
DNA Barcoding and Phylogenetics Relationship of Pangasiid Catfishes in Peninsular Malaysia Revealed the Impacts of Aquaculture on the Native Species Conservation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Samples and DNA Extraction
2.2. PCR Amplification and Sequencing
2.3. Phylogenetic Analysis
2.4. Automatic Barcoding Gap Discovery (ABGD)
3. Results
3.1. Genetic Variation
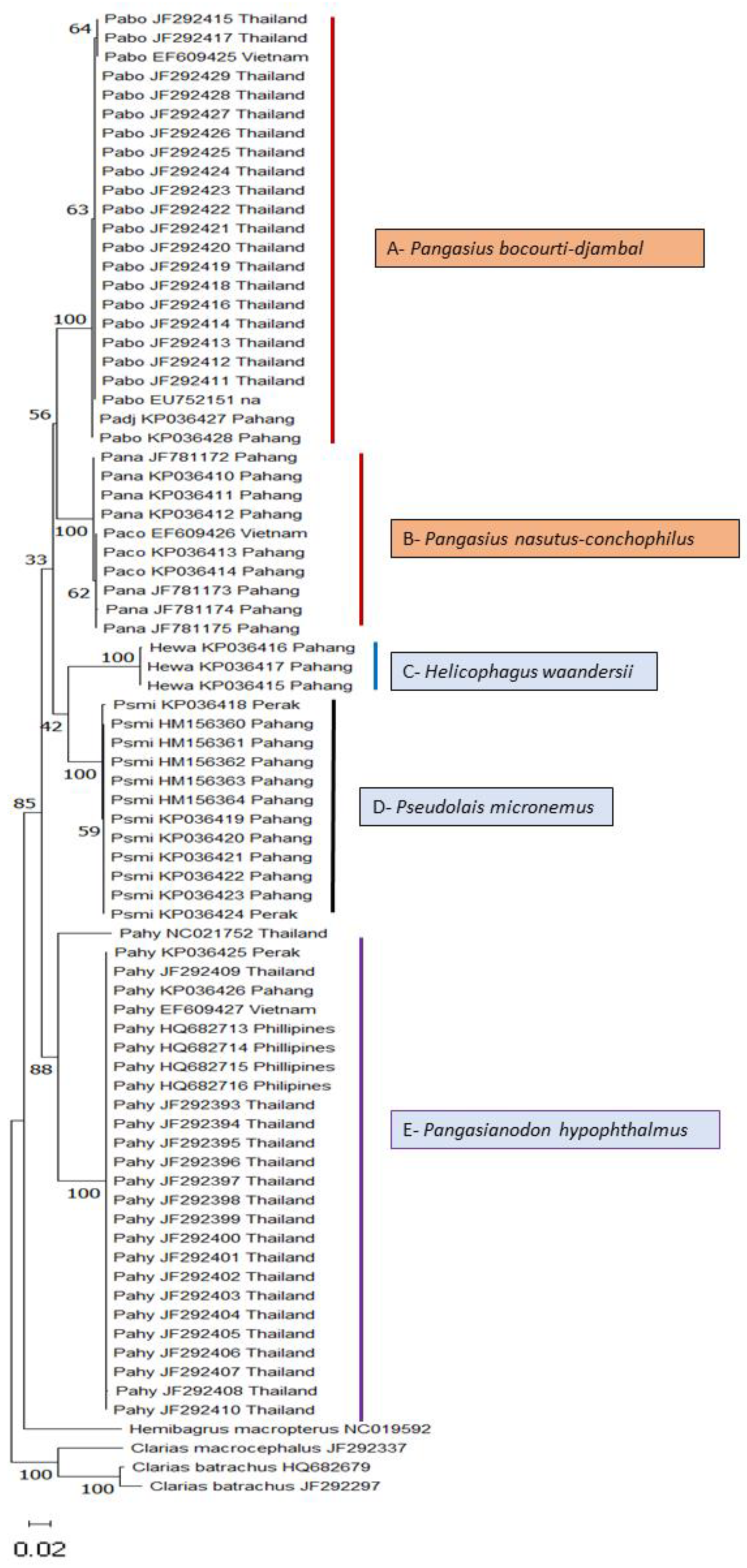
3.2. Phylogenetic Analysis—Neighbour-Joining (NJ)
3.3. Automatic Barcoding Gap Discovery (ABGD)
4. Discussion
4.1. Species Relationship and Taxonomic Accounts
4.2. Pangasiid Species Diversity and Impacts on the Current Aquaculture Practices
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gustiano, R.; Prakoso, V.A.; Ath-thar, M.H.F.; Kusmini, I.I.; Radona, D. Similarity and genetic relationship analysis of 28 species of Pangasiidae (Siluriformes, Ostariophysi). IOP Conf. Ser. Earth Environ. Sci. 2021, 674, 012014. [Google Scholar] [CrossRef]
- Ha, T.T.T.; Nga, T.T.; Hang, T.N.A.; Alam, M.S. Genetic diversity in Pangasius spp. collected in Bangladesh based on mitochondrial cytochrome b gene sequence analysis. Aquac. Rep. 2020, 17, 100351. [Google Scholar] [CrossRef]
- Roberts, T.R.; Vidthayanon, C. Systematic Revision of the Asian Catfish Family Pangasiidae, with Biological Observations and Description of Three New Species. PNAS 1991, 143, 97–144. [Google Scholar]
- Yang, J.X.; Chen, X.Y.; Chen, Y.R. On the population status and migration of Pangasiid catfishes in Lancangjiang River Basin, China. Zool. Res. 2007, 28, 63–67. [Google Scholar]
- Kottelat, M. The fishes of the inland waters of Southeast Asia: A catalogue and core bibliography of the fishes known to occur in freshwaters, mangroves and estuaries. Raffles Bull. Zool. 2013, 27, 1–663. [Google Scholar]
- Gustiano, R. Taxonomy and Phylogeny of Pangasiidae Catfishes from Asia Ostariophysis, Siluriformes). Ph.D. Thesis, Katholieke Universiteit Leuven, Leuven, Belgium, 2003; 296p. [Google Scholar]
- Ayyathurai, K.P.V.; Kodeeswaran, P.; Mohindra, V.; Singh, R.K.; Ravi, C.; Kumar, R.; Valaparambil, B.S.; Thipramalai Thangappan, A.K.; Jena, J.; Lal, K.K. Description of a new Pangasius (Valenciennes, 1840) species, from the Cauvery River extends distribution range of the genus up to South Western Ghats in peninsular India. PeerJ 2022, 10, e14258. [Google Scholar] [CrossRef]
- Gustiano, R.; Teugels, G.G.; Pouyaud, L. Pangasius bedado Roberts, 1999: A junior synonym of Pangasius djambal Bleeker, 1846 (Siluriformes, Pangasiidae). Cybium 2004, 28, 13–18. [Google Scholar]
- Pouyaud, L.; Gustiano, R.; Teugels, G.G. Systematic revision of Pangasius polyuranodon (Siluriformes, Pangasiidae) with description of two new species. Cybium 2002, 26, 243–252. [Google Scholar]
- Haslawati, B.; Rizman-Idid, M.; Sekaran, M.; Zakaria-Ismail, M. The occurrence of Pangasius polyuranodon Bleeker 1852 (Teleostei: Pangasiidae) in Peninsular Malaysia with remarks on the comparative morphology with Pseudolais micronemus Bleeker (1847). Sains Malays. 2014, 43, 1707–1714. [Google Scholar]
- FAO. Food and Agriculture Organisation of the United Nations. Fisheries and Aquaculture Department. 2008. Available online: http://www.fao.org/fishery/culturedspecies/Pangasius_hypophthalmus/en (accessed on 15 January 2023).
- Globefish. Market Report Pangasius. June 2013. Available online: http://www.globefish.org/pangasius-june-2013.html (accessed on 16 September 2013).
- Sittichai Hatachote, S.; Sriphairoj, K.; Na-Nakorn, U. Growth and Flesh Quality of the Hybrid between Pangasianodon hypophthalmus and Pangasius bocourti. Kasetsart J. (Nat. Sci.) 2015, 49, 717–725. [Google Scholar]
- Phan, L.T.; Bui, T.M.; Nguyen, T.T.T.; Gooley, G.J.; Ingram, B.A. Current status of farming practices of striped catfish, Pangasianodon hypophthalmus in the Mekong Delta, Vietnam. Aquaculture 2009, 296, 227–236. [Google Scholar] [CrossRef]
- Singh, A.K.; Lakra, W.S. Culture of Pangasianodon hypophthalmus into India: Impacts and Present Scenario. PJBS 2012, 15, 19–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Więcaszek, B.; Keszka, S.; Sobecka, E.; Boeger, W.A. Asian pangasiids—An emerging problem for European inland waters? Systematic and parasitological aspects. Acta Ichthyol Piscat. 2009, 39, 131–138. [Google Scholar] [CrossRef] [Green Version]
- Osathanunkula, M. An eDNA detection of captive-bred Mekong Giant Catfish in the Chao Phraya River basin for further environmental impacts assessment. Aquaculture 2022, 546, 737328. [Google Scholar] [CrossRef]
- Lim, K.K.P.; Zakaria-Ismail, M. The occurrence of the catfish Helicophagus waandersii (Pisces: Pangasiidae) in Peninsular Malaysia. Malay. Nat. J. 1995, 49, 37–40. [Google Scholar]
- Vidthayanon, C. Pangasius nasutus. The IUCN Red List of Threatened Species. Version 2014.3. 2013. Available online: www.iucnredlist.org (accessed on 8 March 2015).
- Vidthayanon, C. Pangasius polyuranodon. The IUCN Red List of Threatened Species. Version 2014.3. 2013. Available online: www.iucnredlist.org (accessed on 8 March 2015).
- Chong, V.C.; Lee, P.K.Y.; Lau, C.M. Diversity, extinction risk and conservation of Malaysian fishes. J. Fish Biol. 2010, 76, 2009–2066. [Google Scholar] [CrossRef]
- Bellemain, E.; Patricio, H.; Gray, T.; Guegan, F.; Valentini, A.; Miaud, C.; Dejean, T. Trails of river monsters: Detecting critically endangered Mekong giant catfish Pangasianodon gigas using environmental DNA. Glob. Ecol. Conserv. 2016, 7, 148–156. [Google Scholar]
- Anuar, H.; Ambak, M.A.; Abdul-Samad, A.P. Crossbreeding of Pangasianodon hypophthalmus (Sauvage, 1878) and Pangasius nasutus (Bleeker, 1863) and their larval development. J Sustain Sci Manag. 2011, 6, 28–35. [Google Scholar]
- Na-Nakorn, U.; Kamonrat, W.; Ngamsiri, T. Genetic diversity of walking catfish, Clarias microcephalus, in Thailand and evidence of genetic interogression from introduced farmed C. gariepinus. Aquaculture 2004, 240, 145–163. [Google Scholar] [CrossRef]
- Nazir, A.; Khan, M.A. Stock discrimination of Sperata aor from river Ganga using microsatellite markers: Implications for conservation and management. Aquat. Living Resour. 2017, 30, 33. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.A.; Bhadra, A.; Begum, N.; Islam, M.S.; Hussain, M.F. Production of hybrid vigor cross-breeding between Clarias batrachus Lin. & Clarias gariepinus Bur. Aquaculture 1995, 138, 125–130. [Google Scholar]
- Azmi, N.A.R.; Ahmad, L.W.Z.; Kambol, R.; Mohamad, S.A.S.; Aris, F.; Zakaria, N.A.; Yurus, N. Genetic variation analysis between wild and cultured Pangasianodon hypopthalmus using COI and cytochrome b among Asian countries. Sci. Lett. 2022, 16, 84–101. [Google Scholar] [CrossRef]
- Serrao, N.R.; Steinke, D.; Hanner, R.H. Calibrating snakehead diversity with DNA barcodes: Expanding taxonomic coverage to enable identification of potential and established invasive species. PLoS ONE 2014, 9, e99546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D.N. DNA barcoding Australiaʼs fish species. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 1847–1857. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Buhay, J.E.; Whiting, M.F.; Crandall, K.A. Many species in one: DNA barcoding overestimates the number of species when nuclear mitochondrial pseudogenes are coamplified. Proc. Natl. Acad. Sci. USA 2008, 105, 13486–13491. [Google Scholar] [CrossRef] [Green Version]
- MacCarthy, C. CHROMAS 1.45 Program; Griffith University: Brisbane, Australia, 1996. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The ClustalX windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 24, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarty, P.; Warren, M.; Page, L.M.; Baldwin, C.C. GenSeq: An updated nomenclature and ranking for genetic sequences from type and non-type sources. ZooKeys 2013, 346, 29–41. [Google Scholar] [CrossRef]
- Song, L.M.; Munian, K.; Zulkafli, A.R.; Bhassu, S. Characterisation of Asian Snakehead Murrel Channa striata (Channidae) in Malaysia: An insight into molecular data and morphological approach. Sci. World J. 2013, 16, 917506. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Aquino, L.M.G.; Tango, J.M.; Canoy, R.J.C.; Fontanilla, I.K.C.; Basiao, Z.U.; Ong, P.S.; Quilang, J.P. DNA barcoding of fishes of Laguna de Bay, Philippines. Mitochondrial DNA 2011, 22, 143–153. [Google Scholar] [CrossRef]
- Zhao, H.; Kong, X.; Zhou, C. The mitogenome of Pangasius sutchi (Teleotei, Siluriformes: Pangasiidae). Mitochondrial DNA 2014, 25, 342–344. [Google Scholar] [CrossRef] [PubMed]
- Wong, L.L.; Peatman, E.; Lu, J.; Kucuktas, H.; He, S.; Zhou, C.; Na-nakorn, U.; Liu, Z. DNA Barcoding of Catfish: Species Authentication and Phylogenetic Assessment. PLoS ONE 2011, 6, e17812. [Google Scholar] [CrossRef] [PubMed]
- Yancy, H.F.; Zemlak, T.S.; Mason, J.A.; Washington, J.D.; Tenge, B.J.; Nguyen, N.-L.; Barnett, J.D.; Savary, W.E.; Hill, W.E.; Moore, M.M.; et al. The potential use of DNA barcodes in regulatory science: Applications of the Regulatory Fish Encyclopedia. J. Food Prot. 2008, 71, 210–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, R.D.; Holmes, B.H. An analysis of nucleotide and amino acid variability in the barcode region of cytochrome c oxidase I (cox1) in fishes. Mol. Ecol. Notes 2007, 6, 899–907. [Google Scholar] [CrossRef]
- Zeng, Q.; Ye, H.; Peng, Z.; Wang, Z. Mitochondrial genome of Hemibagrus macropterus (Teleostei, Siluriformes). Mitochondrial DNA 2012, 23, 355–357. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef]
- Khare, P.; Mohindra, V.; Barman, A.S.; Singh, R.K.; Lal, K.K. Molecular evidence to reconcile taxonomic instability in mahseer species (Pisces: Cyprinidae) of India. Org. Divers. Evol. 2014, 14, 307–326. [Google Scholar] [CrossRef]
- Khedkar, G.D.; Jamdade, R.; Naik, S.; David, L.; Haymer, D. DNA barcodes for the Fishes of the Narmada, one of Indiaʼs longest rivers. PLoS ONE 2014, 9, e101460. [Google Scholar] [CrossRef] [Green Version]
- Karinthanyakit, W.; Jondeung, A. Molecular phylogenetic relationships of pangasiid and schilbid catfish in Thailand. J. Fish Biol. 2012, 80, 2549–2570. [Google Scholar] [CrossRef]
- Chang, C.-H.; Lin, H.-Y.; Ren, Q.; Lin, Y.-S.; Shao, K.-T. DNA barcode identification of fish products in Taiwan: Government-commissioned authentication cases. Food Control 2016, 66, 38–43. [Google Scholar] [CrossRef]
- Vidthayanon, C. Taxonomic Revision of the Catfish Family Pangasiidae. Ph.D. Thesis, Tokyo University of Fisheries, Tokyo, Japan, 1993; 203p. [Google Scholar]
- Cacot, P.; Eeckhoutte, P.; Muon, D.T.; Trieu, N.V.; Legendre, M.; Mariojouls, C.; Lazard, J.M. Induced spermiation and milt management in Pangasius bocourti (Sauvage, 1880). Aquaculture 2003, 215, 67–77. [Google Scholar] [CrossRef]
- Poulsen, A.F.; Hortle, K.G.; Valbo-Jorgensen, J.; Chan, S.; Chhuon, C.K.; Viravong, S.; Bouakhamvongsa, K.; Suntornratana, U.; Yoorong, N.; Nguyen, T.T.; et al. Distribution and Ecology of Some Important Riverine Fish Species of the Mekong River Basin; MRC Technical Paper No. 10; MRC Secretariat: Vientiane, Laos, 2004. [Google Scholar]
- Haslawati, B. Morphometric, Phylogenetic Analyses and DNA Barcoding of Pangasiid Catfishes (Teleostei: Pangasiidae) in Peninsular Malaysia. Ph.D. Thesis, University of Malaya, Kuala Lumpur, Malaysia, 2006. [Google Scholar]
- Meier, R.; Shiyang, K.; Vaidya, G.; Ng, P.K.L. DNA barcoding and taxonomy in Diptera: A tale of high intraspecific variability and low identification success. Syst. Biol. 2006, 55, 715–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, C.P.; Paulay, G. DNA barcoding: Error rates based on comprehensive sampling. PLoS Biol. 2005, 12, e422. [Google Scholar] [CrossRef] [Green Version]
- Gustiano, R.; Teugels, G.G.; Pouyaud, L. Revision of the Pangasius kunyit catfish complex, with description of two new species from South-East Asia (Siluriformes; Pangasiidae). J. Nat. Hist. 2003, 37, 357–376. [Google Scholar] [CrossRef]
- Gauff, R.P.M.; Lejeusne, C.; Arsenieff, L.; Bohner, O.; Coudret, J.; Desbordes, F.; Jandard, A.; Loisel, S.; Schires, G.; Wafo, E.; et al. Alien vs. predator: Influence of environmental variability and predation on the survival of ascidian recruits of a native and alien species. Biol. Invasions 2022, 24, 1327–1344. [Google Scholar] [CrossRef]
- Chen, C.; Zheng, Z.; Wu, D.; Tan, L.; Yang, C.; Liu, S.; Lu, J.; Cheng, Y.; Sha, L.; Wang, Y.; et al. Morphological, cytological, and molecular evidences for natural hybridization between Roegneria stricta and Roegneria turczaninovii (Triticeae: Poaceae). Ecol. Evol. 2022, 12, e8517. [Google Scholar] [CrossRef]
- Gozlan, R.E.; Britton, J.R.; Cowx, I.; Copp, G.H. Current knowledge on non-native freshwater fish introductions. J. Fish Biol. 2010, 76, 751–786. [Google Scholar] [CrossRef]
- De Silva, S.S.; Nguyen, T.T.T.; Abery, N.W.; Amarasinghe, U.S. An evaluation of the role and impacts of alien finfish in Asian inland aquaculture. Aquac. Res. 2006, 37, 1–17. [Google Scholar] [CrossRef]
- Saidin, T.; Othman, A.A.; Mohamad-Zaini, S. Induced spawning techniques practised at Batu Berendam, Melaka, Malaysia. Aquaculture 1988, 74, 23–33. [Google Scholar]
- Pegg, J.; South, J.; Hill, J.E.; Durland-Donahou, A.; Weyl, O.L.F. Impacts of alien invasive species on large wetlands. Fundam. Trop. Freshw. Wetl. 2022, 1, 87–516. [Google Scholar]
- De Silva, S.S.; Nguyen, T.T.T.; Turchini, G.M.; Amarasinghe, U.S.; Abery, N.W. Alien species in aquaculture and biodiversity: A paradox in food production. Ambio 2009, 38, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Froese, R.; Pauly, D. (Eds.) FishBase. World Wide Web Electronic Publication. version (12/2019). 2019. Available online: www.fishbase.org (accessed on 15 January 2023).
- Crumlish, M.; Dung, T.T.; Turnbull, J.F.; Ngoc, N.T.N.; Ferguson, H.W. Identification of Edwardsiella ictaluri from diseased freshwater catfish, Pangasius hypophthalmus (Sauvage), cultured in the Mekong Delta, Vietnam. J. Fish Dis. 2002, 25, 733–736. [Google Scholar] [CrossRef]
- Faruk, M.A.R. Disease and health management of farmed exotic catfish Pangasius hypophthalmus in Mymensingh district of Bangladesh. In Diseases in Asian Aquaculture VI. Fish Health Section; Bondad-Reantaso, M.G., Mohan, C.V., Crumlish, M., Subasinghe, R.P., Eds.; Asian Fisheries Society: Manila, Philippines, 2008; pp. 193–204. 505p. [Google Scholar]
- Williams, M.; Hernandez-Jover, M.; Shamsi, S. Parasites of zoonotic interest in selected edible freshwater fish imported to Australia. Food Waterborne Parasitol. 2022, 26, e00138. [Google Scholar] [CrossRef]
- Siti-Zahrah, A.; Zamri-Saad, M.; Firdaus-Nawi, M.; Hazreen-Nita, M.K.; Nur-Nazifah, M. Detection of channel catfish virus in cage-cultured Pangasius hypophthalmus (Sauvage, 1878) in Malaysia. J. Fish Dis. 2014, 37, 981–983. [Google Scholar] [CrossRef]
- Reed, C. Import Risk Analysis: Frozen, Skinless and Boneless Fillet Meat of Pangasius spp. Fish Form Vietnam for Human Consumption; Biosecurity New Zealand, Ministry of Agriculture and Forestry: Wellington, New Zealand, 2008. Available online: http://www.biosecurity.govt.nz/files/regs/imports/risk/pangasius-risk-analysis.pdf (accessed on 28 November 2022).
- Székely, C.; Shaharom-Harrison, F.; Cech, G.; Mohamed, K.; Molnár, K. Myxozoan pathogens of Malaysian fishes cultured in ponds and net-cages. Dis. Aquat. Org. 2009, 83, 49–57. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sampling Location | Country | Accession Number@ | BOLD Systems | Specimen Voucher | Reference | |
|---|---|---|---|---|---|---|---|
| Sequence ID | BIN ID | ||||||
| Helicophagus waandersii | Lubok Paku, Pahang | Malaysia | KP036415 ** | GBMIN93860-17 | BOLD:AAE7042 | Hewa03LP | This study |
| KP036416 ** | GBMIN128296-17 | BOLD:ACI8082 | Hewa07LP | ||||
| H. waandersii | Paloh Hinai, Pahang | Malaysia | KP036417 ** | ANGBF43735-19 | BOLD:ACI8082 | Hewa24PH | This study |
| Pangasianodon hypophthalmus | Kuala Kangsar, Perak | Malaysia | KP036425 ** | ANGBF43738-19 | BOLD:AAE3237 | Pahy0901KK | This study |
| P. hypophthalmus | Paloh Hinai, Pahang | Malaysia | KP036426 * | ANGBF43739-19 | BOLD:AAE3237 | Pahy60PH | This study |
| P. hypophthalmus | Laguna, Calabarzon | Philippines | HQ682713-16 | NA | NA | Phyp1-Phyp4-LdB | [36] |
| P. hypophthalmus | NA | NA | NC021752 | NA | NA | NA | [37] |
| P. hypophthalmus | Nakhon Ratchasima | Thailand | JF292393 | ANGBF8216-12 | BOLD:AAE3237 | AUPH1 | [38] |
| JF292394 | ANGBF8282-12 | BOLD:AAE3237 | AUPH2 | ||||
| JF292395 | ANGBF8215-12 | BOLD:AAE3237 | AUPH3 | ||||
| JF292396 | ANGBF8281-12 | BOLD:AAE3237 | AUPH4 | ||||
| JF292397 | ANGBF8214-12 | BOLD:AAE3237 | AUPH5 | ||||
| JF292398 | ANGBF8280-12 | BOLD:AAE3237 | AUPH7 | ||||
| JF292399 | ANGBF8213-12 | BOLD:AAE3237 | AUPH8 | ||||
| JF292400 | ANGBF8279-12 | BOLD:AAE3237 | AUPH10 | ||||
| JF292401 | ANGBF8212-12 | BOLD:AAE3237 | AUPH11 | ||||
| JF292402 | ANGBF43753-19 | BOLD:AAE3237 | AUPH12 | ||||
| JF292403 | ANGBF43753-19 | BOLD:ADW5681 | AUPH13 | ||||
| JF292404 | ANGBF43753-19 | BOLD:AAE3237 | AUPH14 | ||||
| JF292405 | ANGBF43753-19 | BOLD:ADW5681 | AUPH15 | ||||
| JF292406 | ANGBF43753-19 | BOLD:AAE3237 | AUPH16 | ||||
| JF292407 | ANGBF8209-12 | BOLD:AAE3237 | AUPH17 | ||||
| JF292408 | ANGBF43753-19 | BOLD:AAE3237 | AUPH18 | ||||
| JF292409 | ANGBF43753-19 | BOLD:AAE3237 | AUPH19 | ||||
| JF292410 | ANGBF43753-19 | BOLD:AAE3237 | AUPH20 | ||||
| P. hypophthalmus | NA | NA | EU752151 | NA | NA | PANGHYPO-J01-009 | [39] |
| P. hypophthalmus | An Giang | Vietnam | EF609427 | NA | NA | BW-1778 | [40] |
| Pangasianodon bocourti | Paloh Hinai, Pahang | Malaysia | KP036428 * | GBMIN118560-17 | BOLD:AAB7484 | Pabo55PH | This study |
| P. bocourti | NA | NA | EU752149 | NA | NA | PANGBOCO-J01-003 | [40] |
| P. bocourti | An Giang | Viet Nam | EF 609425 | NA | NA | BW-1791 | [40] |
| P. bocourti | Yasothon Province | Thailand | JF292411 | ANGBF8207-12 | BOLD:AAB7484 | AUPH19 | [38] |
| JF292412 | ANGBF8273-12 | BOLD:AAB7484 | AUPH20 | ||||
| JF292413 | ANGBF8206-12 | BOLD:AAB7484 | AUPB2 | ||||
| JF292414 | ANGBF8272-12 | BOLD:AAB7484 | AUPB4 | ||||
| JF292415 | ANGBF8205-12 | BOLD:AAB7484 | AUPB5 | ||||
| JF292416 | ANGBF8271-12 | BOLD:AAB7484 | AUPB6 | ||||
| JF292417 | ANGBF8204-12 | BOLD:AAB7484 | AUPB7 | ||||
| JF292418 | ANGBF8270-12 | BOLD:AAB7484 | AUPB9 | ||||
| JF292419 | ANGBF8203-12 | BOLD:AAB7484 | AUPB10 | ||||
| JF292420 | ANGBF8269-12 | BOLD:AAB7484 | AUPB11 | ||||
| JF292421 | ANGBF8202-12 | BOLD:AAB7484 | AUPB12 | ||||
| JF292422 | ANGBF8268-12 | BOLD:AAB7484 | AUPB13 | ||||
| JF292423 | ANGBF8201-12 | BOLD:AAB7484 | AUPB14 | ||||
| JF292424 | ANGBF8267-12 | BOLD:AAB7484 | AUPB15 | ||||
| JF292425 | ANGBF8200-12 | BOLD:AAB7484 | AUPB16 | ||||
| JF292426 | ANGBF8266-12 | BOLD:AAB7484 | AUPB17 | ||||
| JF292427 | ANGBF8199-12 | BOLD:AAB7484 | AUPB18 | ||||
| JF292428 | ANGBF8265-12 | BOLD:AAB7484 | AUPB19 | ||||
| JF292429 | ANGBF8198-12 | BOLD:AAB7484 | AUPB20 | ||||
| Pangasianodon conchophilus | Paloh Hinai, Pahang | Malaysia | KP036413 * | GBMIN128295-17 | BOLD:AAE7042 | Paco23PH | This study |
| P. conchophilus | Chenor, Pahang | Malaysia | KP036414 ** | GBMIN118559-17 | BOLD:AAE7042 | Paco03CR | This study |
| P. conchophilus | NA | Vietnam | EF609426 | NA | NA | BW-1796 | [40] |
| Pangasianodon djambal | Paloh Hinai, Pahang | Malaysia | KP036427 * | GBMIN128301-17 | BOLD:AAB7484 | Padj53PH | This study |
| P. nasutus | Paloh Hinai, Pahang | Malaysia | KP036410 * | GBMIN123337-17 | BOLD:AAE7042 | Pana57PH | This study |
| KP036411 * | GBMIN93860-17 | BOLD:AAE7042 | Pana61PH | ||||
| KP036412 * | GBMIN123338-17 | BOLD:AAE7042 | Pana93PH | ||||
| P. nasutus | Pahang | Malaysia | JF781172 | NA | NA | SLM-PN(PH)-01 | [34] |
| JF781173 | NA | NA | SLM-PN(PH)-02 | ||||
| JF781174 | NA | NA | SLM-PN(PH)-03 | ||||
| JF781175 | NA | NA | SLM-PN(PH)-04 | ||||
| Pangasianodon micronemus | Kuala Kangsar, Perak | Malaysia | KP036418 ** | GBMIN123340-17 | BOLD:AAU2068 | Pami0801KK | This study |
| P. micronemus | Paloh Hinai, Pahang | Malaysia | KP036419 * | GBMIN128297-17 | BOLD:AAU2068 | Pami50PH | This study |
| KP036420 * | GBMIN128298-17 | BOLD:AAU2068 | Pami51PH | ||||
| KP036421 * | GBMIN128299-17 | BOLD:AAU2068 | Pami52PH | ||||
| KP036422 * | GBMIN128300-17 | BOLD:AAU2068 | Pami59PH | ||||
| KP036423 * | GBMIN123341-17 | BOLD:AAU2068 | Pami85PH | ||||
| P. micronemus | Kuala Kangsar, Perak | Malaysia | KP036424 ** | GBMIN123342-17 | BOLD:AAU2068 | Pami0902KK | This study |
| P. micronemus | Pahang | Malaysia | HM156360 | NA | NA | SLM-PM(PH)-01 | [34] |
| HM156361 | NA | NA | SLM-PM(PH)-02 | ||||
| HM156362 | NA | NA | SLM-PM(PH)-03 | ||||
| HM156363 | NA | NA | SLM-PM(PH)-04 | ||||
| HM156364 | NA | NA | SLM-PM(PH)-05 | ||||
| H. macropterus | Hechuan, Chongqing | China | NC019592 | NA | NA | NA | [41] |
| Clarias macrocephalus | Kasetsart University | Thailand | JF292337 | ANGBF8244-12 | BOLD:AAE8721 | AUCM19 | [38] |
| C. batrachus | Nakhon Ratchasima | Thailand | JF292297 | ANGBF8264-12 | BOLD:ACB6804 | AUCB1 | [38] |
| C. batrachus | Laguna, Calabarzon | Philippines | HQ682679 | NA | NA | Cbat3-LdB | [36] |
| Species | 1 | 2 | 3 | 4 | 5 | 6 | d |
|---|---|---|---|---|---|---|---|
| Pangasius nasutus | - | 0.011 | |||||
| P. conchophilus | 0.030 (3.0) | - | 0.051 | ||||
| P. bocourti | 0.120 (12.0) | 0.111 (11.1) | - | 0.012 | |||
| P. djambal | 0.065 (6.5) | 0.083 (8.3) | 0.071 (7.1) | - | NC | ||
| Pangasianodon hypophthalmus | 0.157 (15.7) | 0.144 (14.4) | 0.117 (11.7) | 0.163 (16.3) | - | 0.014 | |
| Pseudolais micronemus | 0.077 (7.7) | 0.097 (9.7) | 0.133 (13.3) | 0.074 (7.4) | 0.166 (16.6) | - | 0.008 |
| Helicophagus waandersii | 0.113 (11.3) | 0.129 (12.9) | 0.168 (16.8) | 0.097 (9.7) | 0.197 (19.7) | 0.090 (9.0) | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haslawati, B.; Amatul-Samahah, M.A.; Rizman-Idid, M.; Muniandy, S. DNA Barcoding and Phylogenetics Relationship of Pangasiid Catfishes in Peninsular Malaysia Revealed the Impacts of Aquaculture on the Native Species Conservation. Hydrobiology 2023, 2, 431-445. https://doi.org/10.3390/hydrobiology2020028
Haslawati B, Amatul-Samahah MA, Rizman-Idid M, Muniandy S. DNA Barcoding and Phylogenetics Relationship of Pangasiid Catfishes in Peninsular Malaysia Revealed the Impacts of Aquaculture on the Native Species Conservation. Hydrobiology. 2023; 2(2):431-445. https://doi.org/10.3390/hydrobiology2020028
Chicago/Turabian StyleHaslawati, Baharuddin, Md Ali Amatul-Samahah, Mohammed Rizman-Idid, and Sekaran Muniandy. 2023. "DNA Barcoding and Phylogenetics Relationship of Pangasiid Catfishes in Peninsular Malaysia Revealed the Impacts of Aquaculture on the Native Species Conservation" Hydrobiology 2, no. 2: 431-445. https://doi.org/10.3390/hydrobiology2020028