
Recurringly Hypoxic: Bottom Water Oxygen Depletion Is Linked to Temperature and Precipitation in a Great Lakes Estuary



Abstract
:1. Introduction
2. Materials and Methods
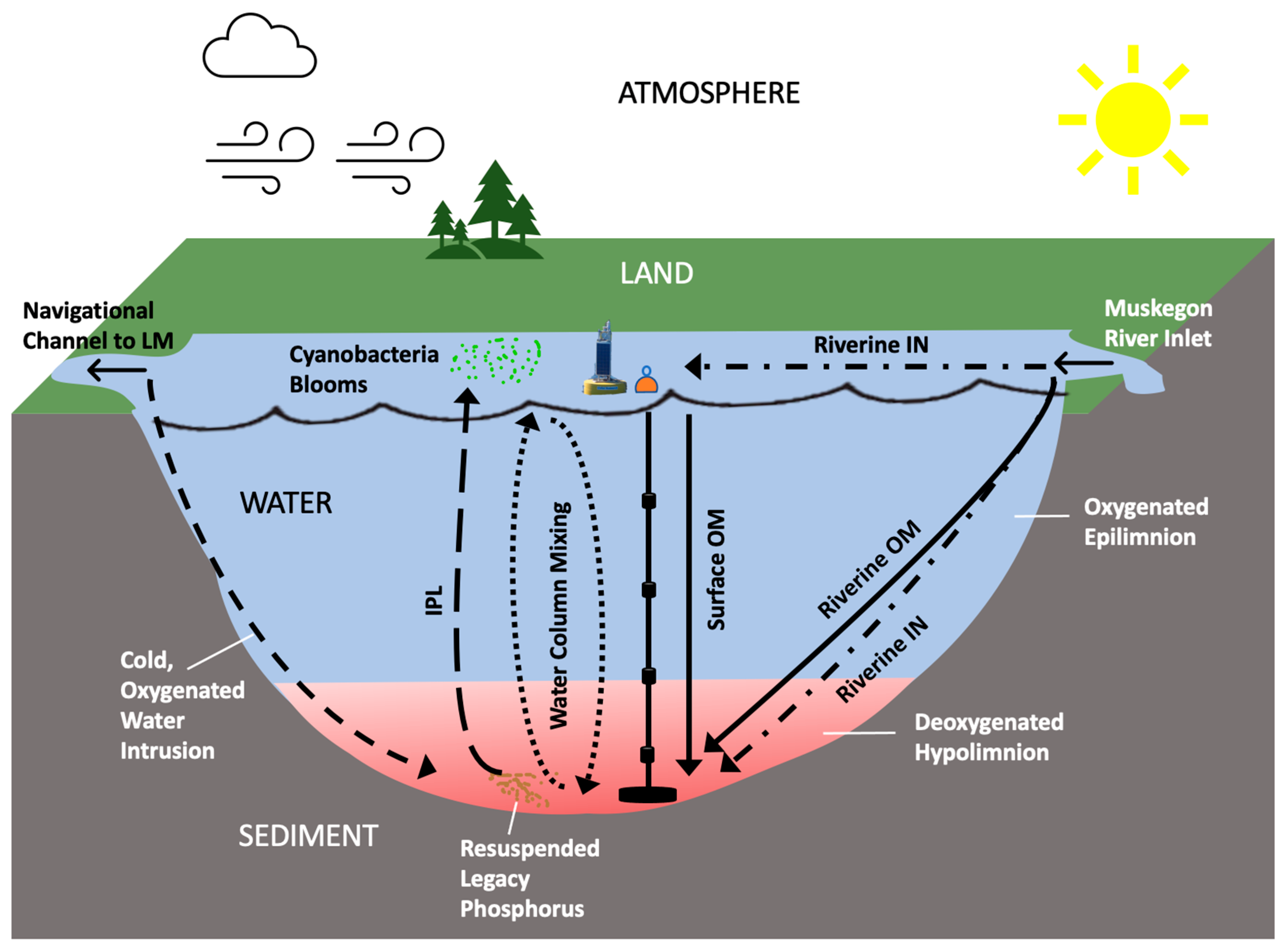
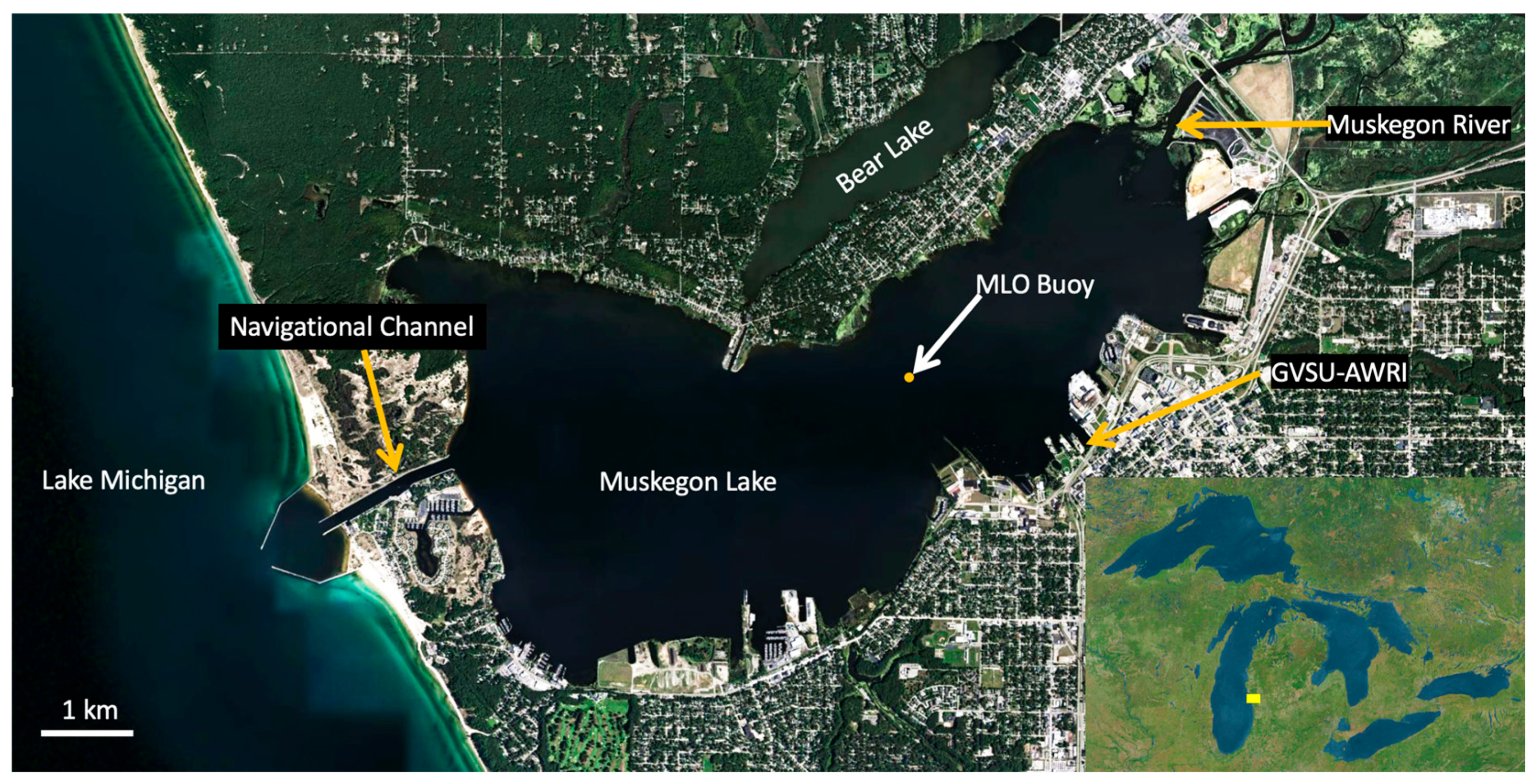
2.1. Study Site
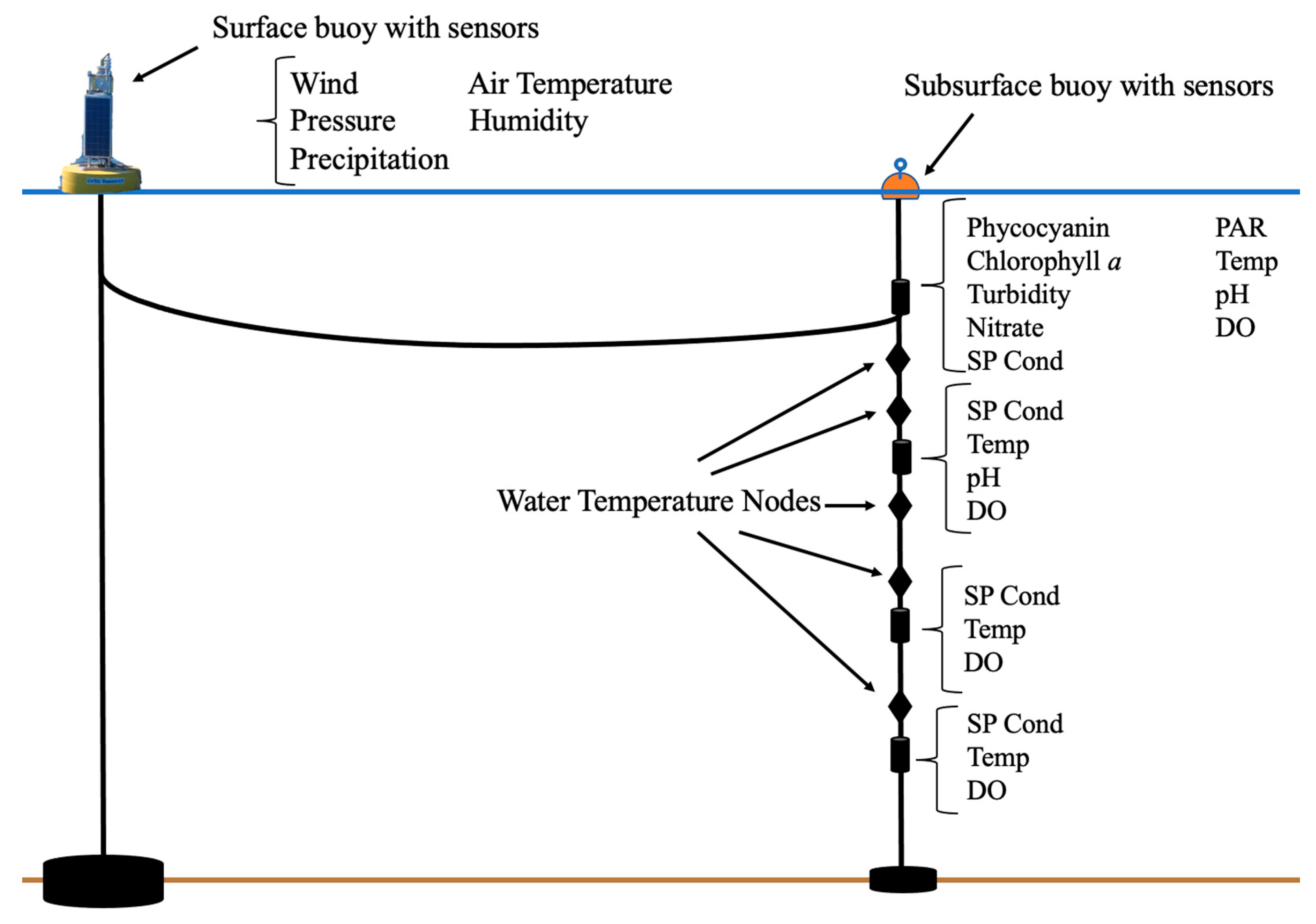
2.2. Muskegon Lake Observatory Data Collection
2.3. Data Visualization and Calculations
2.4. Hypoxic Severity Index
2.5. Schmidt Stability
3. Results
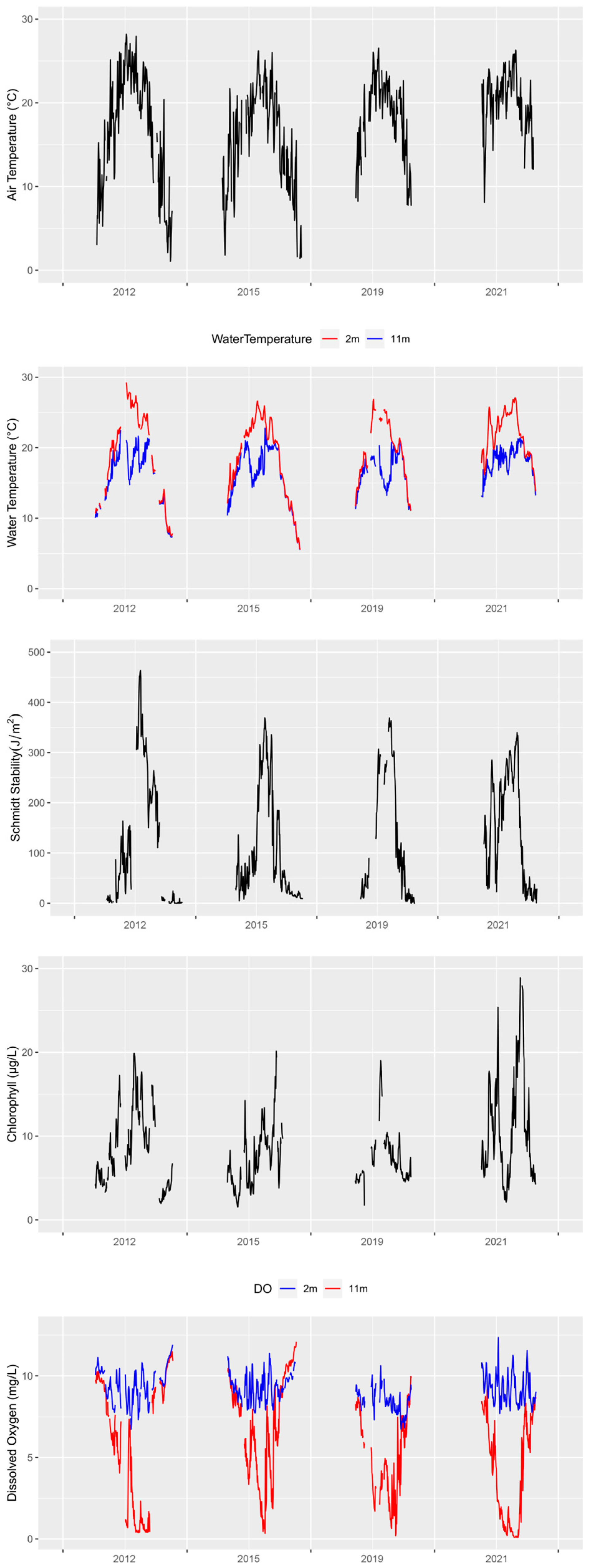
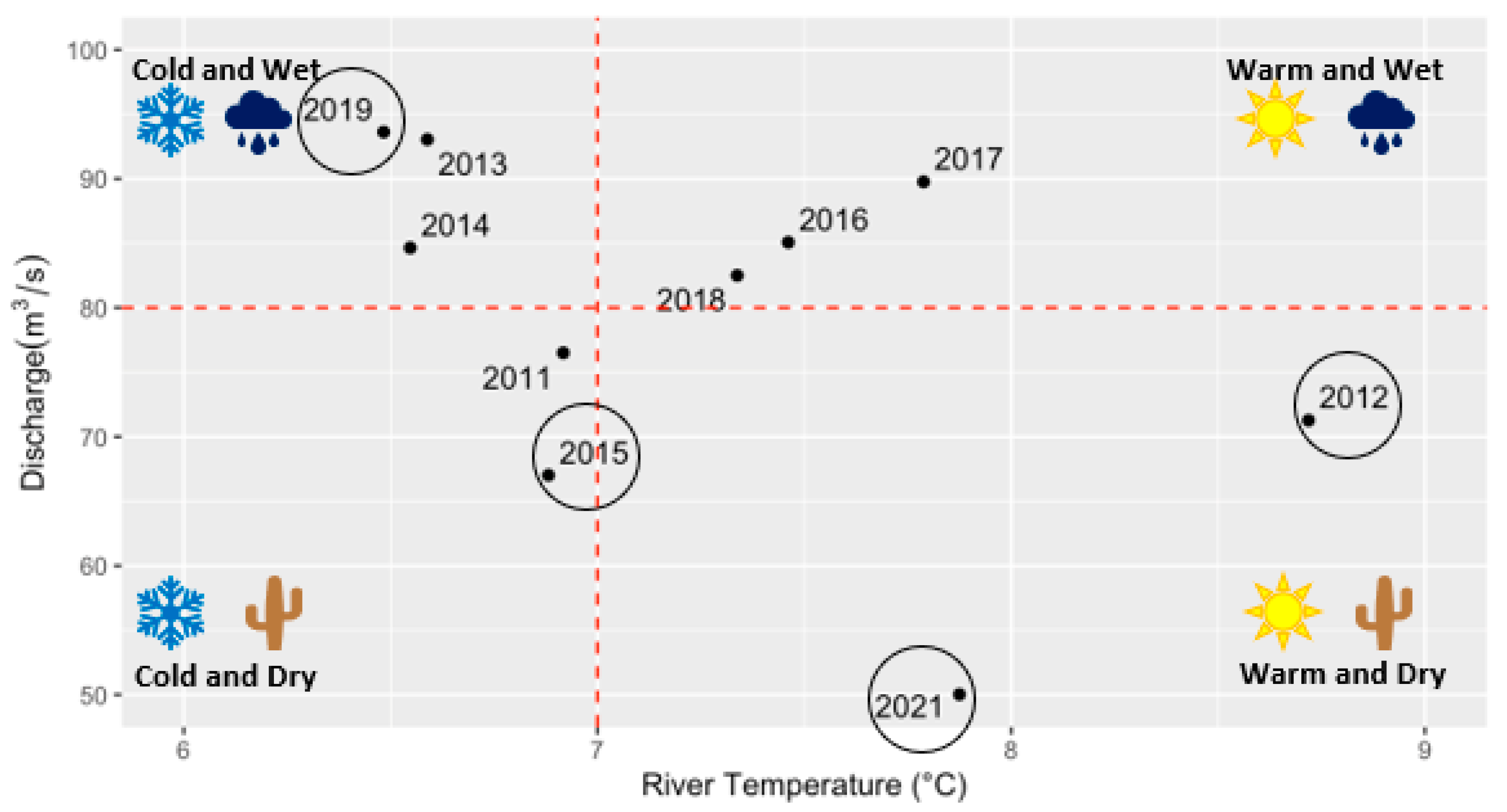
3.1. Environmental Drivers
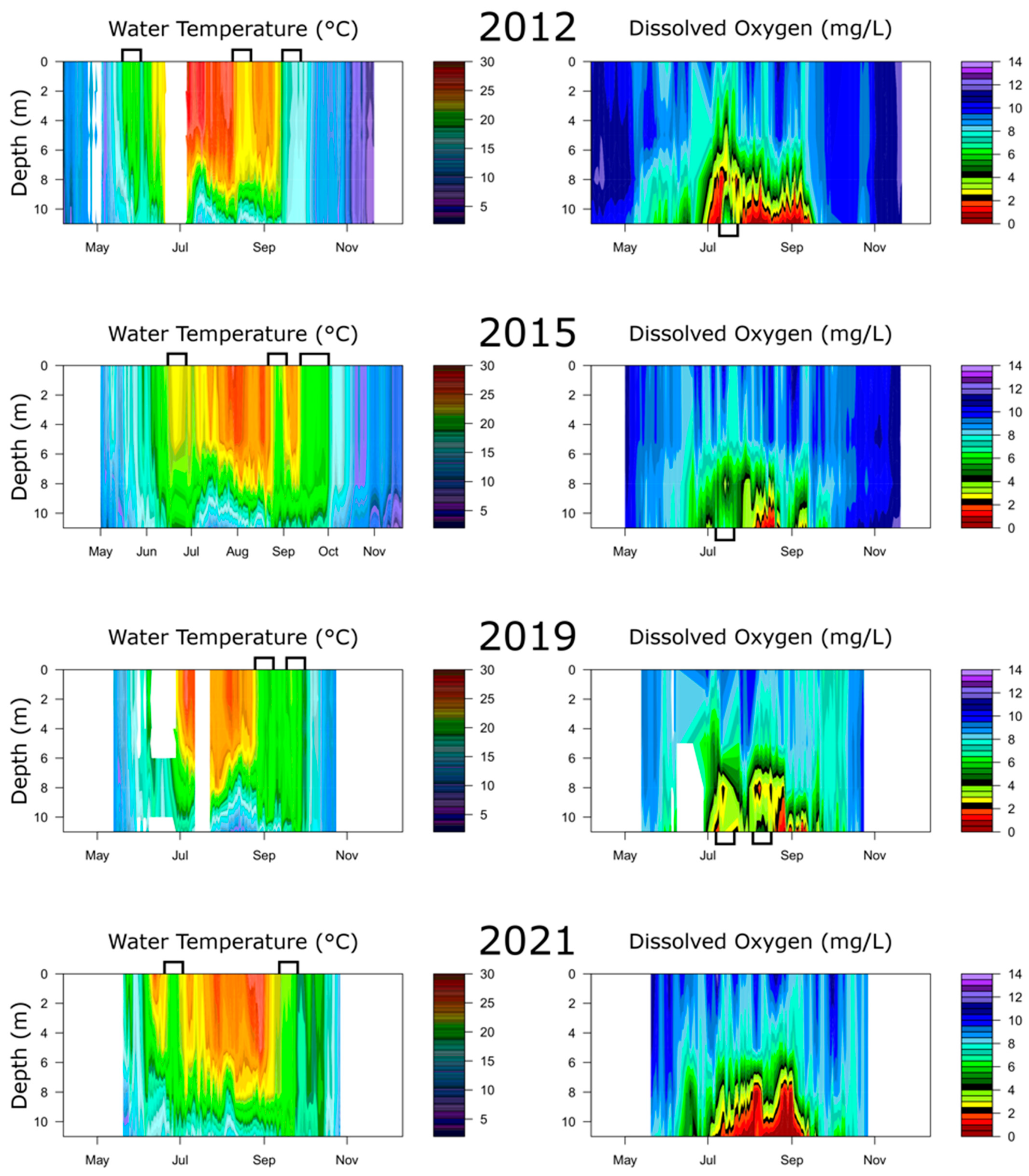
3.2. Comparison of Heat Maps
3.3. Comparison of Stratification Strength and Duration
3.4. Hypoxia Severity Index
4. Discussion
4.1. Variable Inter- and Intra-Annual Hypoxia
4.2. Temperature-Precipitation Regime Regulates Hypoxia
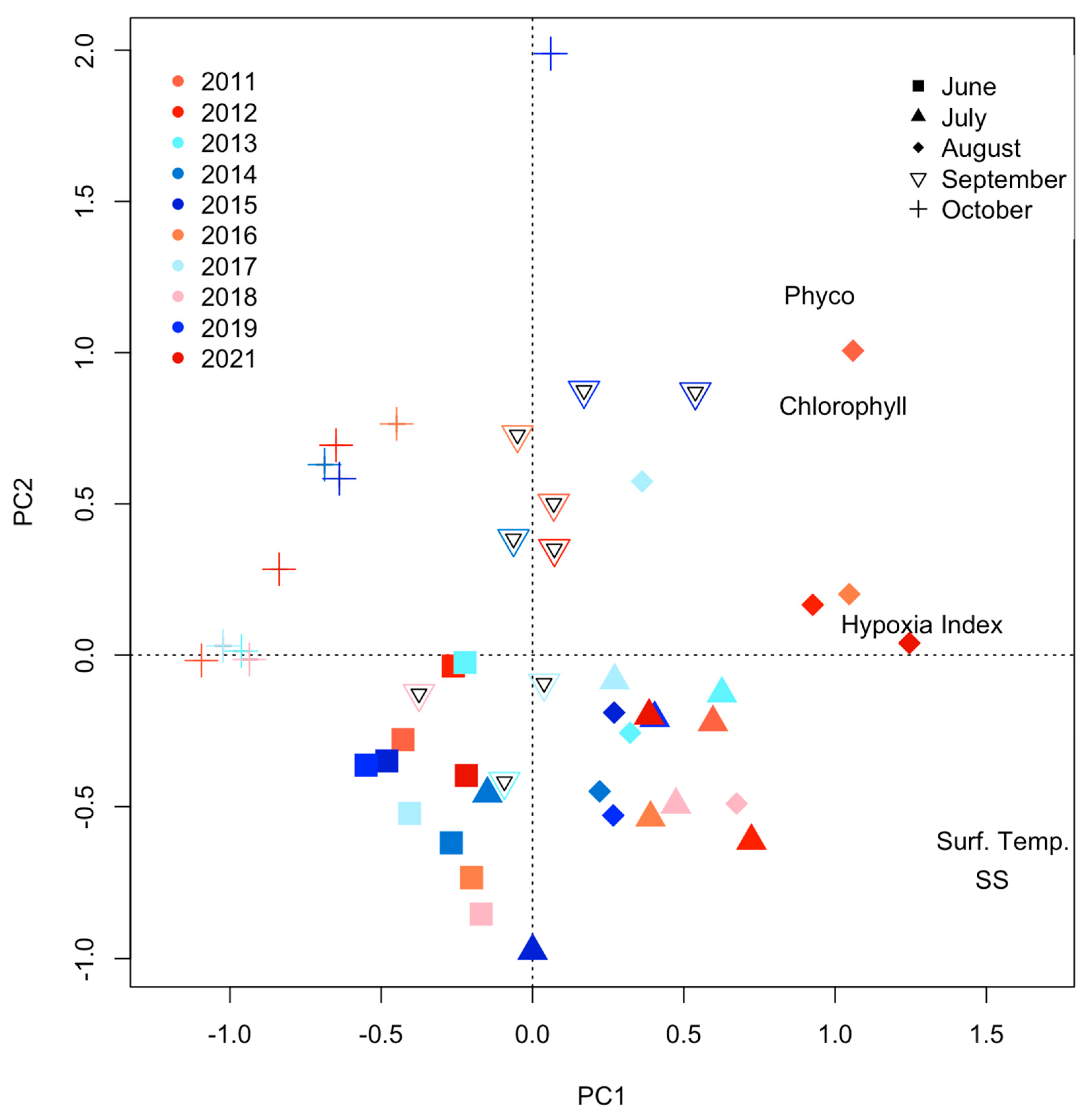
4.3. Role of Surface Production and Hypolimnetic Respiration
4.4. Potential Release and Role of Legacy Phosphorus in Sediment
4.5. Complexities of Hypoxia Dynamics and Management Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Breitburg, D.; Levin, L.A.; Oschlies, A.; Grégoire, M.; Chavez, F.P.; Conley, D.J.; Garçon, V.; Gilbert, D.; Gutiérrez, D.; Isensee, K.; et al. Declining oxygen in the global ocean and coastal waters. Science 2018, 359, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, R.J.; Rosenberg, R. Spreading dead zones and consequences for marine ecosystems. Science 2008, 231, 926–929. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Chen, M.; Gong, M.; Fan, X.; Qin, B.; Xu, H.; Gao, S.; Jin, Z.; Tsang, D.C.W.; Zhang, C. Internal phosphorus loading from sediments causes seasonal nitrogen limitation for harmful algal blooms. Sci. Total Environ. 2018, 625, 872–884. [Google Scholar] [CrossRef] [PubMed]
- Jane, S.F.; Hansen, G.J.A.; Kraemer, B.M.; Leavitt, P.R.; Mincer, J.L.; North, R.L.; Pilla, R.M.; Stetler, J.T.; Williamson, C.E.; Woolway, R.I.; et al. Widespread deoxygenation of temperate lakes. Nature 2021, 594, 66–81. [Google Scholar] [CrossRef]
- Rabalais, N.N.; Díaz, R.J.; Levin, L.A.; Turner, R.E.; Gilbert, D.; Zhang, J. Dynamics and distribution of natural and human-caused hypoxia. Biogeosciences 2010, 7, 585–619. [Google Scholar] [CrossRef] [Green Version]
- Scavia, D.; Allan, J.D.; Arend, K.K.; Bartell, S.; Beletsky, D.; Bosch, N.S.; Brandt, S.B.; Briland, R.D.; Daloğlu, I.; DePinto, J.V.; et al. Assessing and addressing the re-eutrophication of Lake Erie: Central basin hypoxia. J. Great Lakes Res. 2014, 40, 226–246. [Google Scholar] [CrossRef]
- Weinke, A.D.; Biddanda, B.A. From bacteria to fish: Ecological consequences of seasonal hypoxia in a Great Lakes estuary. Ecosystems 2018, 21, 426–442. [Google Scholar] [CrossRef] [Green Version]
- Magaud, H.; Migeon, B.; Morfin, P.; Garric, J.; Vindimian, E. Modelling fish mortality due to urban storm run-off: Interacting effects of hypoxia and unionized ammonia. Water Res. 1997, 31, 211–218. [Google Scholar] [CrossRef]
- Tellier, J.M.; Kalejs, N.I.; Leonhardt, B.S.; Cannon, D.; Höök, T.O.; Collingsworth, P. Widespread prevalence of hypoxia and the classification of hypoxic conditions in the Laurentian Great Lakes. J. Great Lakes Res. 2022, 48, 13–23. [Google Scholar] [CrossRef]
- Biddanda, B.A.; Weinke, A.D.W.; Kendall, S.T.; Gereaux, L.C.; Holcomb, T.M.; Snider, M.J.; Dila, D.K.; Long, S.A.; VandenBurg, C.; Knapp, K.; et al. Chronicles of hypoxia: Time-series buoy observations reveal annually recurring seasonal basin-wide hypoxia in Muskegon Lake—A Great Lakes estuary. J. Great Lakes Res. 2018, 44, 219–229. [Google Scholar] [CrossRef]
- Xu, W.; Collingsworth, P.D.; Kraus, R.; Minsker, B. Spatio-Temporal Analysis of Hypoxia in the Central Basin of Lake Erie of North America. Water Resour. Res. 2021, 57, e2020WR027676. [Google Scholar] [CrossRef]
- Klump, J.V.; Brunner, S.L.; Grunert, B.K.; Kaster, J.L.; Weckerly, K.; Houghton, E.M.; Kennedy, J.A.; Valenta, T.J. Evidence of persistent, recurring summertime hypoxia in Green Bay, Lake Michigan. J. Great Lakes Res. 2018, 44, 841–850. [Google Scholar] [CrossRef]
- Larson, J.H.; Trebitz, A.S.; Steinman, A.D.; Wiley, M.J.; Carlson Mazur, M.; Pebbles, V.; Braun, H.A.; Seelbach, P.W. Great Lakes rivermouth ecosystems: Scientific synthesis and management implications. J. Great Lakes Res. 2013, 39, 513–524. [Google Scholar] [CrossRef]
- Weinke, T.; Stone, I.; Greene, J.; Woznicki, S.; Cook, J.; Dugener, N.; Biddanda, B. Great Lakes Estuaries: Hot spots of productivity, problems, and potential. IAGLR Lakes Lett. 2022, 9–10. Available online: https://iaglr.org/ll/2022-3-Summer_LL14.pdf (accessed on 30 October 2022).
- Steinman, A.D.; Ogdahl, M.; Rediske, R.; Ruetz, C.R.; Biddanda, B.A.; Nemeth, L. Current status and trends in Muskegon Lake, Michigan. J. Great Lakes Res. 2008, 34, 169–188. [Google Scholar] [CrossRef]
- Rahmstorf, S.; Coumou, D. Increase of extreme events in a warming world. Proc. Natl. Acad. Sci. USA 2011, 108, 17905–17909. [Google Scholar] [CrossRef] [Green Version]
- Byun, K.; Hamlet, A.F. Projected changes in future climate over the Midwest and Great Lakes region using downscaled CMIP5 ensembles. Int. J. Climatol. 2018, 38, e531–e553. [Google Scholar] [CrossRef]
- Michalak, A.M. Study role of climate change in extreme threats to water quality. Nature 2016, 535, 349–350. [Google Scholar] [CrossRef] [Green Version]
- Whitney, M.M. Observed and forecasted global warming pressure on coastal hypoxia. Biogeosciences 2021, 19, 4479–4497. [Google Scholar]
- O’Reilly, C.M.; Sharma, S.; Gray, D.K.; Hampton, S.E.; Read, J.S.; Rowley, R.J.; Schneider, P.; Lenters, J.D.; McIntyre, P.B.; Kraemer, B.M.; et al. Rapid and highly variable warming of lake surface waters around the globe. AGUPublications 2015, 42, 10773–10781. [Google Scholar] [CrossRef] [Green Version]
- Tassone, S.J.; Besterman, A.F.; Buelo, C.D.; Ha, D.T.; Walter, J.A.; Pace, M.L. Increasing heatwave frequency in streams and rivers of the United States. Limnol. Oceanogr. 2023, 8, 295–304. [Google Scholar] [CrossRef]
- Wallace, R.B.; Gobler, C.J. The role of algal blooms and community respiration in controlling the temporal and spatial dynamics of hypoxia and acidification in eutrophic estuaries. Marine Pollut. Bull. 2021, 172, 112908. [Google Scholar] [CrossRef] [PubMed]
- Austin, J.A.; Colman, S.M. Lake Superior summer water temperatures are increasing more rapidly than regional air temperatures: A positive ice-albedo feedback. Geophys. Res. Lett. 2007, 34, L06604. [Google Scholar] [CrossRef] [Green Version]
- Winslow, L.A.; Read, J.S.; Hansen, G.J.A.; Rose, K.C.; Robertson, D.M. Seasonality of change: Summer warming rates do not fully represent effects of climate change on lake temperatures. Limno. Oceanogr. 2017, 62, 2168–2178. [Google Scholar] [CrossRef]
- Smith, V.H. Eutrophication of freshwater and coastal marine ecosystems, a global problem. Environ. Sci. Pollut. Res. 2003, 10, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Valdes, L.M.; Peierls, B.L.; Adolf, J.E.; Harding, L.W., Jr. Anthropogenic and climatic influences on the eutrophication of large estuarine ecosystems. Limnol. Oceanogr. 2006, 51, 448–462. [Google Scholar] [CrossRef] [Green Version]
- Paerl, H.W.; Gardner, W.S.; Havens, K.E.; Joyner, A.R.; McCarthy, M.J.; Newell, S.E.; Qin, B.; Scott, J.T. Mitigating cyanobacterial harmful algal blooms in aquatic ecosystems impacted by climate change and anthropogenic nutrients. Harmful Algae 2016, 54, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Marko, K.M.; Rutherford, E.S.; Eadie, B.J.; Johengen, T.H.; Lansing, M.B. Delivery and seston from the Muskegon River Watershed to nearshore Lake Michigan. J. Great Lakes Res. 2013, 39, 672–681. [Google Scholar] [CrossRef]
- Dodds, W.K.; Bouska, W.W.; Eitzmann, J.L.; Pilger, T.J.; Pitts, K.L.; Riley, A.J.; Schloesser, J.T.; Thornbrugh, D.L. Eutrophication of U.S. freshwaters: Analysis of potential economic damages. Environ. Sci. Technol. 2008, 43, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Salk, K.R.; Venkitswaran, J.J.; Couture, R.; Higgins, S.N.; Paterson, M.J.; Schiff, S.L. Warming combined with experimental eutrophication intensifies lake phytoplankton blooms. Limnol. Oceanogr. 2021, 67, 147–158. [Google Scholar] [CrossRef]
- Burkholder, J.M.; Kinder, C.A.; Dickey, D.A.; Reed, R.E.; Arellano, C.; James, J.L.; Mackenzie, L.M.; Allen, E.H.; Lindor, N.L.; Mathis, J.G.; et al. Classic indicators and diel dissolved oxygen versus trend analysis in assessing eutrophication of potable-water reservoirs. Ecol. Appl. 2022, 32, e2541. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Kim, K.H.; Hwang, S.J.; Lee, T.K. Role of Algal Community Stability in Harmful Algal Blooms in River-Connected Lakes. Microbial Ecol. 2021, 82, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Williamson, C.E.; Saros, J.E.; Vincent, W.F.; Smol, J.P. Lakes and reservoirs as sentinels, integrators, and regulators of climate change. Limnol. Oceanogr. 2009, 54, 2273–2282. [Google Scholar] [CrossRef]
- Freedman, P.; Canale, R.; Auer, M. The Impact of Wastewater Diversion Spray Irrigation on Water Quality in Muskegon County Lakes; U.S. Environmental Protection Agency: Washington, DC, USA, 1979; 905/9–79–006-A. [Google Scholar]
- Liu, Q.; Anderson, E.J.; Zhang, Y.; Weinke, A.D.; Knapp, K.L.; Biddanda, B.A. Modeling reveals the role of coastal upwelling and hydrologic inputs on biologically distinct water exchanges in a Great Lakes estuary. Estuar. Coast. Shelf Sci. 2018, 209, 41–55. [Google Scholar] [CrossRef]
- Biddanda, B.; Kendall, S.; Weinke, A.; Stone, I.; Dugener, N.; Ruberg, S.; Leidig, J.; Smith, E.; Berg, M.; Wolffe, G. Muskegon Lake Observatory Buoy Data: Muskegon Lake, Michigan: 2011–2019 ver 1. Environmental Data Initiative. Available online: https://doi.org/10.6073/pasta/d1ef6101a1870a0b0bd54d3914dc13dc (accessed on 30 October 2022).
- Mancuso, J.L.; Weinke, A.D.; Stone, I.P.; Hamsher, S.E.; Villar-Argaiz, M.; Biddanda, B.A. Cold and wet: Diatoms dominate the phytoplankton community during a year of anomalous weather in a Great Lakes estuary. J. Great Lakes Res. 2021, 47, 1305–1315. [Google Scholar] [CrossRef]
- Winslow, L.; Read, J.; Woolway, R.; Brentrup, J.; Leach, T.; Zwart, J.; Albers, S.; Collinge, D. rLakeAnalyzer: Lake Physics Tools, R package version 1 (11). 2015. Available online: https://rdrr.io/cran/rLakeAnalyzer/ (accessed on 30 October 2022).
- Read, J.S.; Hamilton, D.P.; Jones, I.D.; Muraoka, K.; Winslow, L.A.; Kroiss, R.; Wu, C.H.; Gaiser, E. Derivation of lake mixing and stratification indices from high-resolution lake buoy data. Environ. Model. Softw. 2011, 26, 1325–1336. [Google Scholar] [CrossRef]
- Nürnberg, G.K. Quantifying anoxia in lakes. Limnol. Oceanogr. 1995, 40, 1100–1111. [Google Scholar] [CrossRef]
- Matthews, D.A.; Effler, S.W. Long-term changes in the areal hypolimnetic oxygen deficit (AHOD) of Onondaga Lake: Evidence of sediment feedback. Limnol. Oceanogr. 2006, 51, 702–714. [Google Scholar] [CrossRef]
- Foley, B.; Jones, I.D.; Maberly, S.C.; Rippey, B. Long-term changes in oxygen depletion in a small temperate lake: Effects of climate change and eutrophication. Freshw. Biol. 2012, 57, 278–289. [Google Scholar] [CrossRef]
- Nürnberg, G.K. Quantification of Oxygen Depletion in Lakes and Reservoirs with the Hypoxic Factor. Lake Reserv. Manag. 2002, 18, 299–306. [Google Scholar] [CrossRef]
- USGS Muskegon River Near Croton, MI. Available online: https://waterdata.usgs.gov/monitoring-location/04121970/#parameterCode=00060&period=P365D. (accessed on 7 December 2022).
- Weinke, A.D.; Biddanda, B.A. Influence of episodic wind events on thermal stratification and bottom water hypoxia in a Great Lakes estuary. J. Great Lakes Res. 2019, 45, 1103–1112. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Wickham, H.; François, R.; Henry, L.; Müller, K. dplyr: A Grammar of Data Manipulation, R package version 0.7.5. 2018. Available online: https://cran.r-project.org/web/packages/dplyr/index.html (accessed on 30 October 2022).
- Verzani, J. UsingR: Data Sets, Etc. For the Test “Using R for Introductory Statistics”, R Package Version 2.0-6, 2nd ed. 2018. Available online: https://cran.r-project.org/web/packages/UsingR/UsingR.pdf (accessed on 30 October 2022).
- Defore, A.L.; Weinke, A.D.; Lindback, M.M.; Biddanda, B.A. Year-round measures of planktonic metabolism reveal net autotrophy in surface waters of a Great Lakes estuary. Aquat. Microb. Ecol. 2016, 77, 139–153. [Google Scholar] [CrossRef] [Green Version]
- Steinman, A.D.; Spears, B.M. Internal Phosphorus Loading in Lakes: Causes, Case Studies, and Management; J. Ross Publishing: Plantation, FL, USA, 2020. [Google Scholar]
- Schmid, M.; Read, J. Heat Budget of Lakes. Reference Module In Earth Systems and Environmental Sciences; Encyclopedia of Inland Waters: Amsterdam, The Netherlands, 2021; Volume 7. [Google Scholar]
- Salk, K.R.; Ostrom, P.H.; Biddanda, B.A.; Weinke, A.D.; Kendall, S.T.; Ostrom, N.E. Ecosystem metabolism and greenhouse gas production in a mesotrophic northern temperate lake experiencing seasonal hypoxia. Biogeochem. 2016, 131, 303–319. [Google Scholar] [CrossRef]
- Carey, C.C.; Hanson, P.C.; Thomas, R.Q.; Gerling, A.B.; Hounshell, A.G.; Lewis, A.S.L.; Lofton, M.E.; McClure, R.P.; Wander, H.L.; Woelmer, W.M.; et al. Anoxia decreases the magnitude of the carbon, nitrogen, and phosphorus sink in freshwaters. Global Chang. Biol. 2022, 28, 4861–4881. [Google Scholar] [CrossRef] [PubMed]
- Villar-Argaiz, M.; Medina-Sánchez, J.M.; Biddanda, B.A.; Carrillo, P. Predominant Non-additive Effects of Multiple Stressors on Autotroph C:N:P Ratios Propagate in Freshwater and Marine Food Webs. Front. Microbiol. 2018, 9, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheffer, M.; Carpenter, S.R.; Lenton, T.M.; Bascompte, J.; Brock, W.; Dakos, V.; Van De Koppel, J.; Van De Leemput, I.A.; Levin, S.A.; Van Nes, E.H.; et al. Anticipating Critical Transitions. Science 2012, 338, 344–348. [Google Scholar] [CrossRef] [Green Version]
- Scheffer, M.; Jeppesen, E. Alternative Stable States. Ecol. Studies 1998, 131, 397–406. [Google Scholar]
- Schneider, P.; Hook, S.J. Space observations of inland waterbodies show rapid surface warming since 1985. Geophys. Res. 2010, 37, L22405. [Google Scholar]
- Dokulil, M.T. Impact of climate warming on European inland waters. Inland Waters 2014, 4, 27–40. [Google Scholar] [CrossRef]
- Jenny, J.-P.; Francus, P.; Normandeau, A.; Lapointe, F.; Perga, M.-E.; Ojala, A.; Schimmelmann, A.; Zolitschka, B. Global spread of hypoxia in freshwater ecosystems during the last three centuries is caused by rising local human pressure. Global Chang. Biol. 2016, 22, 1481–1489. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Blooms like it hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meire, L.; Soetaert, K.E.R.; Meysman, F.J.R. Impact of global change on coastal oxygen dynamics and risk of hypoxia. Biogeosciences 2013, 10, 2633–2653. [Google Scholar] [CrossRef] [Green Version]
- Hayhoe, K.; VanDorn, J.; Croley, T.; Schlegal, N.; Wuebbles, D. Regional climate change projections for Chicago and the US Great Lakes. J. Great Lakes Res. 2010, 36, 7–21. [Google Scholar] [CrossRef]
- Hondzo, M.; Stefan, H.G. Three case studies of lake temperature and stratification response to warmer climate. Water Resour. Res. 1991, 27, 1837–1848. [Google Scholar] [CrossRef]
- Del Giudice, D.; Zhou, Y.; Sinha, E.; Michalak, A.M. Long-term phosphorous loading and springtime temperatures explain interannual variability of hypoxia in a large temperate lake. Environ. Sci. Technol. 2018, 52, 2046–2054. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, G.B.; Schladow, S.G.; Reuter, J.E.; Coats, R. Effects of climate change on thermal properties of lakes, reservoirs, and possible implications. Stoch. Environ. Res. Risk Assess. 2011, 25, 445–456. [Google Scholar] [CrossRef] [Green Version]
- Scully, M.E.; Geyer, W.R.; Borkman, D.; Pugh, T.L.; Costa, A.; Nichols, O.C. Unprecedented Summer Hypoxia in Southern Cape Cod Bay: An Ecological Response to Regional Climate Change? Biogeosciences 2022, 19, 3523–3536. [Google Scholar] [CrossRef]
- Pal, I.; Anderson, B.T.; Salvucci, G.D.; Gianatti, D.J. Shifting seasonality and increasing frequency of precipitation in wet and dry seasons across the U.S. Geophys. Res. Lett. 2013, 40, 4030–4035. [Google Scholar] [CrossRef]
- Crockford, L.; Jordan, P.; Melland, A.R.; Taylor, D. Storm-triggered, increased-supply of sediment-derived phosphorus to the epilimnion in a small freshwater lake. Inland Waters 2015, 5, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Bouffard, D.; Ackerman, J.D.; Boegman, L. Factors affecting the development and dynamics of hypoxia in a large shallow stratified lake: Hourly to seasonal patterns. Water Resour. Res. 2013, 49, 2380–2394. [Google Scholar] [CrossRef] [Green Version]
- Fennel, K.; Testa, J.M. Biogeochemical Controls on Coastal Hypoxia. Annu. Rev. Mar. Sci. 2019, 11, 105–130. [Google Scholar] [CrossRef] [PubMed]
- Watson, S.B.; Miller, C.; Arhonditsis, G.; Boyer, G.L.; Carmichael, W.; Charlton, M.N.; Confesor, R.; Depew, D.C.; Höök, T.O.; Ludsin, S.A.; et al. The re-eutrophication of Lake Erie: Harmful algal blooms and hypoxia. Harmful Algae 2016, 56, 44–66. [Google Scholar] [CrossRef]
- Edwards, W.J.; Conroy, J.D.; Culver, D.A. Hypolimnetic Oxygen Depletion Dynamics in the Central Basin of Lake Erie. J. Great Lakes Res. 2005, 31, 262–271. [Google Scholar] [CrossRef]
- Harp, R.D.; Horton, D.E. Observed changes in daily precipitation intensity in the United States. Geophys. Res. Lett. 2022, 49, e2022GL099955. [Google Scholar] [CrossRef]
- Deng, J.; Qin, B.; Paerl, H.; Zhang, Y.; Ma, J.; Chen, Y. Earlier and warmer springs increase cyanobacterial (Microcystis spp.) blooms in subtropical Lake Taihu, China. Freshw. Biol. 2014, 59, 1076–1085. [Google Scholar] [CrossRef]
- Gillett, N.M.D.; Steinman, A.D. An analysis of long-term phytoplankton dynamics in Muskegon Lake—A Great Lakes Area of Concern. J. Great Lakes Res. 2011, 37, 335–342. [Google Scholar] [CrossRef]
- Bocaniov, S.A.; Lamb, K.G.; Liu, W.; Rao, Y.R.; Smith, R.E.H. High sensitivity of lake hypoxia to air temperatures, winds, and nutrient loading: Insights from a 3-D lake model. Water Resour. Res. 2020, 56, e2019WR027040. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Bernhardt, E.S. Biogeochemistry: An Analysis of Global Change, 3rd ed.; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Dugener, N.M.; Stone, I.P.; Weinke, A.D.; Biddanda, B.A. Out of oxygen: Stratification and loading drove hypoxia during a warm, wet, and productive year in a Great Lakes estuary. J. Great Lakes Res. 2023. submitted. [Google Scholar]
- Jabbari, A.; Ackerman, J.D.; Boegman, L.; Zhao, Y. Episodic hypoxia in the western basin of Lake Erie. Limnol. Oceanogr. 2019, 64, 2220–2236. [Google Scholar] [CrossRef]
- Hamidi, S.A.; Bravo, H.R.; Klump, J.V.; Waples, J.T. The role of circulation and heat fluxes in the formation of stratification leading to hypoxia in Green Bay, Lake Michigan. J. Great Lakes Res. 2015, 41, 1024–1036. [Google Scholar] [CrossRef]
- Grunert, B.K.; Brunner, S.L.; Hamadi, S.A.; Bravo, H.R.; Klump, J.V. Quantifying the influence of cold water intrusions in a shallow, coastal system across contrasting years: Green Bay, Lake Michigan. J. Great Lakes Res. 2018, 44, 851–863. [Google Scholar] [CrossRef]
- Kraus, R.T.; Knight, C.T.; Farmer, T.M.; Gorman, A.M.; Collingsworth, P.D.; Warren, G.J.; Kocovsky, P.M.; Conroy, J.D. Dynamic hypoxic zones in Lake Erie compress fish habitat, altering vulnerability to fishing gears. Can. J. Fish. Aquat. Sci. 2015, 72, 797–806. [Google Scholar] [CrossRef]
- Chamberlin, D.W.; Knight, C.T.; Kraus, R.T.; Gorman, A.M.; Xu, W.; Collingsworth, P.D. Hypoxia augments edge effects of water column stratification on fish distribution. Fisheries Res. 2020, 231, 105684. [Google Scholar] [CrossRef]
- Bertram, P.E. Total phosphorous and dissolved oxygen trends in the Central Basin of Lake Erie, 1970–1991. J. Great Lakes. Res. 1993, 19, 224–236. [Google Scholar] [CrossRef]
- Schindler, D.W.; Carpenter, S.R.; Chapra, S.C.; Hecky, R.E.; Orihel, D.M. Reducing phosphorus to curb lake eutrophication is a success. Environ. Sci Technol. 2016, 50, 8923–8929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donner, S.D.; Scavia, D. How climate controls the flux of nitrogen by the Mississippi River and the development of hypoxia in the Gulf of Mexico. Limno. Oceanogr. 2007, 52, 856–861. [Google Scholar] [CrossRef] [Green Version]
- Biddanda, B.A. Global significance of the changing freshwater carbon cycle. Eos Trans. AGU 2017, 98, 15–17. [Google Scholar] [CrossRef]
- Fergen, J.T.; Bergstrom, R.D.; Twiss, M.R.; Johnson, L.; Steinman, A.; Gagnon, V. Updated census in the Laurentian Great Lakes Watershed: A framework for determining the relationship between the population and this aquatic resource. J. Great Lakes Res. 2022, 48, 1337–1344. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Duration of Stratification (Days) | Mild Hypoxia 8 m | Severe Hypoxia 8 m | Near Anoxia 8 m | Mild Hypoxia 11 m | Severe Hypoxia 11 m | Near Anoxia 11 m | Hypoxic Severity Index |
|---|---|---|---|---|---|---|---|---|
| 2015 | 70 | 12 | 0 | 0 | 23 | 15 | 2 | 20.95 |
| 2019 | 79 | 25 | 1 | 0 | 45 | 16 | 2 | 25.86 |
| 2014 | 79 | 18 | 0 | 0 | 41 | 13 | 0 | 26.13 |
| 2013 | 77 | 33 | 21 | 0 | 52 | 40 | 0 | 42.32 |
| 2017 | 54 | 8 | 7 | 0 | 36 | 36 | 1 | 44.96 |
| 2018 | 122 | 16 | 22 | 0 | 8 | 26 | 9 | 51.52 |
| 2016 | 67 | 22 | 14 | 2 | 27 | 42 | 6 | 56.96 |
| 2011 | 114 | 17 | 40 | 8 | 35 | 34 | 16 | 63.38 |
| 2012 | 61 | 31 | 7 | 0 | 6 | 39 | 19 | 91.27 |
| 2021 | 87 | 24 | 14 | 0 | 13 | 32 | 28 | 97.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dugener, N.M.; Weinke, A.D.; Stone, I.P.; Biddanda, B.A. Recurringly Hypoxic: Bottom Water Oxygen Depletion Is Linked to Temperature and Precipitation in a Great Lakes Estuary. Hydrobiology 2023, 2, 410-430. https://doi.org/10.3390/hydrobiology2020027
Dugener NM, Weinke AD, Stone IP, Biddanda BA. Recurringly Hypoxic: Bottom Water Oxygen Depletion Is Linked to Temperature and Precipitation in a Great Lakes Estuary. Hydrobiology. 2023; 2(2):410-430. https://doi.org/10.3390/hydrobiology2020027
Chicago/Turabian StyleDugener, Nathan M., Anthony D. Weinke, Ian P. Stone, and Bopaiah A. Biddanda. 2023. "Recurringly Hypoxic: Bottom Water Oxygen Depletion Is Linked to Temperature and Precipitation in a Great Lakes Estuary" Hydrobiology 2, no. 2: 410-430. https://doi.org/10.3390/hydrobiology2020027