
Opportunities and Hurdles to the Adoption and Enhanced Efficacy of Feed Additives towards Pronounced Mitigation of Enteric Methane Emissions from Ruminant Livestock

Abstract
:1. Enteric Methane Emissions and Climate Change
2. Intensification, Productivity, and Enteric Methane Emissions
3. Mitigation of Enteric Methane Emissions
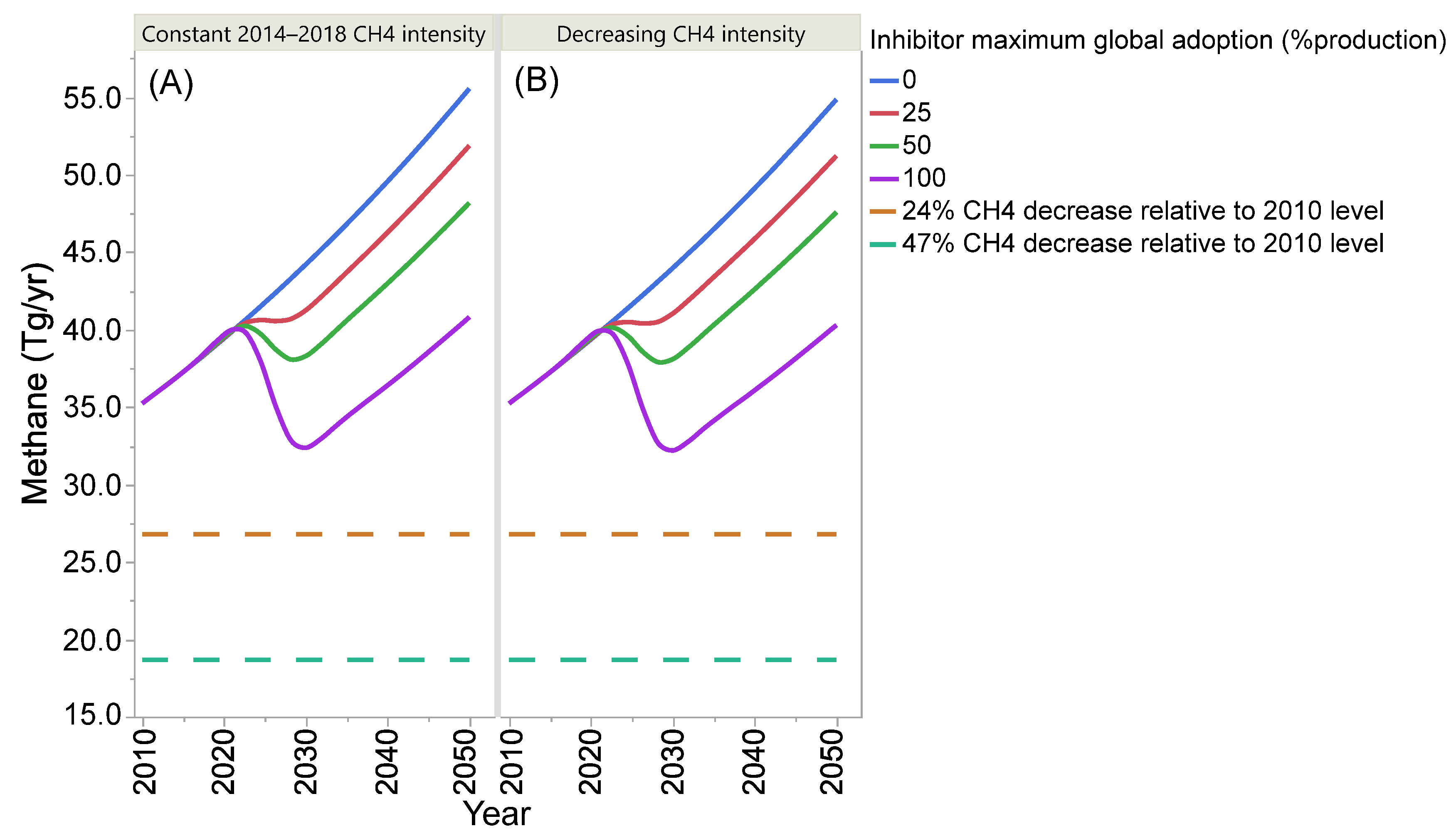
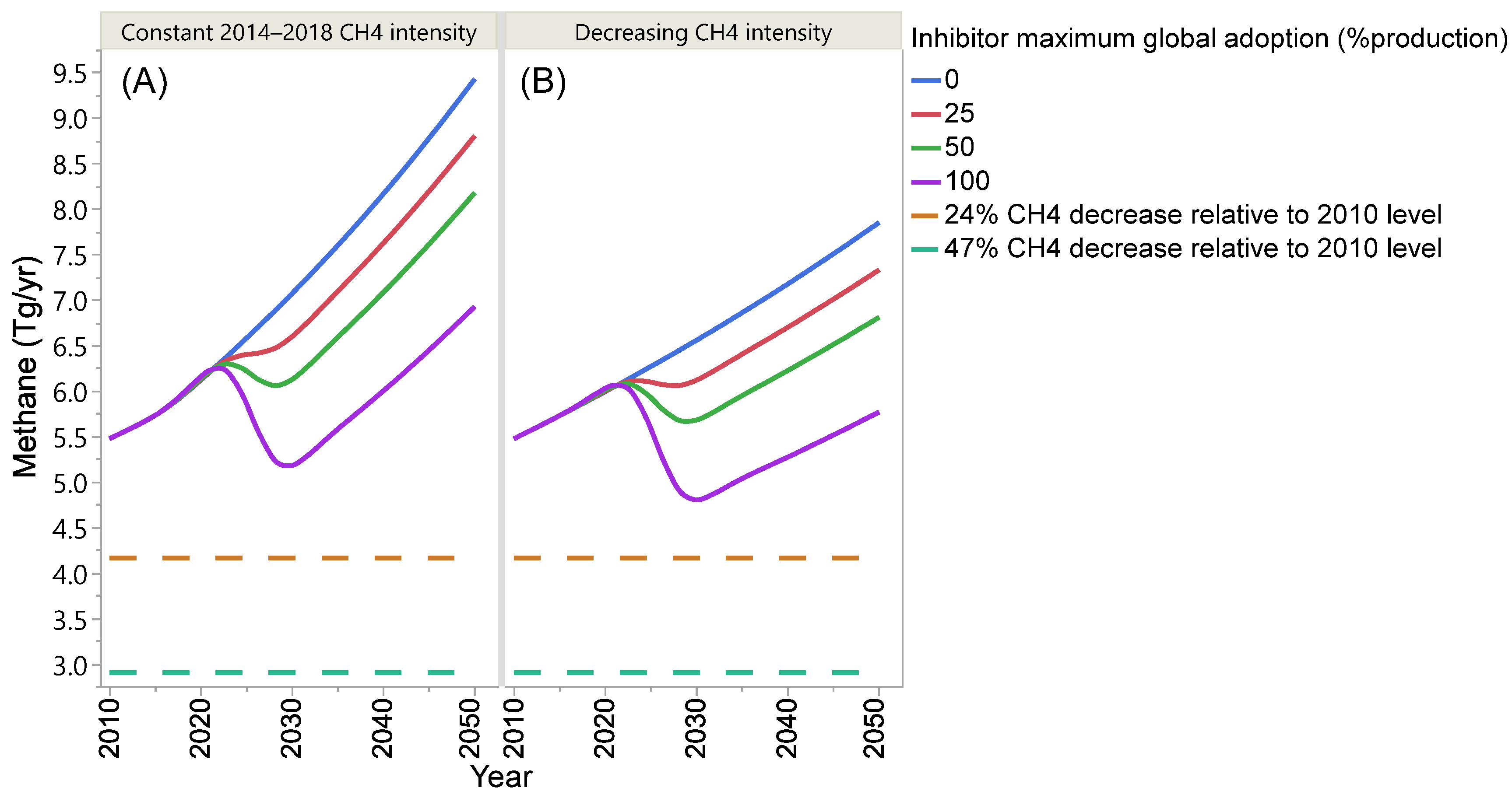
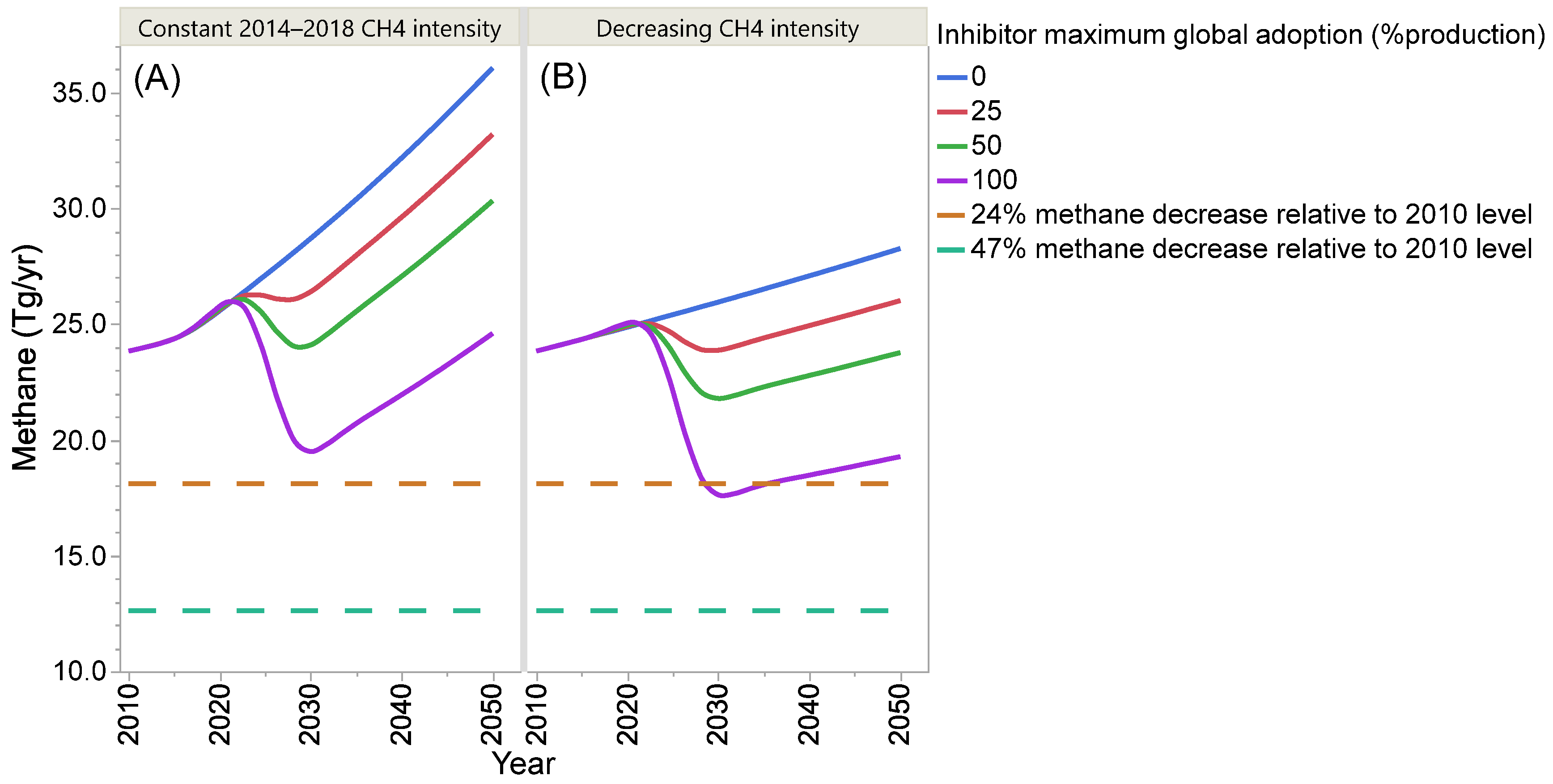
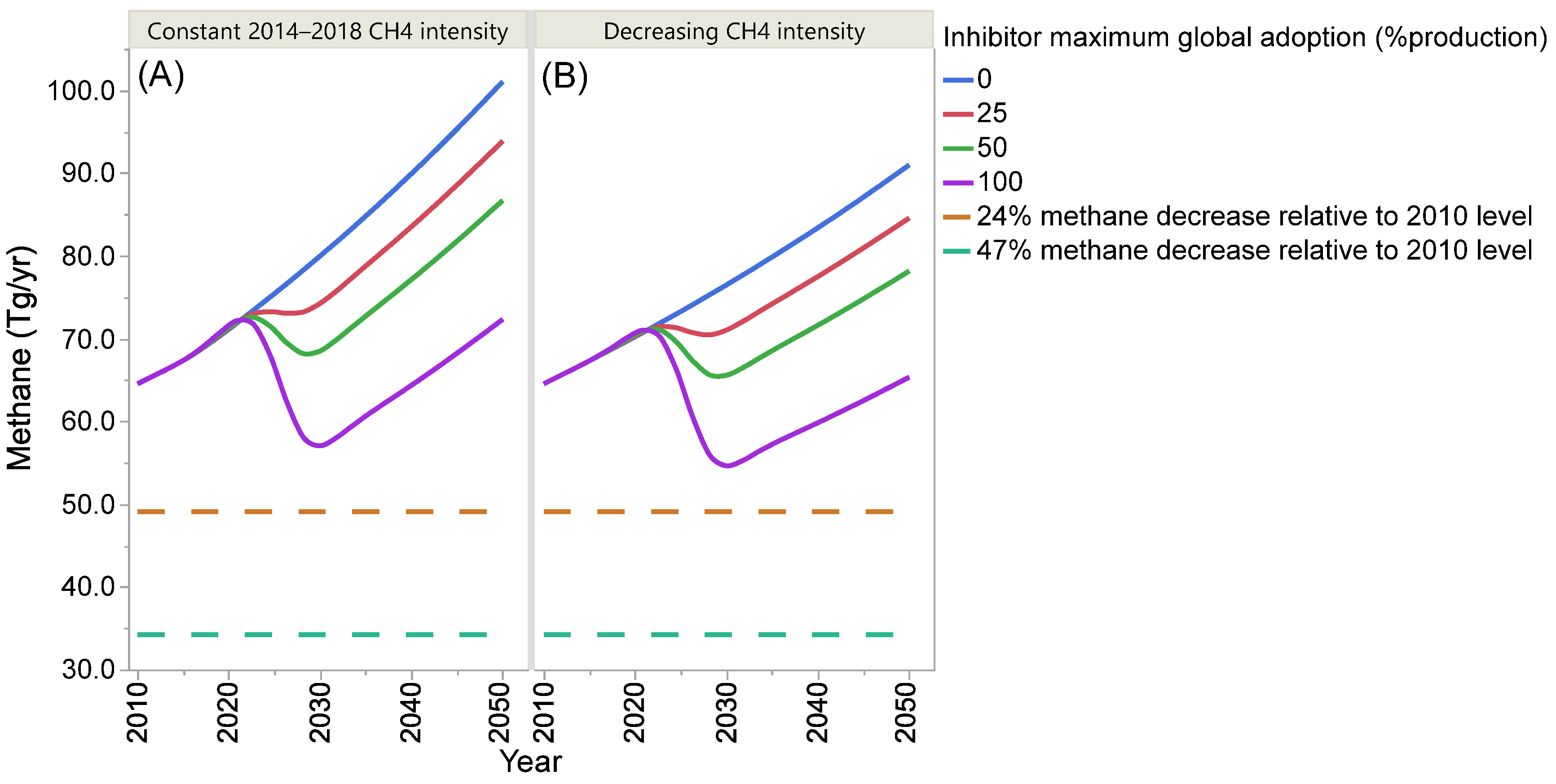
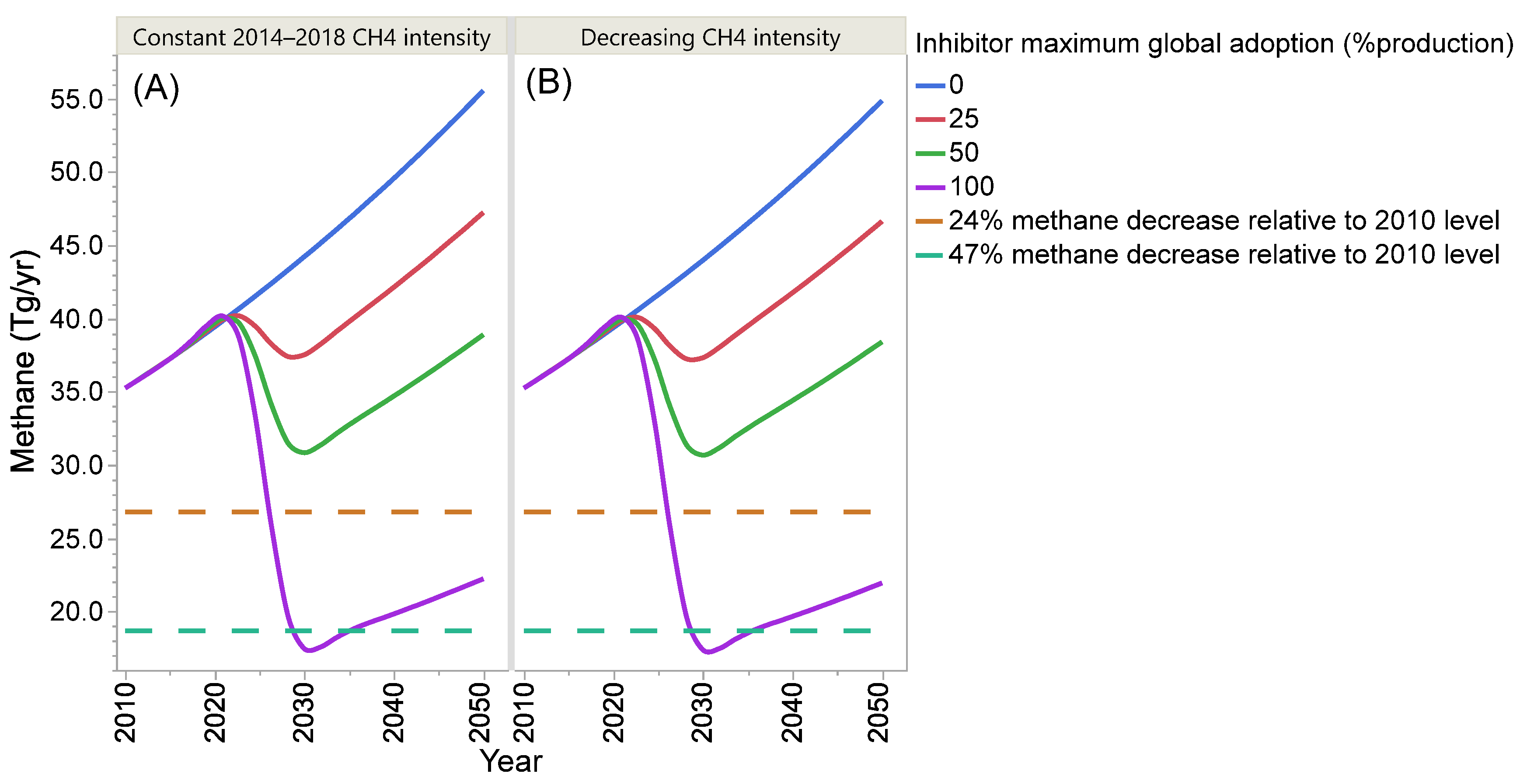
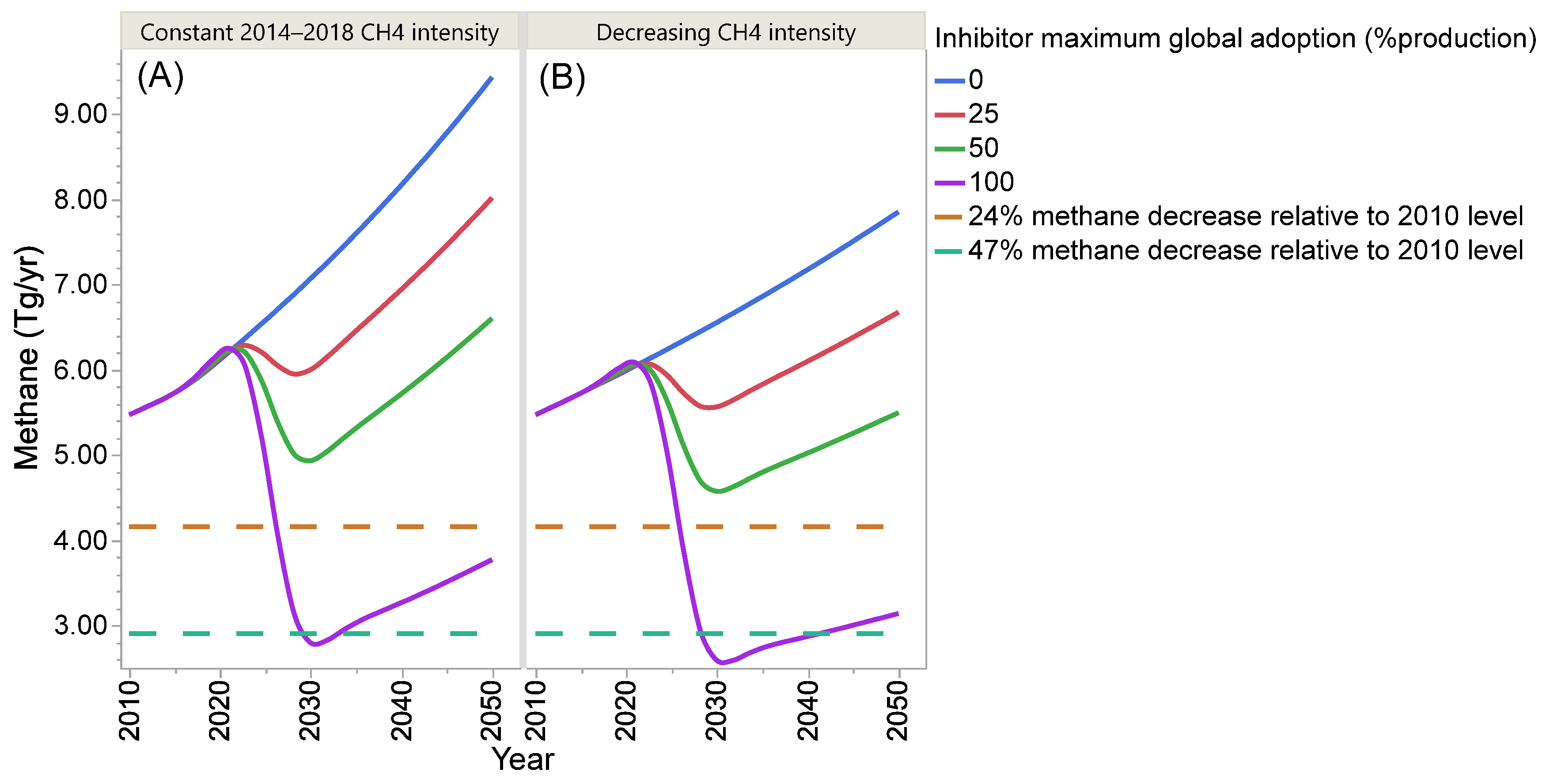
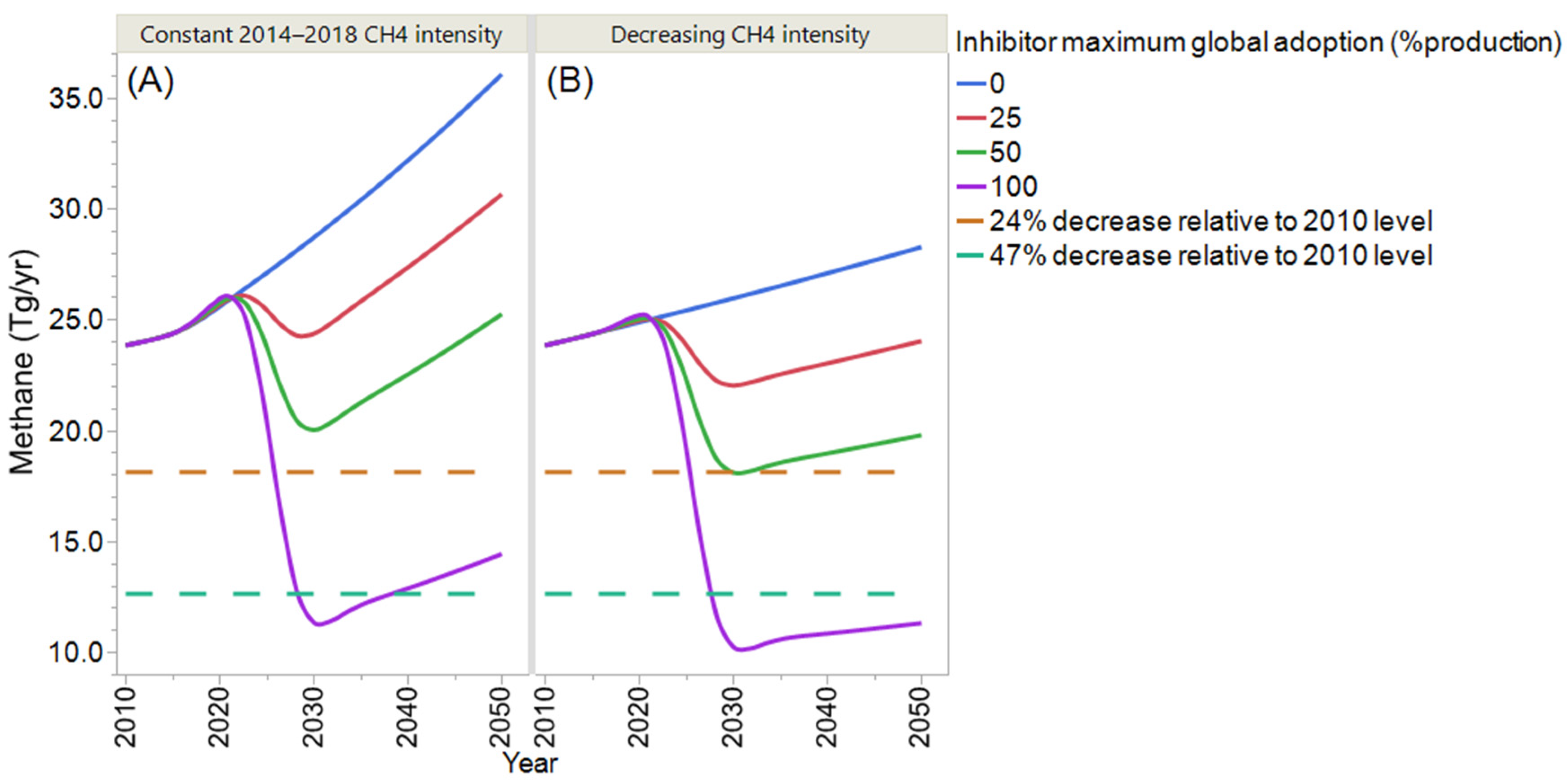
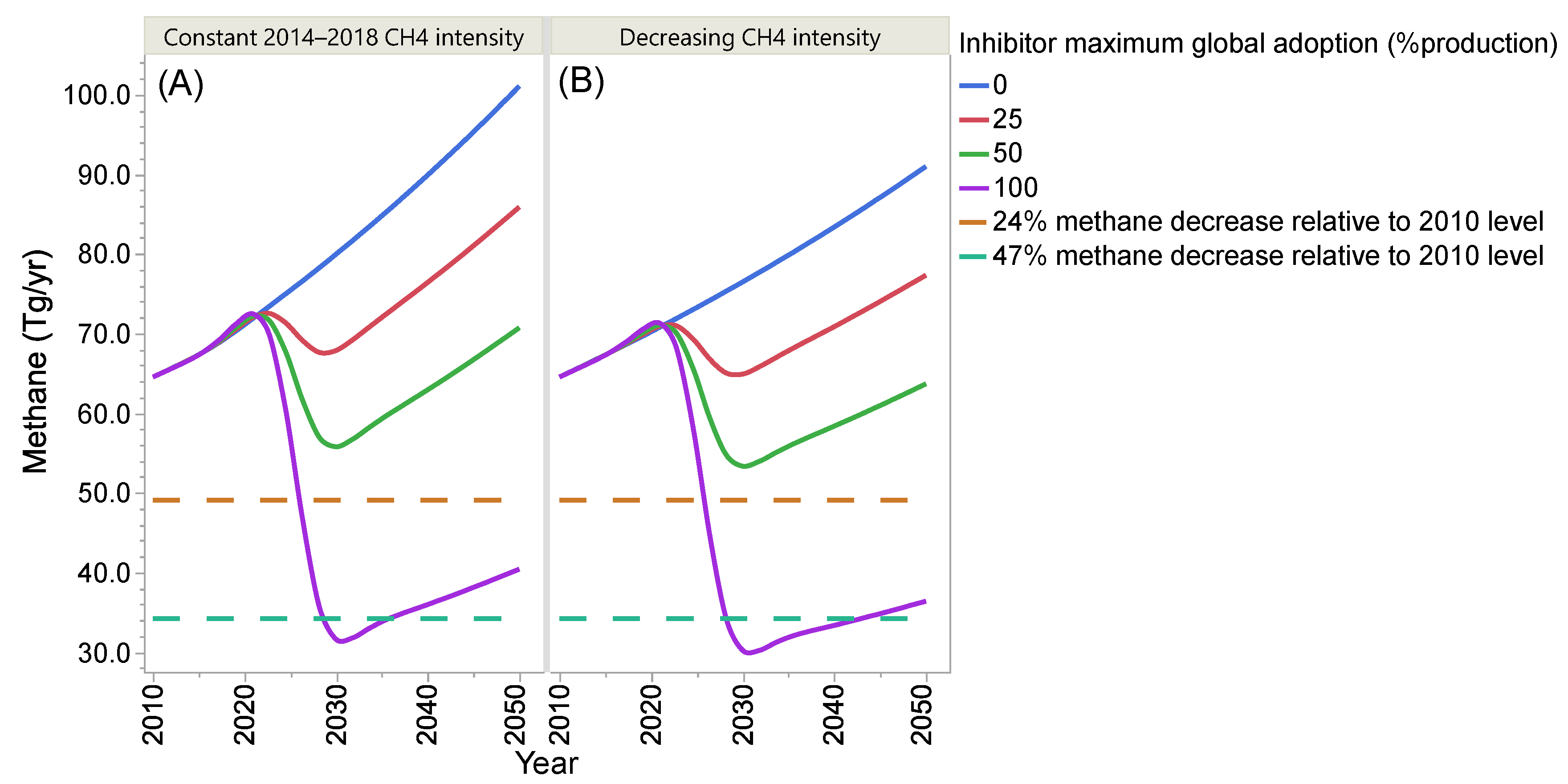
4. Projection of Global Enteric Methane Emission under Different Scenarios of Intensification and Adoption of Inhibitors of Methanogenesis
5. Pronounced Inhibition of Rumen Methanogenesis with Feed Additives
6. Cost Effectiveness and Co-Benefits of Inhibiting Methanogenesis
- Economic incentives;
- Methanogenesis inhibition increasing feed efficiency;
- Adjusting basal diet composition to the inhibition of methanogenesis.
6.1. Economic Incentives
6.2. Methanogenesis Inhibition Increasing Feed Efficiency
6.3. Adjusting Basal Diet Composition to the Inhibition of Methanogenesis
7. Adoption of Inhibitors of Methanogenesis in Grazing Systems
8. Safety and Other Aspects Important for the Adoption of Inhibitors of Methanogenesis
9. Possibilities for Enhancing the Effectiveness of Inhibitors of Methanogenesis
10. Final Remarks
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hoegh-Guldberg, O.; Jacob, D.; Taylor, M.; Bindi, M.; Brown, S.; Camilloni, I.; Diedhiou, A.; Djalante, R.; Ebi, K.L.; Engelbrecht, F.; et al. Impacts of 1.5 °C global warming on natural and human systems. In Global Warming of 1.5 °C; Masson-Delmotte, V., Zhai, P., Pörtner, H.-O., Roberts, D., Skea, J., Shukla, P.R., Pirani, A., Moufouma-Okia, W., Péan, C., Pidcock, R., et al., Eds.; An IPCC Special Report on the Impacts of Global Warming of 1.5 °C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2018; pp. 175–311. [Google Scholar]
- United Nations Environment Programme and Climate and Clean Air Coalition. Global Methane Assessment: Benefits and Costs of Mitigating Methane Emissions; United Nations: Nairobi, Kenia, 2021; 173p.
- IPCC. Summary for policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021; pp. 3–32. [Google Scholar] [CrossRef]
- Szopa, S.; Naik, V.; Adhikary, B.; Artaxo, P.; Berntsen, T.; Collins, W.D.; Fuzzi, S.; Gallardo, L.; Kiendler-Scharr, A.; Klimont, Z.; et al. Short-lived climate forcers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021; pp. 817–922. [Google Scholar] [CrossRef]
- Saunois, M.; Stavert, A.R.; Poulter, B.; Bousquet, P.; Canadell, J.G.; Jackson, R.B.; Raymond, P.A.; Dlugokencky, E.J.; Houweling, S.; Patra, P.K.; et al. The global methane budget 2000–2017. Earth Syst. Sci. Data 2020, 12, 1561–1623. [Google Scholar] [CrossRef]
- Reisinger, A.; Clark, H.; Cowie, A.L.; Emmet-Booth, J.; Gonzalez Fischer, C.; Herrero, M.; Howden, M.; Leahy, S. How necessary and feasible are reductions of methane emissions from livestock to support stringent temperature goals? Philos. Trans. R. Soc. A 2021, 379, 20200452. [Google Scholar] [CrossRef]
- Chang, J.; Peng, S.; Yin, Y.; Ciais, P.; Havlik, P.; Herrero, M. The key role of production efficiency changes in livestock methane emission mitigation. AGU Adv. 2021, 2, e2021AV000391. [Google Scholar] [CrossRef]
- Niu, M.; Kebreab, E.; Hristov, A.N.; Oh, J.; Arndt, C.; Bannink, A.; Bayat, A.R.; Brito, A.F.; Boland, T.; Casper, D.; et al. Prediction of enteric methane production, yield, and intensity in dairy cattle using an intercontinental database. Glob. Chang. Biol. 2018, 24, 3368–3389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capper, J.L.; Cady, R.A.; Bauman, D.E. The environmental impact of dairy production: 1944 compared with 2007. J. Anim. Sci. 2009, 87, 2160–2167. [Google Scholar] [CrossRef]
- Capper, J.L. The environmental impact of beef production in the United States: 1977 compared with 2007. J. Anim. Sci. 2011, 89, 4249–4261. [Google Scholar] [CrossRef] [Green Version]
- Hristov, A.N.; Ott, T.; Tricarico, J.; Rotz, A.; Waghorn, G.; Adesogan, A.; Dijkstra, J.; Montes, F.; Oh, J.; Kebreab, E.; et al. Mitigation of methane and nitrous oxide emissions from animal operations: III. A review of animal management mitigation options. J. Anim. Sci. 2013, 91, 5095–5113. [Google Scholar] [CrossRef] [Green Version]
- Gebbels, J.N.; Kragt, M.E.; Thomas, D.T.; Vercoe, P.E. Improving productivity reduces methane intensity but increases the net emissions of sheepmeat and wool enterprises. Animal 2022, 16, 100490. [Google Scholar] [CrossRef]
- FAO. World Livestock 2011. Livestock in Food Security; Food and Agriculture Organization of the United Nations: Rome, Italy, 2011; 130p. [Google Scholar]
- FAO. The Future of Food and Agriculture—Alternative Pathways to 2050; Food and Agriculture Organization of the United Nations: Rome, Italy, 2018; 224p. [Google Scholar]
- IPCC. Summary for policymakers. In Global Warming of 1.5 °C; Masson-Delmotte, V., Zhai, P., Pörtner, H.-O., Roberts, D., Skea, J., Shukla, P.R., Pirani, A., Moufouma-Okia, W., Péan, C., Pidcock, R., et al., Eds.; An IPCC Special Report on the Impacts of Global Warming of 1.5 °C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2018; pp. 3–24. [Google Scholar] [CrossRef]
- Jackson, R.B.; Saunois, M.; Bousquet, P.; Canadell, J.G.; Poulter, B.; Stavert, A.R.; Bergamaschi, P.; Niwa, Y.; Segers, A.; Tsuruta, A. Increasing anthropogenic methane emissions arise equally from agricultural and fossil fuel sources. Environ. Res. Lett. 2020, 15, 071002. [Google Scholar] [CrossRef]
- Ungerfeld, E.M.; Beauchemin, K.A.; Muñoz, C. Current perspectives on achieving pronounced enteric methane mitigation from ruminant production. Front. Anim. Sci. 2022, 2, 795200. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Ungerfeld, E.M.; Eckard, R.J.; Wang, M. Review: Fifty years of research on rumen methanogenesis: Lessons learned and future challenges for mitigation. Animal 2020, 14, s2–s16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cottle, D.J.; Nolan, J.V.; Wiedemann, S.G. Ruminant enteric methane mitigation: A review. Anim. Prod. Sci. 2011, 51, 491–514. [Google Scholar] [CrossRef]
- Clark, H. Nutritional and host effects on methanogenesis in the grazing ruminant. Animal 2012, 7, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Clark, H.; Eckard, R.J. Mitigating methane in a systems context. In Proceedings of the 4th Australasian Dairy Science Symposium, Christchurch, New Zealand, 31 August–2 September 2010; pp. 78–85. [Google Scholar]
- Eckard, R.J.; Grainger, C.; De Klein, C.A.M. Options for the abatement of methane and nitrous oxide from ruminant production: A review. Livest. Sci. 2010, 130, 47–56. [Google Scholar] [CrossRef]
- Goopy, J. Creating a low enteric methane emission ruminant: What is the evidence of success to the present and prospects for developing economies? Anim. Prod. Sci. 2019, 59, 1759–1776. [Google Scholar] [CrossRef] [Green Version]
- Hristov, A.N.; Oh, J.; Firkins, J.L.; Dijkstra, J.; Kebreab, E.; Waghorn, G.; Makkar, H.P.S.; Adesogan, A.T.; Yang, W.; Lee, C.; et al. Mitigation of methane and nitrous oxide emissions from animal operations: I. A review of enteric methane mitigation options. J. Anim. Sci. 2013, 91, 5045–5069. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.; Morgavi, D.P.; Doreau, M. Methane mitigation in ruminants: From microbe to the farm scale. Animal 2010, 4, 351–365. [Google Scholar] [CrossRef]
- Almeida, A.K.; Hegarty, R.S.; Cowie, A. Meta-analysis quantifying the potential of dietary additives and rumen modifiers for methane mitigation in ruminant production systems. Anim. Nutr. 2021, 7, 1219–1230. [Google Scholar] [CrossRef] [PubMed]
- Arndt, C.; Hristov, A.N.; Price, W.J.; McClelland, S.C.; Pelaez, A.M.; Cueva, S.F.; Oh, J.; Dijkstra, J.; Bannink, A.; Bayat, A.R.; et al. Full adoption of the most effective strategies to mitigate methane emissions by ruminants can help meet the 1.5 °C target by 2030 but not 2050. Proc. Natl. Acad. Sci. USA 2022, 119, e2111294119. [Google Scholar] [CrossRef]
- Veneman, J.B.; Saetnan, E.R.; Clare, A.J.; Newbold, C.J. MitiGate; an online meta-analysis database for quantification of mitigation strategies for enteric methane emissions. Sci. Total Environ. 2016, 572, 1166–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dijkstra, J.; Bannink, A.; France, J.; Kebreab, E.; van Gastelen, S. Short communication: Antimethanogenic effects of 3-nitrooxypropanol depend on supplementation dose, dietary fiber content, and cattle type. J. Dairy Sci. 2018, 101, 9041–9047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayanegara, A.; Sarwono, K.A.; Kondo, M.; Matsui, H.; Ridla, M.; Laconi, E.B.; Nahrowi. Use of 3-nitrooxypropanol as feed additive for mitigating enteric methane emissions from ruminants: A meta-analysis. Ital. J. Anim. Sci. 2018, 17, 650–656. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.; Beauchemin, K.A. A review of feeding supplementary nitrate to ruminant animals: Nitrate toxicity, methane emissions, and production performance. Can. J. Anim. Sci. 2014, 94, 557–570. [Google Scholar] [CrossRef]
- Kim, H.; Lee, H.G.; Baek, Y.C.; Lee, S.; Seo, J. The effects of dietary supplementation with 3-nitrooxypropanol on enteric methane emissions, rumen fermentation, and production performance in ruminants: A meta-analysis. J. Anim. Sci. Technol. 2020, 62, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Lean, I.J.; Golder, H.M.; Grant, T.M.D.; Moate, P.J. A meta-analysis of effects of dietary seaweed on beef and dairy cattle performance and methane yield. PLoS ONE 2021, 16, e0249053. [Google Scholar] [CrossRef]
- Patra, A.K. Meta-analyses of effects of phytochemicals on digestibility and rumen fermentation characteristics associated with methanogenesis. J. Sci. Food Agric. 2010, 90, 2700–2708. [Google Scholar] [CrossRef]
- Legesse, G.; Beauchemin, K.A.; Ominski, K.H.; McGeough, E.J.; Kroebel, R.; MacDonald, D.; Little, S.M.; McAllister, T.A. Greenhouse gas emissions of Canadian beef production in 1981 as compared with 2011. Anim. Prod. Sci. 2016, 56, 153–168. [Google Scholar] [CrossRef]
- Global Livestock Environmental Assessment Model (GLEAM). Available online: www.fao.org/gleam/en/ (accessed on 22 July 2021).
- Ungerfeld, E.M.; Prediction of Enteric Methane Emissions and Mitigation under Different Scenarios of Intensification and Adoption and Effectiveness of Inhibitors of Rumen Methanogenesis. Released on: 30 August 2022. Available online: https://osf.io/drste/?view_only=2d0ee909617444a8b8568e50721d6e01 (accessed on 30 August 2022).
- Trei, J.E.; Parish, R.C.; Singh, Y.K. Effect of methane inhibitors on rumen metabolism and feedlot performance of sheep. J. Dairy Sci. 1971, 54, 536–540. [Google Scholar] [CrossRef]
- Johnson, E.D.; Wood, A.S.; Stone, J.B.; Moran, E.T., Jr. Some effects of methane inhibition in ruminants (steers). Can. J. Anim. Sci. 1972, 52, 703–712. [Google Scholar] [CrossRef]
- Davies, A.; Nwaonu, H.N.; Stanier, G.; Boyle, F.T. Properties of a novel series of inhibitors of rumen methanogenesis; in vitro and in vivo experiments including growth trials on 2,4-bis (trichloromethyl)-benzo(1,3)dioxin-6-carboxylic acid. Brit. J. Nutr. 1982, 47, 565–576. [Google Scholar] [CrossRef] [Green Version]
- Romero-Perez, A.; Okine, E.K.; McGinn, S.M.; Guan, L.L.; Oba, M.; Duval, S.M.; Kindermann, M.; Beauchemin, K.A. Sustained reduction in methane production from long-term addition of 3-nitrooxypropanol to a beef cattle diet. J. Anim. Sci. 2015, 93, 1780–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vyas, D.; McGinn, S.M.; Duval, S.M.; Kindermann, M.; Beauchemin, K.A. Effects of sustained reduction of enteric methane emissions with dietary supplementation of 3-nitrooxypropanol on growth performance of growing and finishing beef cattle. J. Anim. Sci. 2016, 94, 2024–2034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinley, R.D.; Martinez-Fernandez, G.; Matthews, M.K.; de Nys, R.; Magnusson, M.; Tomkins, N.W. Mitigating the carbon footprint and improving productivity of ruminant livestock agriculture using a red seaweed. J. Clean. Prod. 2020, 259, 120836. [Google Scholar] [CrossRef]
- Roque, B.M.; Venegas, M.; Kinley, R.D.; de Nys, R.; Duarte, T.L.; Yang, X.; Kebreab, E. Red seaweed (Asparagopsis taxiformis) supplementation reduces enteric methane by over 80 percent in beef steers. PLoS ONE 2021, 16, e0247820. [Google Scholar] [CrossRef] [PubMed]
- Alemu, A.W.; Shreck, A.L.; Booker, C.W.; McGinn, S.M.; Pekrul, L.K.D.; Kindermann, M.; Beauchemin, K.A. Use of 3-nitrooxypropanol in a commercial feedlot to decrease enteric methane emissions from cattle fed a corn-based finishing diet. J. Anim. Sci. 2021, 99, skaa394. [Google Scholar] [CrossRef]
- Cristobal-Carballo, O.; McCoard, S.A.; Cookson, A.L.; Ganesh, S.; Lowe, K.; Laven, R.A.; Muetzel, S. Effect of methane inhibitors on ruminal microbiota during early life and its relationship with ruminal metabolism and growth in calves. Front. Microbiol. 2021, 12, 710914. [Google Scholar] [CrossRef]
- Trei, J.E.; Scott, G.C.; Parish, R.C. Influence of methane inhibition on energetic efficiency of lambs. J. Anim. Sci. 1972, 34, 510–515. [Google Scholar] [CrossRef]
- Haisan, J.; Sun, Y.; Guan, L.L.; Beauchemin, K.A.; Iwaasa, A.; Duval, S.; Barreda, D.R.; Oba, M. The effects of feeding 3-nitrooxypropanol on methane emissions and productivity of Holstein cows in mid lactation. J. Dairy Sci. 2014, 97, 3110–3119. [Google Scholar] [CrossRef] [Green Version]
- Roque, B.M.; Salwen, J.K.; Kinley, R.; Kebreab, E. Inclusion of Asparagopsis armata in lactating dairy cows’ diet reduces enteric methane emission by over 50 percent. J. Clean. Prod. 2019, 234, 132–138. [Google Scholar] [CrossRef]
- Hristov, A.N.; Oh, J.; Lee, C.; Meinen, R.; Montes, F.; Ott, T.; Firkins, J.; Rotz, A.; Dell, C.; Adesogan, A.; et al. Mitigation of Greenhouse Gas Emissions in Livestock Production—A Review of Technical Options for Non-CO2 Emissions; FAO: Rome, Italy, 2013; Volume 177, 231p. [Google Scholar]
- Leahy, S.; Clark, H.; Reisinger, A. Challenges and prospects for agricultural greenhouse gas mitigation pathways consistent with the Paris agreement. Front. Sustain. Food Syst. 2020, 4, 69. [Google Scholar] [CrossRef]
- Feucht, Y.; Zander, K. Consumers’ Attitudes on Carbon Footprint Labelling: Results of the SUSDIET Project; Johann Heinrich von Thünen Institute, Federal Research Institute for Rural Areas, Forestry and Fisheries: Hamburg, Germany, 2017; 62p. [Google Scholar]
- FAO. Five Practical Actions towards Low-Carbon Livestock; FAO: Rome, Italy, 2019. [Google Scholar]
- Ritzman, E.G.; Benedict, F.G. Nutritional Physiology of the Adult Ruminant; Carnegie Institution of Washington: Washington, DC, USA, 1938; 200p. [Google Scholar]
- Blaxter, K.L.; Czerkawski, J. Modification of the methane production of the sheep by supplementation of its diet. J. Sci. Food Agric. 1966, 17, 417–421. [Google Scholar] [CrossRef]
- Hogan, K.; Hoffman, J.; Thompson, A. Methane on the greenhouse agenda. Nature 1991, 354, 181–182. [Google Scholar] [CrossRef]
- Crutzen, P.J.; Aselmann, I.; Seiler, W. Methane production by domestic animals, wild ruminants, other herbivorous fauna, and humans. Tellus B 1986, 38B, 271–284. [Google Scholar] [CrossRef]
- Johnson, K.A.; Johnson, D.E. Methane emissions from cattle. J. Anim. Sci. 1995, 73, 2483–2492. [Google Scholar] [CrossRef] [PubMed]
- McCrabb, G.J.; Berger, K.T.; Magner, T.; May, C.; Hunter, R.A. Inhibiting methane production in Brahman cattle by dietary supplementation with a novel compound and the effects on growth. Aust. J. Agr. Res. 1997, 48, 323–329. [Google Scholar] [CrossRef]
- Ungerfeld, E.M. Inhibition of rumen methanogenesis and ruminant productivity: A meta-analysis. Front. Vet. Sci. 2018, 5, 113. [Google Scholar] [CrossRef]
- Alemu, A.W.; Pekrul, L.K.D.; Shreck, A.L.; Booker, C.W.; McGinn, S.M.; Kindermann, M.; Beauchemin, K.A. 3-Nitrooxypropanol decreased enteric methane production from growing beef cattle in a commercial feedlot: Implications for sustainable beef cattle production. Front. Anim. Sci. 2021, 2, 641590. [Google Scholar] [CrossRef]
- Czerkawski, J.W. An Introduction to Rumen Studies, 1st ed.; Pergamon Press: Exeter, UK, 1986. [Google Scholar] [CrossRef]
- Hungate, R.E. Hydrogen as an intermediate in the rumen fermentation. Arch. Mikrobiol. 1967, 59, 158–164. [Google Scholar] [CrossRef]
- Ungerfeld, E.M. Metabolic hydrogen flows in rumen fermentation: Principles and possibilities of interventions. Front. Microbiol. 2020, 11, 589. [Google Scholar] [CrossRef] [Green Version]
- Ungerfeld, E.M. A theoretical comparison between two ruminal electron sinks. Front. Microbiol. 2013, 4, 319. [Google Scholar] [CrossRef] [Green Version]
- Hino, T.; Russell, J.B. Effect of reducing-equivalent disposal and NADH/NAD on deamination of amino acids by intact rumen microorganisms and their cell extracts. Appl. Environ. Microbiol. 1985, 50, 1368–1374. [Google Scholar] [CrossRef] [Green Version]
- Nagaraja, T.G.; Newbold, C.J.; van Nevel, C.J.; Demeyer, D.I. Manipulation of ruminal fermentation. In The Rumen Microbial Ecosystem, 2nd ed.; Hobson, P.N., Stewart, C.S., Eds.; Blackie Academic & Professional: London, UK, 1997; pp. 524–632. [Google Scholar] [CrossRef]
- Ungerfeld, E.M. Shifts in metabolic hydrogen sinks in the methanogenesis-inhibited ruminal fermentation: A meta-analysis. Front. Microbiol. 2015, 6, 37. [Google Scholar] [CrossRef] [PubMed]
- Ungerfeld, E.M.; Rust, S.R.; Burnett, R. Use of some novel alternative electron sinks to inhibit ruminal methanogenesis. Reprod. Nutr. Dev. 2003, 43, 189–202. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Fernandez, G.; Denman, S.E.; Yang, C.; Cheung, J.; Mitsumori, M.; McSweeney, C.S. Methane inhibition alters the microbial community, hydrogen flow, and fermentation response in the rumen of cattle. Front. Microbiol. 2016, 7, 1122. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Fernandez, G.; Denman, S.E.; Cheung, J.; McSweeney, C.S. Phloroglucinol degradation in the rumen promotes the capture of excess hydrogen generated from methanogenesis inhibition. Front. Microbiol. 2017, 8, 1871. [Google Scholar] [CrossRef]
- Melgar, A.; Harper, M.T.; Oh, J.; Giallongo, F.; Young, M.E.; Ott, T.L.; Duval, S.; Hristov, A.N. Effects of 3-nitrooxypropanol on rumen fermentation, lactational performance, and resumption of ovarian cyclicity in dairy cows. J. Dairy Sci. 2020, 103, 410–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, P.; Edwards, R.A.; Greenhalgh, J.F.D.; Morgan, C.A.; Sinclair, L.A.; Wilkinson, R.G. Animal Nutrition, 7th ed.; Prentice Hall: Harlow, UK, 2010; 714p. [Google Scholar]
- Qumar, M.; Khiaosa-Ard, R.; Pourazad, P.; Wetzels, S.U.; Klevenhusen, F.; Kandler, W.; Aschenbach, J.R.; Zebeli, Q. Evidence of in vivo absorption of lactate and modulation of short chain fatty acid absorption from the reticulorumen of non-lactating cattle fed high concentrate diets. PLoS ONE 2016, 11, e0164192. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Fernandez, G.; Duval, S.; Kindermann, M.; Schirra, H.J.; Denman, S.E.; McSweeney, C.S. 3-NOP vs. halogenated compound: Methane production, ruminal fermentation and microbial community response in forage fed cattle. Front. Microbiol. 2018, 9, 1582. [Google Scholar] [CrossRef]
- Janssen, P.H. Influence of hydrogen on rumen methane formation and fermentation balances through microbial growth kinetics and fermentation thermodynamics. Anim. Feed Sci. Technol. 2010, 160, 1–22. [Google Scholar] [CrossRef]
- Larsen, M.; Kristensen, N.B. Precursors for liver gluconeogenesis in periparturient dairy cows. Animal 2013, 7, 1640–1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loncke, C.; Nozière, P.; Vernet, J.; Lapierre, H.; Bahloul, L.; Al-Jammas, M.; Sauvant, D.; Ortigues-Marty, I. Net hepatic release of glucose from precursor supply in ruminants: A meta-analysis. Animal 2020, 14, 1422–1437. [Google Scholar] [CrossRef] [Green Version]
- Allen, M.S.; Bradford, B.J.; Oba, M. Board-invited review: The hepatic oxidation theory of the control of feed intake and its application to ruminants. J. Anim. Sci. 2009, 87, 3317–3334. [Google Scholar] [CrossRef] [Green Version]
- Nollet, L.; Demeyer, D.I.; Verstraete, W. Effect of 2-bromoethanesulfonic acid and Peptostreptococcus productus ATCC 35244 addition on stimulation of reductive acetogenesis in the ruminal ecosystem by selective inhibition of methanogenesis. Appl. Environ. Microbiol. 1997, 63, 194–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Van, T.D.; Robinson, J.A.; Ralph, J.; Greening, R.C.; Smolenski, W.J.; Leedle, J.A.Z.; Schaefer, D.M. Assessment of reductive acetogenesis with indigenous ruminal bacterium populations and Acetitomaculum ruminis. Appl. Environ. Microbiol. 1998, 64, 3429–3436. [Google Scholar] [CrossRef] [Green Version]
- Lopez, S.; McIntosh, F.M.; Wallace, R.J.; Newbold, C.J. Effect of adding acetogenic bacteria on methane production by mixed rumen microorganisms. Anim. Feed Sci. Technol. 1999, 78, 1–9. [Google Scholar] [CrossRef]
- Raju, P. Homoacetogenesis as an Alternative Hydrogen Sink in the Rumen. Ph.D. Thesis, University of Massey, Palmerston North, New Zealand, 2016. [Google Scholar]
- Ungerfeld, E.M.; Aedo, M.F.; Muñoz, C.; Urrutia, N.L.; Martínez, E.D.; Saldivia, M. Inhibiting methanogenesis stimulated de novo synthesis of microbial amino acids in mixed rumen batch cultures growing on starch but not on cellulose. Microorganisms 2020, 8, 799. [Google Scholar] [CrossRef]
- Abecia, L.; Toral, P.G.; Martín-García, A.I.; Martínez, G.; Tomkins, N.W.; Molina-Alcaide, E.; Newbold, C.J.; Yañez-Ruiz, D.R. Effect of bromochloromethane on methane emission, rumen fermentation pattern, milk yield, and fatty acid profile in lactating dairy goats. J. Dairy Sci. 2012, 95, 2027–2036. [Google Scholar] [CrossRef] [Green Version]
- Hristov, A.N.; Oh, J.; Giallongo, F.; Frederick, T.W.; Harper, M.T.; Weeks, H.L.; Branco, A.F.; Moate, P.J.; Deighton, M.H.; Williams, S.R.; et al. An inhibitor persistently decreased enteric methane emission from dairy cows with no negative effect on milk production. Proc. Natl. Acad. Sci. USA 2015, 112, 10663–10668. [Google Scholar] [CrossRef] [Green Version]
- Melgar, A.; Lage, C.F.A.; Nedelkov, K.; Räisänen, S.E.; Stefenoni, H.; Fetter, M.E.; Chen, X.; Oh, J.; Duval, S.; Kindermann, M.; et al. Enteric methane emission, milk production, and composition of dairy cows fed 3-nitrooxypropanol. J. Dairy Sci. 2021, 104, 357–366. [Google Scholar] [CrossRef]
- Melgar, A.; Welter, K.C.; Nedelkov, K.; Martins, C.M.M.R.; Harper, M.T.; Oh, J.; Räisänen, S.E.; Chen, X.; Cueva, S.F.; Duval, S.; et al. Dose-response effect of 3-nitrooxypropanol on enteric methane emissions in dairy cows. J. Dairy Sci. 2020, 103, 6145–6156. [Google Scholar] [CrossRef]
- Stefenoni, H.A.; Räisänen, S.E.; Cueva, S.F.; Wasson, D.E.; Lage, C.F.A.; Melgar, A.; Fetter, M.E.; Smith, P.; Hennessy, M.; Vecchiarelli, B.; et al. Effects of the macroalga Asparagopsis taxiformis and oregano leaves on methane emission, rumen fermentation, and lactational performance of dairy cows. J. Dairy Sci. 2021, 104, 4157–4173. [Google Scholar] [CrossRef]
- Vargas, J.; Ungerfeld, E.; Muñoz, C.; DiLorenzo, N. Feeding strategies to mitigate enteric methane emission from ruminants in grassland systems. Animals 2022, 12, 1132. [Google Scholar] [CrossRef] [PubMed]
- Hegarty, R.S.; Passetti, R.A.C.; Dittmer, K.M.; Wang, Y.; Shelton, S.; Emmet-Booth, J.; Wollenberg, E.; McAllister, T.A.; Leahy, S.; Beauchemin, K.; et al. An Evaluation of Emerging Feed Additives to Reduce Methane Emissions from Livestock; Climate Change, Agriculture and Food Security (CCAFS) and the New Zealand Agricultural Greenhouse Gas Research Centre (NZAGRC) Initiative of the Global Research Alliance (GRA): 2021; 104p. Available online: https://cgspace.cgiar.org/handle/10568/116489 (accessed on 30 August 2022).
- Callaghan, M.J.; Tomkins, N.W.; Hepworth, G.; Parker, A.J. The effect of molasses nitrate lick blocks on supplement intake, bodyweight, condition score, blood methaemoglobin concentration and herd scale methane emissions in Bos indicus cows grazing poor quality forage. Anim. Prod. Sci. 2021, 61, 445–458. [Google Scholar] [CrossRef]
- Rowe, S.; Hickey, S.; Jonker, A.; Hess, M.; Janssen, P.; Johnson, T.; Bryson, B.; Knowler, K.; Pinares-Patino, C.; Bain, W.; et al. Selection for divergent methane yield in New Zealand sheep—A ten year perspective. In Proceedings of the Association for the Advancement of Animal Breeding and Genetics Conference, Armindale, NSW, Australia, 27 October–1 November 2019; Association for the Advancement of Animal Breeding and Genetics: Armindale, NSW, Australia, 2019; pp. 306–309. [Google Scholar]
- Meale, S.J.; Popova, M.; Saro, C.; Martin, C.; Bernard, A.; Lagree, M.; Yáñez-Ruiz, D.R.; Boudra, H.; Duval, S.; Morgavi, D.P. Early life dietary intervention in dairy calves results in a long-term reduction in methane emissions. Sci. Rep. 2021, 11, 3003. [Google Scholar] [CrossRef] [PubMed]
- Baca-González, V.; Asensio-Calavia, P.; González-Acosta, S.; Pérez de la Lastra, J.M.; Morales de la Nuez, A. Are vaccines the solution for methane emissions from ruminants? A systematic review. Vaccines 2020, 8, 460. [Google Scholar] [CrossRef] [PubMed]
- Thiel, A.; Rumbeli, R.; Mair, P.; Yeman, H.; Beilstein, P. 3-NOP: ADME studies in rats and ruminating animals. Food Chem. Toxicol. 2019, 125, 528–539. [Google Scholar] [CrossRef]
- Thiel, A.; Schoenmakers, A.C.M.; Verbaan, I.A.J.; Chenal, E.; Etheve, S.; Beilstein, P. 3-NOP: Mutagenicity and genotoxicity assessment. Food Chem. Toxicol. 2019, 123, 566–573. [Google Scholar] [CrossRef]
- Li, X.; Norman, H.C.; Kinley, R.D.; Laurence, M.; Wilmot, M.; Bender, H.; de Nys, R.; Tomkins, N. Asparagopsis taxiformis decreases enteric methane production from sheep. Anim. Prod. Sci. 2016, 58, 681–688. [Google Scholar] [CrossRef]
- Muizelaar, W.; Groot, M.; van Duinkerken, G.; Peters, R.; Dijkstra, J. Safety and transfer study: Transfer of bromoform present in Asparagopsis taxiformis to milk and urine of lactating dairy cows. Foods 2021, 10, 584. [Google Scholar] [CrossRef]
- Glasson, C.R.K.; Kinley, R.D.; de Nys, R.; King, N.; Adams, S.L.; Packer, M.A.; Svenson, J.; Eason, C.T.; Magnusson, M. Benefits and risks of including the bromoform containing seaweed Asparagopsis in feed for the reduction of methane production from ruminants. Algal Res. 2022, 64, 102673. [Google Scholar] [CrossRef]
- Jia, Y.; Quack, B.; Kinley, R.D.; Pisso, I.; Tegtmeier, S. Potential environmental impact of bromoform from Asparagopsis farming in Australia. Atmos. Chem. Phys. 2022, 22, 7631–7646. [Google Scholar] [CrossRef]
- Romero, P.; Belanche, A.; Hueso, R.; Ramos-Morales, E.; Salwen, J.K.; Kebreab, E.; Yañez-Ruiz, D.R. In In vitro rumen microbial degradation of bromoform and the impact on rumen fermentation. In Proceedings of the 8th International Greenhouse Gas & Animal Agriculture Conference, Orlando, FL, USA, 5–10 June 2022; University of Florida: Orlando, FL, USA, 2022; p. 253. [Google Scholar]
- PubChem Annotation Record for Dibromomethane. Available online: https://pubchem.ncbi.nlm.nih.gov/source/hsdb/1334 (accessed on 8 July 2022).
- Haisan, J.; Sun, Y.; Guan, L.L.; Beauchemin, K.A.; Iwaasa, A.; Duval, S.; Kindermann, M.; Barreda, D.R.; Oba, M. The effects of feeding 3-nitrooxypropanol at two doses on milk production, rumen fermentation, plasma metabolites, nutrient digestibility, and methane emissions in lactating Holstein cows. Anim. Prod. Sci. 2017, 57, 282–289. [Google Scholar] [CrossRef]
- Schilde, M.; von Soosten, D.; Hüther, L.; Kersten, S.; Meyer, U.; Zeyner, A.; Dänicke, S. Dose–response effects of 3-nitrooxypropanol combined with low- and high-concentrate feed proportions in the dairy cow ration on fermentation parameters in a rumen simulation technique. Animals 2021, 11, 1784. [Google Scholar] [CrossRef]
- Pitta, D.; Indugu, N.; Melgar, A.; Hristov, A.; Challa, K.; Vecchiarelli, B.; Hennessy, M.; Narayan, K.; Duval, S.; Kindermann, M.; et al. The Effect of 3-nitrooxypropanol, a Potent Methane Inhibitor, on Ruminal Microbial Gene Expression Profiles in Dairy Cows. Microbiome 2022, 10, 146. [Google Scholar] [CrossRef] [PubMed]
- Pitta, D.W.; Melgar, A.; Hristov, A.N.; Indugu, N.; Narayan, K.S.; Pappalardo, C.; Hennessy, M.L.; Vecchiarelli, B.; Kaplan-Shabtai, V.; Kindermann, M.; et al. Temporal changes in total and metabolically active ruminal methanogens in dairy cows supplemented with 3-nitrooxypropanol. J. Dairy Sci. 2021, 104, 8721–8735. [Google Scholar] [CrossRef] [PubMed]
- Long, F.; Cheung, C.Y.; Whitman, W.B.; Cook, G.M.; Ronimus, R.S. Using genome comparisons of wild-type and resistant mutants of Methanococcus maripaludis to help understand mechanisms of resistance to methane inhibitors. Access Microbiol. 2021, 3, 000244. [Google Scholar] [CrossRef]
- Duin, E.C.; Wagner, T.; Shima, S.; Prakash, D.; Cronin, B.; Yanez-Ruiz, D.R.; Duval, S.; Rumbeli, R.; Stemmler, R.T.; Thauer, R.K.; et al. Mode of action uncovered for the specific reduction of methane emissions from ruminants by the small molecule 3-nitrooxypropanol. Proc. Natl. Acad. Sci. USA 2016, 113, 6172–6177. [Google Scholar] [CrossRef] [Green Version]
- Ungerfeld, E.M.; Rust, S.R.; Boone, D.R.; Liu, Y. Effects of several inhibitors on pure cultures of ruminal methanogens. J. Appl. Microbiol. 2004, 97, 520–526. [Google Scholar] [CrossRef]
- Gunsalus, R.P.; Romesser, J.A.; Wolfe, R.S. Preparation of coenzyme M analogs and their activity in the methyl coenzyme M reductase system of Methanobacterium thermoautotrophicum. Biochemistry 1978, 17, 2374–2377. [Google Scholar] [CrossRef] [PubMed]
- Van Nevel, C.J.; Demeyer, D.I. Control of rumen methanogenesis. Environ. Monit. Assess. 1996, 42, 73–97. [Google Scholar] [CrossRef]
- Leahy, S.C.; Kelly, W.J.; Altermann, E.; Ronimus, R.S.; Yeoman, C.J.; Pacheco, D.M.; Li, D.; Kong, Z.; McTavish, S.; Sang, C.; et al. The genome sequence of the rumen methanogen Methanobrevibacter ruminantium reveals new possibilities for controlling ruminant methane emissions. PLoS ONE 2010, 5, e8926. [Google Scholar] [CrossRef] [Green Version]
- Taylor, C.D.; McBride, B.C.; Wolfe, R.S.; Bryant, M.P. Coenzyme M, essential for growth of a rumen strain of Methanobacterium ruminantium. J. Bacteriol. 1974, 120, 974–975. [Google Scholar] [CrossRef] [Green Version]
- Balch, W.E.; Wolfe, R.S. Transport of coenzyme M (2-mercaptoethanesulfonic acid) in Methanobacterium ruminantium. J. Bacteriol. 1979, 137, 264–273. [Google Scholar] [CrossRef] [Green Version]
- Santoro, N.; Konisky, J. Characterization of bromoethanesulfonate-resistant mutants of Methanococcus voltae: Evidence of a coenzyme M transport system. J. Bacteriol. 1987, 169, 660–665. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.R. Reversal of 2-bromoethanesulfonate inhibition of methanogenesis in Methanosarcina sp. J. Bacteriol. 1983, 156, 516–523. [Google Scholar] [CrossRef]
- Gruninger, R.J.; Zhang, X.M.; Smith, M.L.; Kung, L.; Vyas, D.; McGinn, S.M.; Kindermann, M.; Wang, M.; Tan, Z.L.; Beauchemin, K.A. Application of 3-nitrooxypropanol and canola oil to mitigate enteric methane emissions of beef cattle results in distinctly different effects on the rumen microbial community. Anim. Microbiome 2022, 4, 35. [Google Scholar] [CrossRef] [PubMed]
- Feldewert, C.; Lang, K.; Brune, A. The hydrogen threshold of obligately methyl-reducing methanogens. FEMS Microbiol. Lett. 2020, 367, fnaa137. [Google Scholar] [CrossRef]
- Wood, J.M.; Kennedy, F.S.; Wolfe, R.S. The reaction of multihalogenated hydrocarbons with free and bound reduced vitamin B12. Biochemistry 1968, 7, 1707–1713. [Google Scholar] [CrossRef] [PubMed]
- Dumitru, R.; Palencia, H.; Schroeder, S.D.; DeMontigny, B.A.; Takacs, J.M.; Rasche, M.E.; Miner, J.L.; Ragsdale, S.W. Targeting methanopterin biosynthesis to inhibit methanogenesis. Appl. Environ. Microbiol. 2003, 69, 7236–7241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, T.L.; Wolin, M.J. Inhibition of growth of methane-producing bacteria of the ruminant forestomach by hydroxymethylglutaryl∼scoa reductase inhibitors. J. Dairy Sci. 2001, 84, 1445–1448. [Google Scholar] [CrossRef]
- Van Lingen, H.J.; Fadel, J.G.; Yáñez-Ruiz, D.R.; Kindermann, M.; Kebreab, E. Inhibited methanogenesis in the rumen of cattle: Microbial metabolism in response to supplemental 3-nitrooxypropanol and nitrate. Front. Microbiol. 2021, 12, 705613. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.K.; Humphries, D.J.; Kirton, P.; Kindermann, M.; Duval, S.; Steinberg, W. Effects of 3-nitrooxypropanol on methane emission, digestion, and energy and nitrogen balance of lactating dairy cows. J. Dairy Sci. 2014, 97, 3777–3789. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal Product | Metric | Period Used for Estimation | Total Change (%) | Rate (%/yr) | Source Used for Estimation |
|---|---|---|---|---|---|
| Beef | Production | 2010–2050 | 58 | 1.15 | FAO [13] |
| Lamb | Production | 2010–2050 | 78 | 1.45 | FAO [13] |
| Milk | Production | 2010–2050 | 58 | 1.15 | FAO [13] |
| Beef | CH4 intensity | 2002–2016 | −0.52 | −0.037 | Chang et al. [7] 1 |
| Lamb | CH4 intensity | 2002–2016 | −7.27 | −0.54 | Chang et al. [7] 2 |
| Milk | CH4 intensity | 2002–2016 | −9.55 | −0.71 | Chang et al. [7] 2 |
| Reference | Animal, Diet | Inhibitor/Algae (g/kg Diet DM 2) | Experimental Period (d) | Inhibition Relative to Control Treatment (% Decrease in CH4 Animal−1 d−1) | Inhibition Relative to Control Treatment (% Decrease in CH4 kg ADG−1) | Performance | ||
|---|---|---|---|---|---|---|---|---|
| DMI | ADG | G:F | ||||||
| Trei et al. [38] | Lambs, mixed | 2, 2, 2-trichloroacetamide (0.080) | 90 | 67 3 | 67 4 | NS 5 | NS | ↑ |
| Johnson et al. [39] | Steers, mixed | BCM (0.50) | 28 | ~65 6 | ~68 4 | NS | NS | - |
| Davies et al. [40] | Calves, mixed | ICI 13409 (0.20) | 196 | 63 3 | 66 4 | ↓ | ↑ | ↑ |
| Romero-Perez et al. [41] | Heifers, mixed | 3-NOP (0.28) | 112 | 59 | 60 | R | NS | NS |
| Vyas et al. [42], finishing diet | Steers, high concentrate | 3-NOP (0.2) | 105 | 84 | 83 | ↓ | ↓ | NS |
| Kinley et al. [43] | Steers, high concentrate | Asparagopsis taxiformis (1.8) | 90 | 98 | 98 | NS | ↑ | NS |
| Roque et al. [44] | Steers, high concentrate | Asparagopsis taxiformis (4.7) | 63 | 82 | 83 | ↓ | NS | NS |
| Alemu et al. [45] | Steers, high concentrate | 3-NOP (0.108) | 112 | 77 | 76 4 | ↓ 7 | ↓ 7 | ↑ 7 |
| Cristobal-Carballo et al. [46] | Calves, milk replacer, concentrate, partial mixed ration, pasture | Chloroform (0.050) plus 9, 10-anthraquinone (0.50) | 84 | 90 3 | 90 | NS | NS | - |
| Reference | Animal, Diet | Inhibitor/Algae (g/kg Diet DM 2) | Experimental Period (d) | Inhibition Relative to Control Treatment (% Decrease in CH4 Animal−1 d−1) | Inhibition Relative to Control Treatment (% Decrease in CH4 kg FPCM−1) | Performance | ||
|---|---|---|---|---|---|---|---|---|
| DMI | MY | MY:F | ||||||
| Haisan et al. [48] | Cows, mixed | 3-NOP (0.13) | 28 | 60 | 61 3 | NS 4 | NS | NS |
| Roque et al. [49] | Cows, mixed | Asparagopsis armata (10) | 21 | 67 | 61 3 | ↓ | ↓ | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ungerfeld, E.M. Opportunities and Hurdles to the Adoption and Enhanced Efficacy of Feed Additives towards Pronounced Mitigation of Enteric Methane Emissions from Ruminant Livestock. Methane 2022, 1, 262-285. https://doi.org/10.3390/methane1040021
Ungerfeld EM. Opportunities and Hurdles to the Adoption and Enhanced Efficacy of Feed Additives towards Pronounced Mitigation of Enteric Methane Emissions from Ruminant Livestock. Methane. 2022; 1(4):262-285. https://doi.org/10.3390/methane1040021
Chicago/Turabian StyleUngerfeld, Emilio M. 2022. "Opportunities and Hurdles to the Adoption and Enhanced Efficacy of Feed Additives towards Pronounced Mitigation of Enteric Methane Emissions from Ruminant Livestock" Methane 1, no. 4: 262-285. https://doi.org/10.3390/methane1040021