539. [Google Scholar] [CrossRef] [Green Version]
- Gong, X.Y.; Zhang, Q.M.; Gui, J.F.; Zhang, Y.B. SVCV infection triggers fish IFN response through RLR signaling pathway. Fish Shellfish. Immunol. 2019, 86, 1058–1063. [Google Scholar] [CrossRef]
- Rosa, J.L.; Casaroli-Marano, R.P.; Buckler, A.J.; Vilaro, S.; Barbacid, M. p619, a giant protein related to the chromosome condensation regulator RCC1, stimulates guanine nucleotide exchange on ARF1 and Rab proteins. EMBO J. 1996, 15, 4262–4273. [Google Scholar] [CrossRef]
- Jaillon, O.; Aury, J.-M.; Brunet, F.; Petit, J.-L.; Stange-Thomann, N.; Mauceli, E.; Bouneau, L.; Fischer, C.; Ozouf-Costaz, C.; Bernot, A.; et al. Genome duplication in the teleost fish Tetraodon nigroviridis reveals the early vertebrate proto-karyotype. Nature 2004, 431, 946–957. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, X.Y.; Xu, W.J.; Wang, K.; Wu, B.; Xu, M.; Chen, Y.; Miao, L.J.; Wang, Z.W.; Li, Z.; et al. Comparative genome anatomy reveals evolutionary insights into a unique amphitriploid fish. Nat. Ecol. Evol. 2022, 6, 1354–1366. [Google Scholar] [CrossRef]
- Gui, J.F.; Zhou, L.; Li, X.Y. Rethinking fish biology and biotechnologies in the challenge era for burgeoning genome resources and strengthening food security. Water Biol. Secur. 2022, 1, 100002. [Google Scholar] [CrossRef]
- Feng, H.; Zhang, Q.M.; Zhang, Y.B.; Li, Z.; Zhang, J.; Xiong, Y.W.; Wu, M.; Gui, J.F. Zebrafish IRF1, IRF3, and IRF7 Differentially Regulate IFNPhi1 and IFNPhi3 Expression through Assembly of Homo- or Heteroprotein Complexes. J. Immunol. 2016, 197, 1893–1904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, L.L.; Zhao, X.; Gong, X.Y.; Li, Y.L.; Qu, Z.L.; Sun, H.Y.; Guo, W.H.; Dan, C.; Gui, J.F.; Zhang, Y.B. Promoter Binding and Nuclear Retention Features of Zebrafish IRF Family Members in IFN Response. Front. Immunol. 2022, 13, 861262. [Google Scholar] [CrossRef]
- Au, W.C.; Moore, P.A.; Lowther, W.; Juang, Y.T.; Pitha, P.M. Identification of a Member of the Interferon Regulatory Factor Family That Binds to the Interferon-Stimulated Response Element and Activates Expression of Interferon-Induced Genes. Proc. Natl. Acad. Sci. USA 1995, 92, 11657–11661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, F.; Zhang, Y.B.; Liu, T.K.; Gan, L.; Yu, F.F.; Liu, Y.; Gui, J.F. Characterization of fish IRF3 as an IFN-inducible protein reveals evolving regulation of IFN response in vertebrates. J. Immunol. 2010, 185, 7573–7582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathieu, N.A.; Paparisto, E.; Barr, S.D.; Spratt, D.E. HERC5 and the ISGylation Pathway: Critical Modulators of the Antiviral Immune Response. Viruses 2021, 13, 1102. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Mei, J.; Ge, C.T.; Liu, X.L.; Gui, J.F. Sex determination mechanisms and sex control approaches in aquaculture animals. Sci. China Life Sci. 2022, 65, 1091–1122. [Google Scholar] [CrossRef]
- Gong, X.; Zhang, Y.B. A protocol to characterize zebrafish LGP2 as a dual regulator of IFN response during viral infection. STAR Protoc. 2022, 3, 101844. [Google Scholar] [CrossRef]
- Feng, H.; Zhang, Y.B.; Zhang, Q.M.; Li, Z.; Zhang, Q.Y.; Gui, J.F. Zebrafish IRF1 Regulates IFN Antiviral Response through Binding to IFNvarphi1 and IFNvarphi3 Promoters Downstream of MyD88 Signaling. J. Immunol. 2015, 194, 1225–1238. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.M.; Zhao, X.; Li, Z.; Wu, M.; Gui, J.F.; Zhang, Y.B. Alternative Splicing Transcripts of Zebrafish LGP2 Gene Differentially Contribute to IFN Antiviral Response. J. Immunol. 2018, 200, 688–703. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence | Usage |
|---|---|---|
| DrH7c-F1 | CCGTGTGATATGACTGAG | Amplify herc7c ORF |
| DrH7c-F2 | TAAAGCTTTCATACTCGCGG | |
| DrH7c-R1 | GCAAAGTTTAGCAACAAACTTGAC | |
| DrH7c-R2 | AGGCAAGTTCACCCTCGTCC | |
| Check-F | GTGCTTTGGATGGCATCGCGGC | Check primers |
| Check-R | CCAGTGAACTCTCATGAACTTCC | |
| DrH7c-pro-F | GTTGATCACCCCAATTTGG | Amplify 5′ flanking |
| DrH7c-pro-R | GCAAAGTTTAGCAACAAACTTGAC | |
| DrH7cpro-luc-F | GAGCTCTTACGCGTGGTTGATCACCCCAATTTGG | Construct luciferase plasmid |
| DrH7cpro-luc-R | TCGAGCCCGGGCTAGAACAAACAACACTAACGTTAAGC | |
| DrH7cpro-mut1-F | TACTCCCGCCTTCGA | |
| DrH7cpro-mut1-R | TCGAAGGCGGGAGTA | |
| DrH7cpro-mut2-F | GGAGAGGTATGAGGCGAC | |
| DrH7cpro-mut2-R | GTCGCCTCATACCTCTCC | |
| 3.1-F | TGCTGGATATCTGCAGCCACCGCCACCATG | Construct eukaryotic expression plasmids |
| 7c-FLAG-R | CGAGCTCGGATCGCTTATCGTCGTCATCCTTGTAATCTCGCCCTCGTCCAAAAACAGC | |
| 7c-HA-R | CGAGCTCGGATCGAGCGTAGTCTGGGACGTCGTATGGGTATCGCCCTCGTCCAAAAACAG | |
| EPC-actin-Q-F | CAGATCATGTTTGAGACC | RT-PCR |
| EPC-actin-Q-R | ATTGCCAATGGTGATGAC | |
| EPC-Mx-Q-F | GGCTGGAGCAGGTGTTGGTATC | |
| EPC-Mx-Q-R | TCCACCAGGTCCGGCTTTGTTAA | |
| EPC-IFN-Q-F | ATGAAAACTCAAATGTGGACGTA | |
| EPC-IFN-Q-R | GATAGTTTCCACCCATTTCCTTAA | |
| EPC-Viperin-Q-F | AGCGAGGCTTACGACTTCTG | |
| EPC- Viperin-Q-R | GCACCAACTCTCCCAGAAAA | |
| VT-pcDNA3.1-Q-F | CGACTCACTATAGGGAGACC | |
| VT-DrSTING-Q-R | CCTTGAATGGAAGAGCAATTCCTC | |
| VT-DrMAVS-Q-R | CCCGATCAGAGATTGTGAGGCA | |
| VT-DrIRF7-Q-R | CTCGTTGATCTTGCCGCTGAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.-L.; Gong, X.-Y.; Qu, Z.-L.; Zhao, X.; Dan, C.; Sun, H.-Y.; An, L.-L.; Gui, J.-F.; Zhang, Y.-B. Zebrafish HERC7c Acts as an Inhibitor of Fish IFN Response. Int. J. Mol. Sci. 2023, 24, 4592. https://doi.org/10.3390/ijms24054592
Li Y-L, Gong X-Y, Qu Z-L, Zhao X, Dan C, Sun H-Y, An L-L, Gui J-F, Zhang Y-B. Zebrafish HERC7c Acts as an Inhibitor of Fish IFN Response. International Journal of Molecular Sciences. 2023; 24(5):4592. https://doi.org/10.3390/ijms24054592
Chicago/Turabian StyleLi, Yi-Lin, Xiu-Ying Gong, Zi-Ling Qu, Xiang Zhao, Cheng Dan, Hao-Yu Sun, Li-Li An, Jian-Fang Gui, and Yi-Bing Zhang. 2023. "Zebrafish HERC7c Acts as an Inhibitor of Fish IFN Response" International Journal of Molecular Sciences 24, no. 5: 4592. https://doi.org/10.3390/ijms24054592