
Morphological and Molecular Characterization of the Benthic Dinoflagellate Amphidinium from Coastal Waters of Mexico

 ,
,  ,
,  ,
,  ,
,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling, Cell Isolation, and Culture of Amphidinium
2.2. Cell Morphology and Statistical Analysis
2.3. Total Genomic DNA Extraction and Amplification
2.4. Sequencing and Phylogenetic Analysis
3. Results
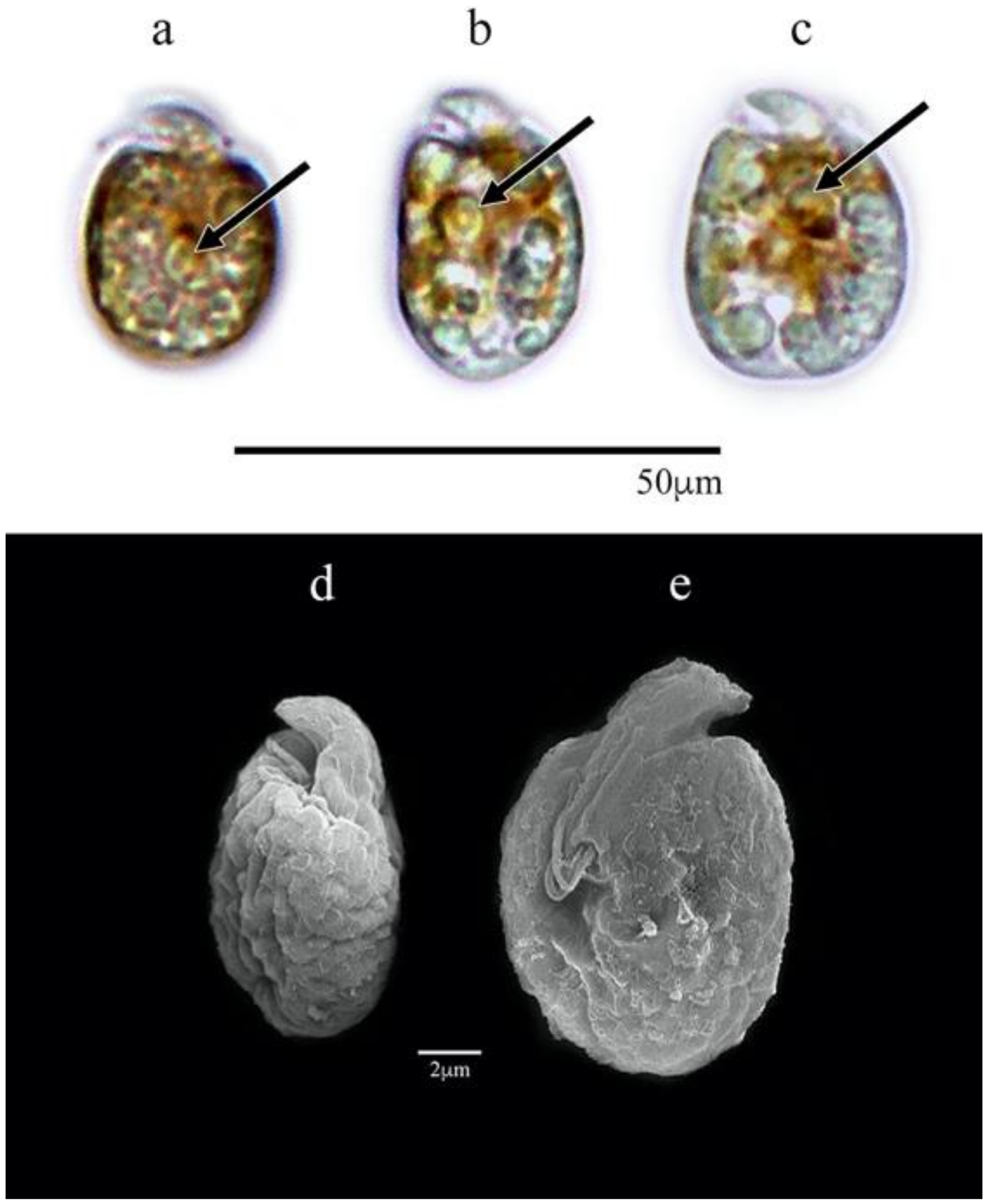
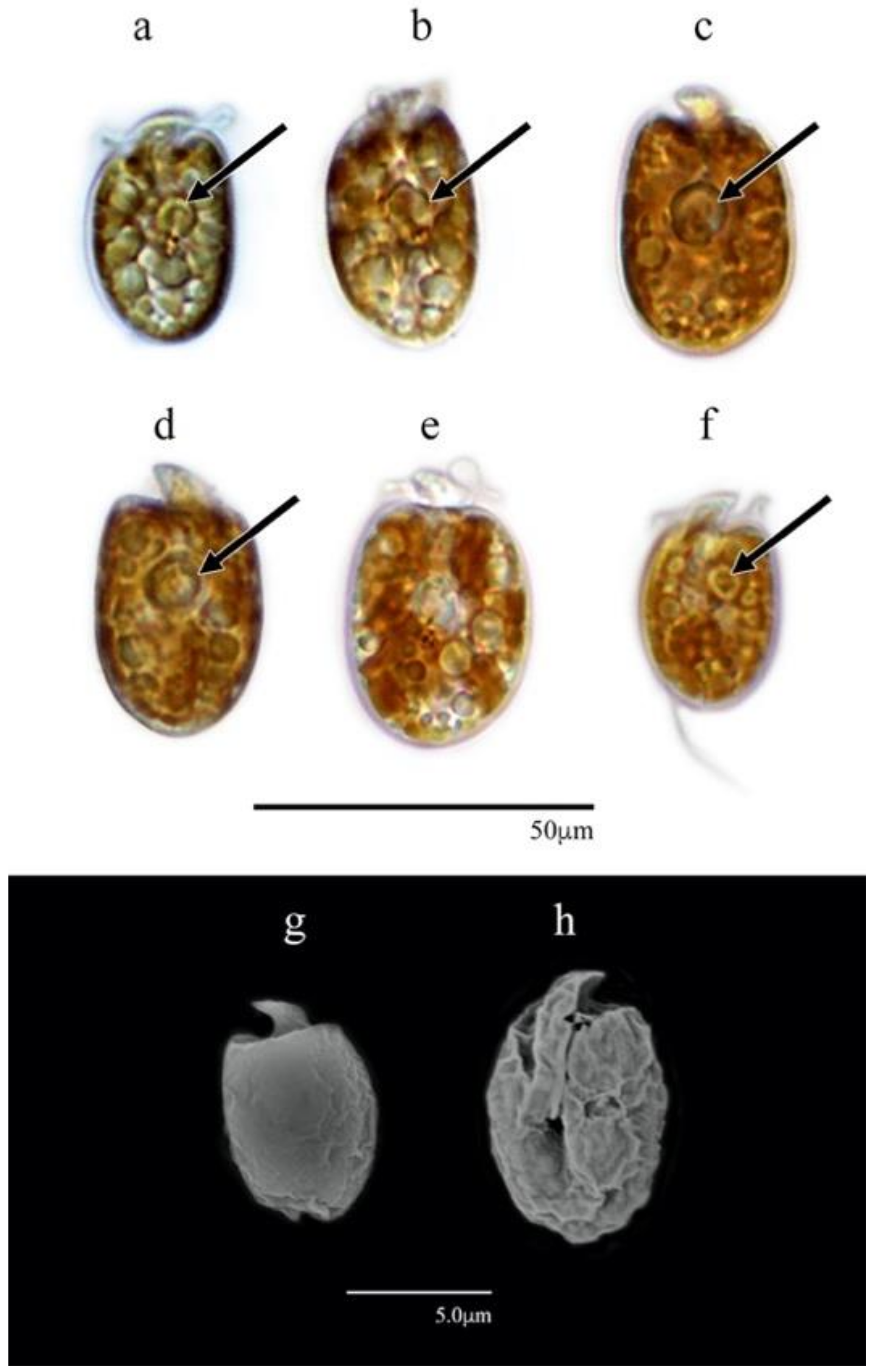
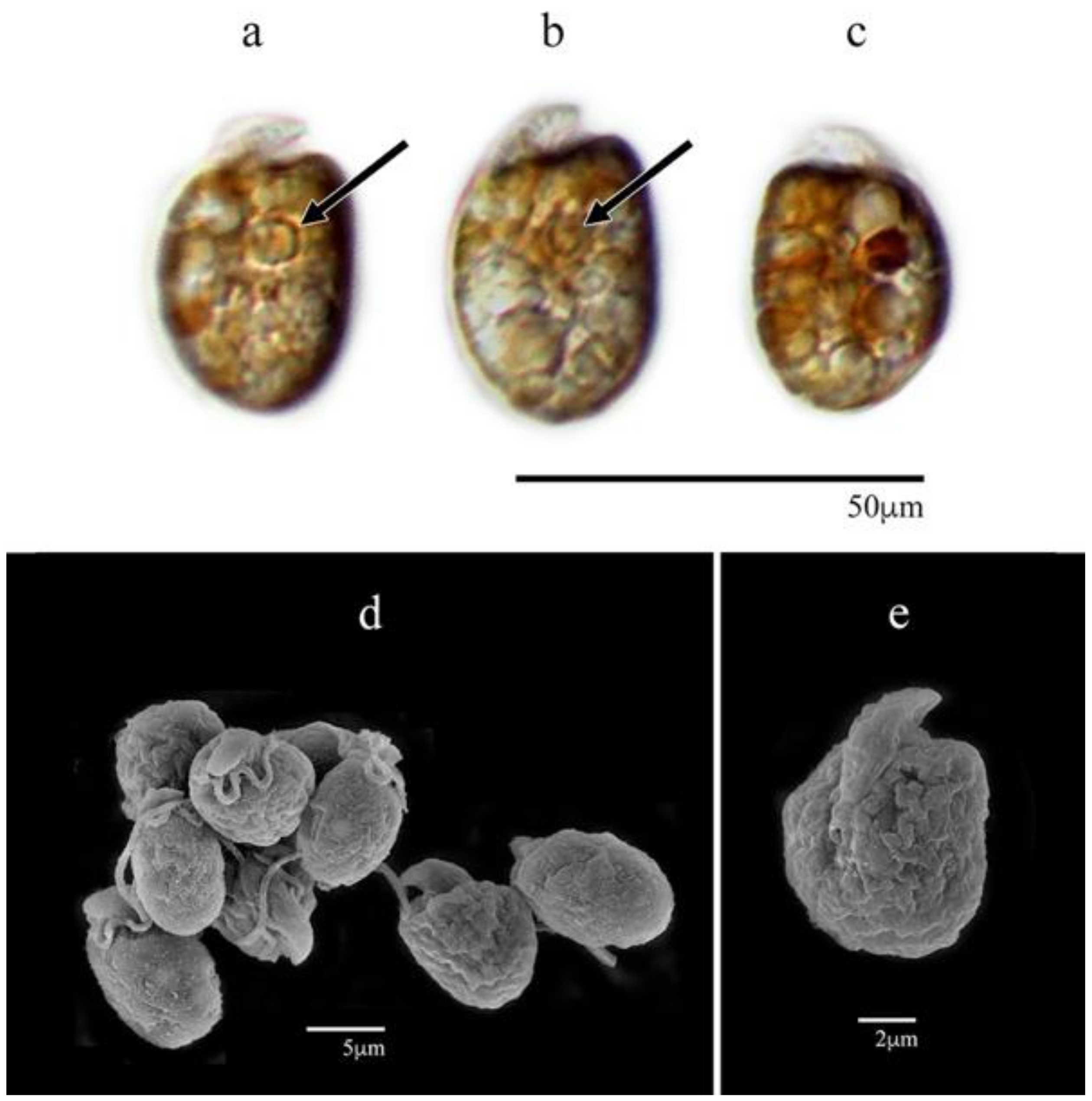
3.1. Morphological Characterization
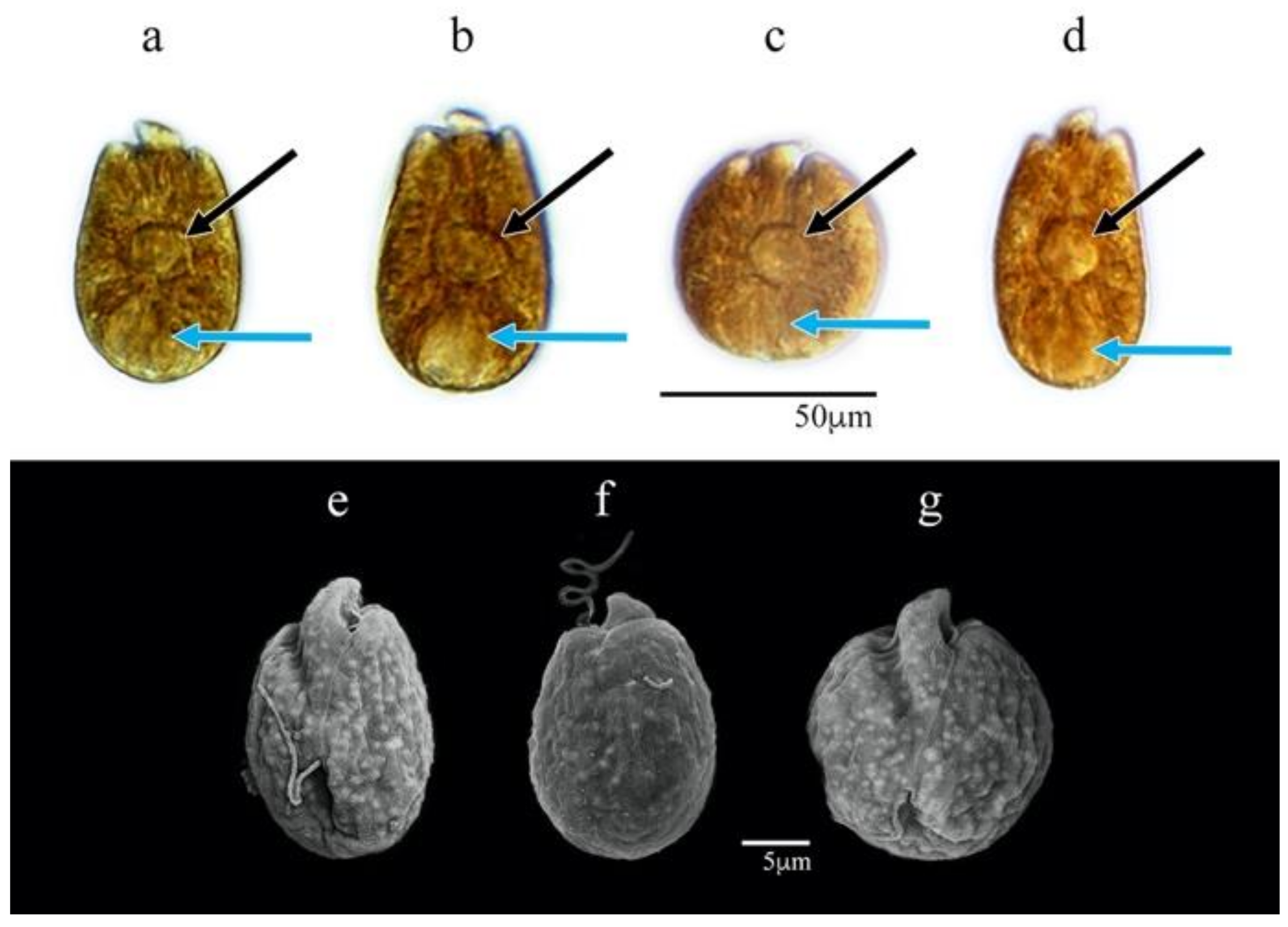
3.1.1. Amphidinium carterae (AcSAV105)
3.1.2. Amphidinium cf. carterae (AeSQ172, AeSQ177, and AeSQ181)
3.1.3. Amphidinium massartii (AmLT112)
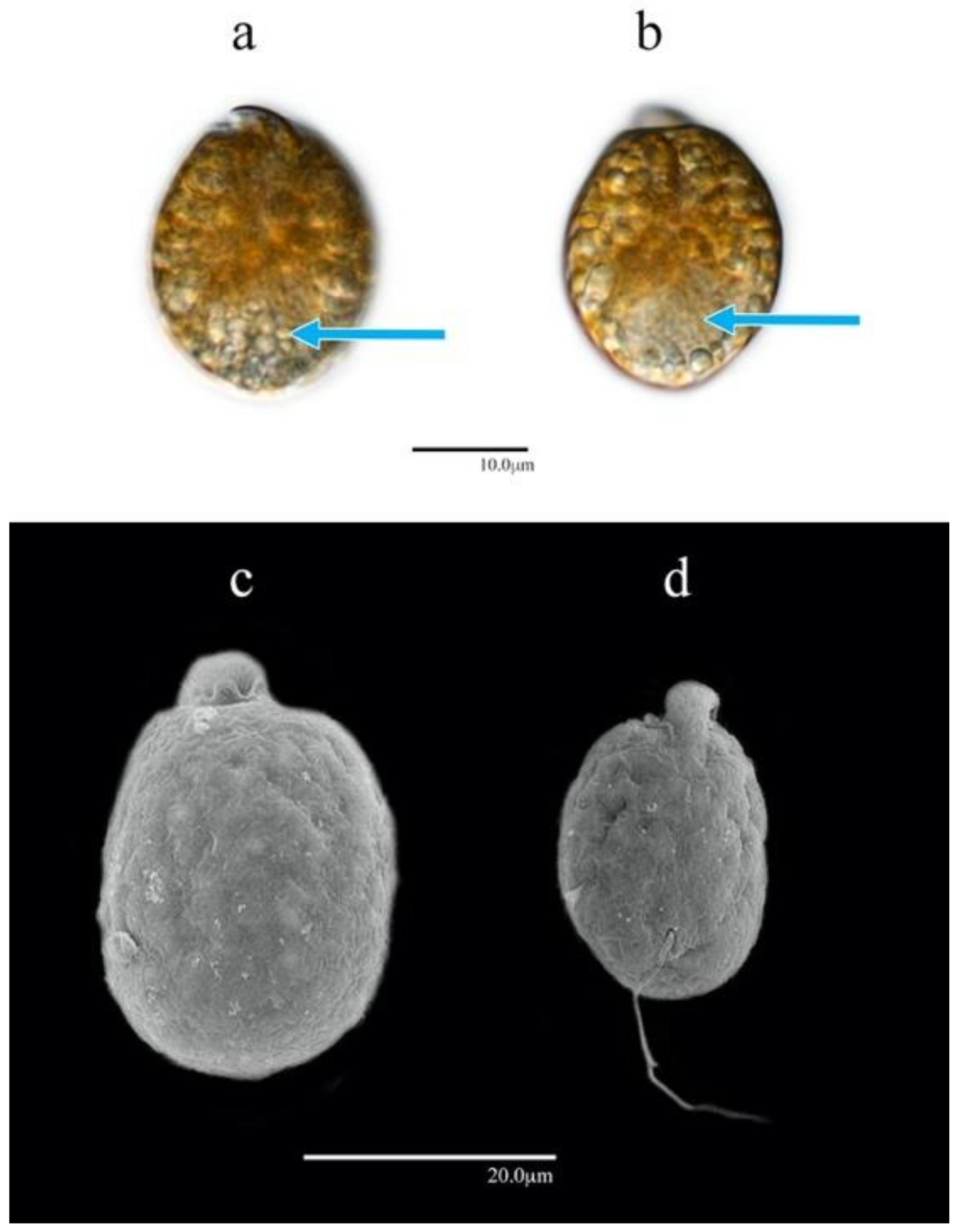
3.1.4. Amphidinium operculatum (AA60)
3.1.5. Amphidinium theodorei (AtLPZ38)
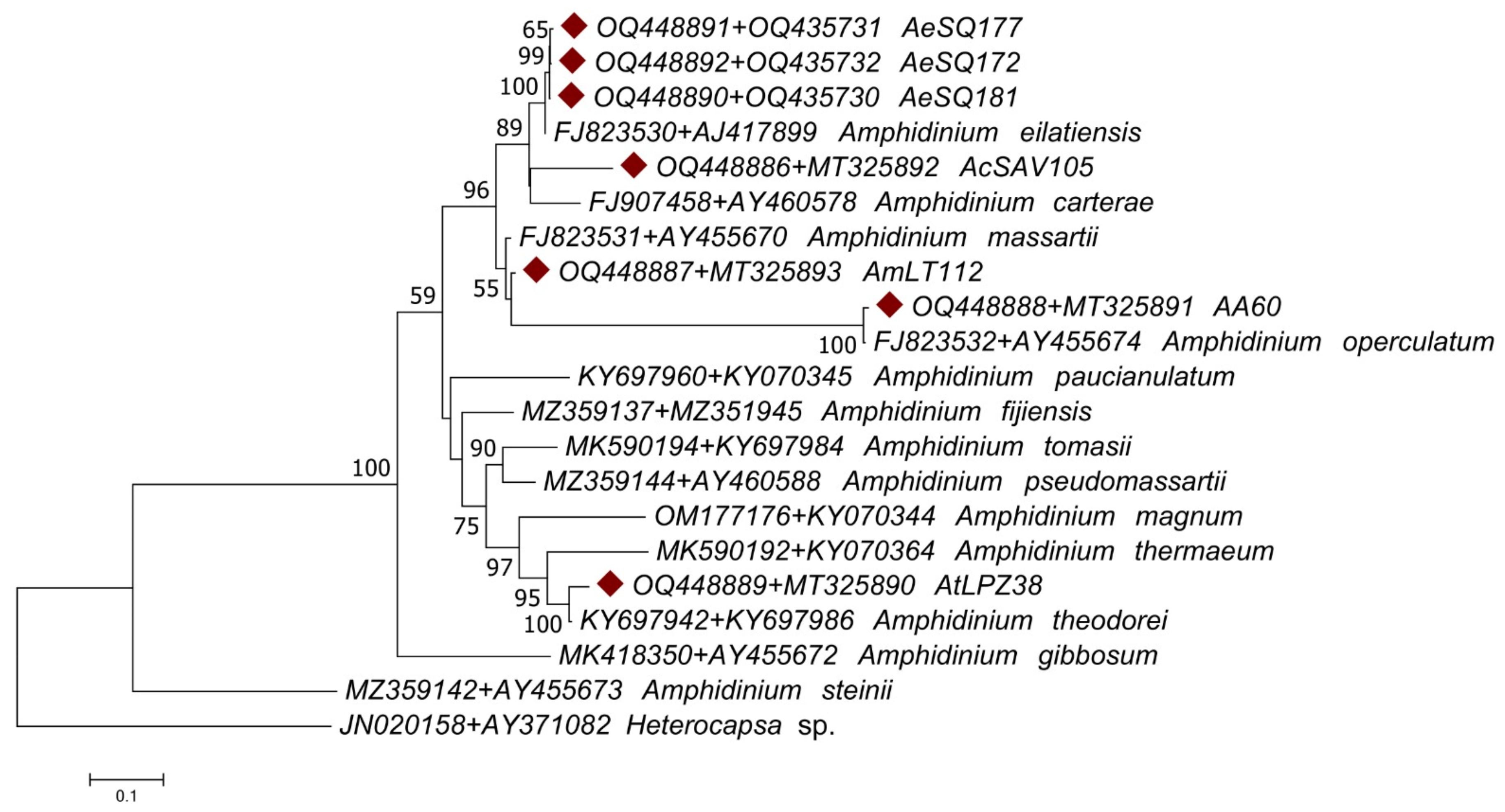
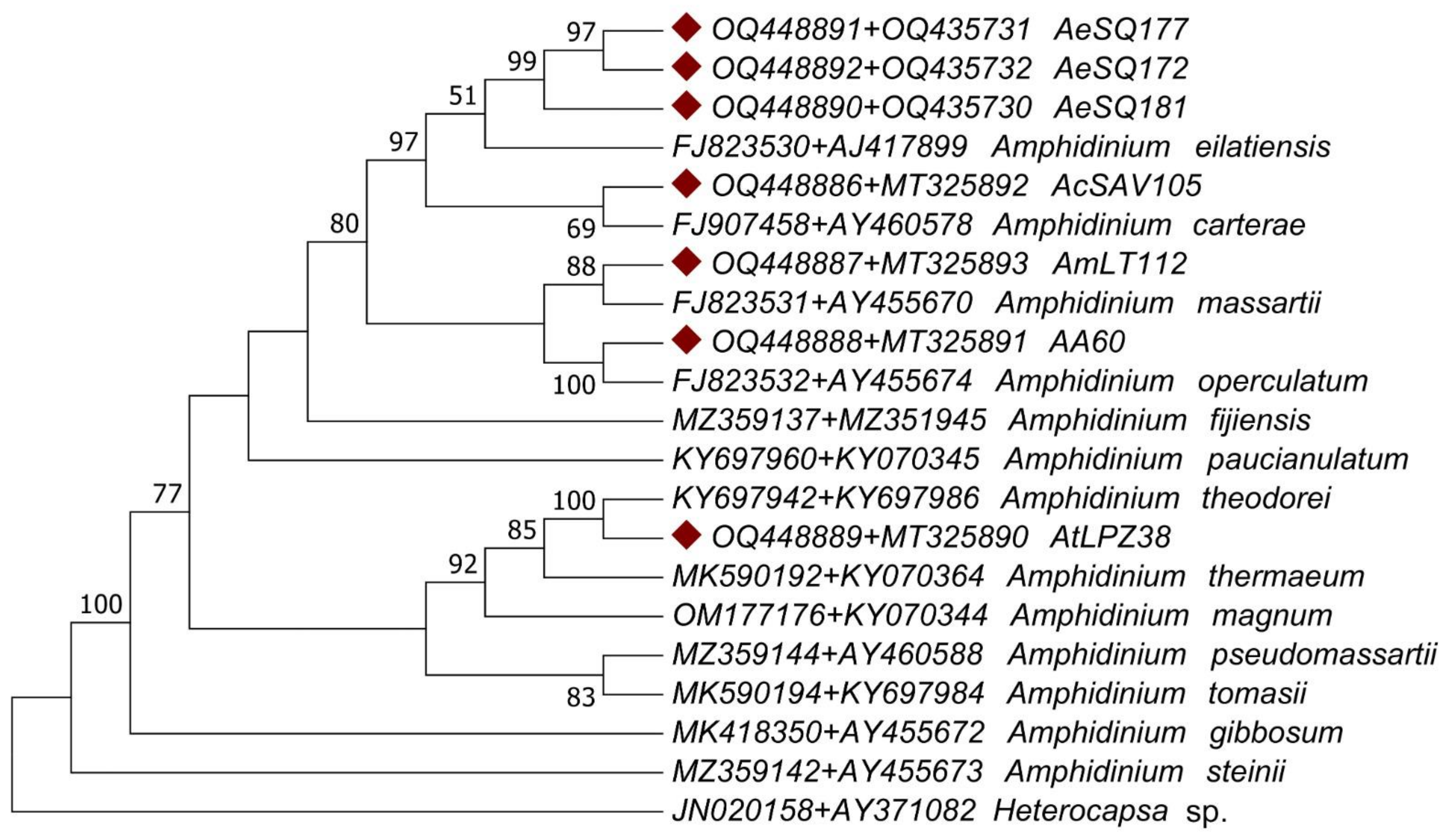
3.2. Molecular Characterization
4. Discussion
4.1. Morphological Characterization
4.2. Molecular Phylogenetic Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karafas, S.; Teng, S.T.; Leaw, C.P.; Alves-de-Souza, C. An evaluation of the genus Amphidinium (Dinophyceae) combining evidence from morphology, phylogenetics, and toxin production, with the introduction of six novel species. Harmful Algae 2017, 68, 128–151. [Google Scholar] [CrossRef] [PubMed]
- Flø Jørgensen, M.; Murray, S.; Daugbjerg, N. Amphidinium revisited. I. Redefinition of Amphidinium (Dinophyceae) based on cladistic and molecular phylogenetic analyses. J. Phycol. 2004, 40, 351–365. [Google Scholar] [CrossRef]
- Hoppenrath, M.; Murray, S.A.; Chomérat, N.; Horiguchi, T. Marine Benthic Dinoflagellates-Unveiling Their Worldwide Biodiversity; Schweizerbart’sche Verlagsbuchhandlung: Stuttgart, Germany, 2014; Volume 54, p. 276. [Google Scholar]
- Murray, S.; Flø Jørgensen, M.; Daugbjerg, N.; Rhodes, L. Amphidinium revisited. II. Resolving species boundaries in the Amphidinium operculatum species complex (Dinophyceae), including the descriptions of Amphidinium trulla sp. nov. and Amphidinium gibbosum. comb. nov. J. Phycol. 2004, 40, 366–382. [Google Scholar] [CrossRef]
- Kofoid, C.A.; Swezy, O. The Free-Living Unarmored Dinoflagellata; University of California Press: Berkeley, CA, USA, 1921; Volume 5, p. 563. [Google Scholar]
- Steidinger, K.A.; Tangen, K. Dinoflagellates. In Identifying Marine Phytoplankton; Tomas, C., Ed.; Academic Press: San Diego, CA, USA, 1997; pp. 387–584. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. 2023. Available online: https://www.algaebase.org, (accessed on 5 January 2022).
- Durán-Riveroll, L.D.; Cembella, A.D.; Okolodkov, Y.B. A review on the biodiversity and biogeography of toxigenic benthic marine dinoflagellates of the coasts of Latin America. Front. Mar. Sci. 2019, 6, 148. [Google Scholar] [CrossRef]
- Tester, P.A.; Litaker, R.W.; Berdalet, E. Climate change and harmful benthic microalgae. Harmful Algae 2020, 91, e101655. [Google Scholar] [CrossRef]
- Okolodkov, Y.B.; Campos-Bautista, G.; Gárate-Lizárraga, I.; González-González, J.A.G.; Hoppenrath, M.; Arenas, V. Seasonal changes of benthic and epiphytic dinoflagellates in the Veracruz reef zone, Gulf of Mexico. Aquat. Microb. Ecol. 2007, 47, 223–237. [Google Scholar] [CrossRef]
- Gárate-Lizárraga, I. Proliferation of Amphidinium carterae (Gymnodiniales: Gymnodiniaceae) in Bahía de La Paz, Gulf of California. CICIMAR Oceánides 2012, 27, 37–49. [Google Scholar] [CrossRef]
- Gárate-Lizárraga, I.; González-Armas, R.; Verdugo-Díaz, G.; Okolodkov, Y.B.; Pérez-Cruz, B.; Díaz-Ortíz, J.A. Seasonality of the dinoflagellate Amphidinium cf. carterae (Dinophyceae: Amphidiniales) in Bahía de la Paz, Gulf of California. Mar. Poll. Bull. 2019, 146, 532–541. [Google Scholar] [CrossRef]
- Murray, S.A.; Kohli, G.S.; Farrell, H.; Spiers, Z.B.; Place, A.R.; Dorantes-Aranda, J.J.; Ruszczyk, J. A fish kill associated with a bloom of Amphidinium carterae in a coastal lagoon in Sydney, Australia. Harmful Algae 2015, 49, 19–28. [Google Scholar] [CrossRef]
- Wellkamp, M.; García-Camacho, F.; Durán-Riveroll, L.M.; Tebben, J.; Tillmann, U.; Krock, B. LC-MS/MS method development for the discovery and identification of amphidinols produced by Amphidinium. Mar. Drugs 2020, 18, 1–22. [Google Scholar] [CrossRef]
- Echigoya, R.; Rhodes, L.; Oshima, Y.; Satake, M. The structures of five new antifungal and hemolytic amphidinol analogs from Amphidinium carterae collected in New Zealand. Harmful Algae 2005, 4, 383–389. [Google Scholar] [CrossRef]
- Morsy, N.; Houdai, T.; Matsuoka, S.; Matsumori, N.; Adachi, S.; Oishi, T.; Murata, M.; Iwashita, T.; Fujita, T. Structures of new amphidinols with truncated polyhydroxyl chain and their membrane-permeabilizing activities. Bioorg. Med. Chem. 2006, 14, 6548–6554. [Google Scholar] [CrossRef]
- Satake, M.; Cornelio, K.; Hanashima, S.; Malabed, R.; Murata, M.; Matsumori, N.; Zhang, H.; Hayashi, F.; Mori, S.; Kim, J.S. Structures of the largest amphidinol homologues from the dinoflagellate Amphidinium carterae and structure–activity relationships. J. Nat. Prod. 2017, 80, 2883–2888. [Google Scholar] [CrossRef]
- Mejía-Camacho, A.L.; Durán-Riveroll, L.M.; Cembella, A.D. Toxicity bioassay and cytotoxic effects of the benthic marine dinoflagellate Amphidinium operculatum. J. Xenobiot. 2021, 11, 33–45. [Google Scholar] [CrossRef]
- Martínez, K.A.; Lauritano, C.; Druka, D.; Romano, G.; Grohmann, T.; Jaspars, M.; Martín, J.; Díaz, C.; Cautain, B.; de la Cruz, M.; et al. Amphidinol 22, a new cytotoxic and antifungal amphidinol from the dinoflagellate Amphidinium carterae. Mar. Drugs 2019, 17, 385. [Google Scholar] [CrossRef]
- Rhodes, L.L.; Smith, K.F.; Munday, R.; Selwood, A.I.; McNabb, P.S.; Holland, P.T.; Bottein, M.-Y. Toxic dinoflagellates (Dinophyceae) from Rarotonga, Cook Islands. Toxicon 2010, 56, 751–758. [Google Scholar] [CrossRef]
- Dolapsakis, N.P.; Economou-Amilli, A. A new marine species of Amphidinium (Dinophyceae) from Thermaikos Gulf, Greece. Acta Protozool. 2009, 48, 153–170. [Google Scholar]
- Murray, S.; Patterson, D. The benthic dinoflagellate genus Amphidinium in south-eastern Australian waters, including three new species. Eur. J. Phycol. 2002, 37, 279–298. [Google Scholar] [CrossRef]
- Okolodkov, Y.; Durán-Riveroll, L.D.; Band-Schmidt, C.J.; Leyva-Valencia, I.; Gárate-Lizárraga, I.; Cembella, A.D. A review on marine benthic dinoflagellates in Mexico. Hidrobiológica 2022, 32, 183–210. [Google Scholar] [CrossRef]
- Okolodkov, Y.B.; Gárate-Lizárraga, I. An annotated checklist of dinoflagellates (Dinophyceae) from the Mexican Pacific. Acta Bot. Mex. 2006, 74, 1–154. [Google Scholar] [CrossRef]
- Blackburn, S.I.; Bolch, C.J.S.; Haskard, K.A.; Hallegraeff, G.M. Reproductive compatibility among four global populations of the toxic dinoflagellate Gymnodinium catenatum (Dinophyceae). Phycologia 2001, 40, 78–87. [Google Scholar] [CrossRef]
- Markham, J.; Hagmeier, E. Observations on the effects of germanium dioxide on the growth of macro-algae and diatoms. Phycologia 1982, 21, 125–130. [Google Scholar] [CrossRef]
- Pérez-López, H.; Durán-Riveroll, L.; Gómez-Lizárraga, L.E.; Mendoza-Garfias, M.B. Simple method for preparing delicate dinoflagellate of the genus Amphidinium for scanning electron microscopy. Microsc. Microanal. 2020, 26, 1366–1369. [Google Scholar] [CrossRef]
- RStudio-Team. RStudio: Integrated Development for R; RStudio: Boston, MA, USA, 2020. [Google Scholar]
- Gómez, F. A quantitative review of the lifestyle, habitat and trophic diversity of dinoflagellates (Dinoflagellata, Alveolata). Syst. Biodiver. 2012, 10, 267–275. [Google Scholar] [CrossRef]
- Taylor, F.; Hoppenrath, M.; Saldarriaga, J.F. Dinoflagellate diversity and distribution. Biodivers. Conserv. 2008, 17, 407–418. [Google Scholar] [CrossRef]
- Scholin, C.; Herzog, M.; Sogin, M.; Anderson, D. Identification of group- and strain-specific genetic markers for globally distributed Alexandrium (Dinophyceae). II. sequence analysis of a fragment of the LSU rRNA gene1. J. Phycol. 1994, 30, 999–1011. [Google Scholar] [CrossRef]
- Daugbjerg, N.; Hansen, G.; Larsen, J.; Moestrup, Ø. Phylogeny of some of the major genera of dinoflagellates based on ultrastructure and partial LSU rDNA sequence data, including the erection of three new genera of unarmoured dinoflagellates. Phycologia 2000, 39, 302–317. [Google Scholar] [CrossRef]
- Litaker, R.W.; Vandersea, M.W.; Kibler, S.R.; Reece, K.S.; Stokes, N.A.; Steidinger, K.A.; Millie, D.F.; Bendis, B.J.; Pigg, R.J.; Tester, P.A. Identification of Pfiesteria piscicida (Dinophyceae) and Pfiesteria-like organisms using internal transcribed spacer-specific PCR assays 1. J. Phycol. 2003, 39, 754–761. [Google Scholar] [CrossRef]
- Moreira-González, A.R.; Fernandes, L.F.; Uchida, H.; Uesugi, A.; Suzuki, T.; Chomérat, N.; Bilien, G.; Pereira, T.A.; Mafra Jr, L.L. Morphology, growth, toxin production, and toxicity of cultured marine benthic dinoflagellates from Brazil and Cuba. J. Appl. Phycol. 2019, 31, 3699–3719. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 1–19. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Lee, K.H.; Jeong, H.J.; Park, K.; Kang, N.S.; Yoo, Y.D.; Lee, M.J.; Lee, J.; Lee, S.; Kim, T.; Kim, H.S.; et al. Morphology and molecular characterization of the epiphytic dinoflagellate Amphidinium massartii, isolated from the temperate waters off Jeju Island, Korea. Algae 2013, 28, 213–231. [Google Scholar] [CrossRef]
- Lee, J.J.; Olea, R.; Cevasco, M.; Pochon, X.; Correia, M.; Shpigel, M.; Pawlowski, J. A marine dinoflagellate, Amphidinium eilatiensis n. sp., from the benthos of a mariculture sedimentation pond in Eilat, Israel. J. Eukar. Microbiol. 2003, 50, 439–448. [Google Scholar] [CrossRef]
- Hulburt, E.M. The taxonomy of unarmored Dinophyceae of shallow embayments on Cape Cod, Massachusetts. Biol. Bull. 1957, 112, 196–219. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Murray, S.; Garby, T.; Hoppenrath, M.; Neilan, B. Genetic diversity, morphological uniformity and polyketide production in dinoflagellates (Amphidinium, Dinoflagellata). PLoS ONE 2012, 7, e38253. [Google Scholar] [CrossRef]
- Tamura, M.; Takano, Y.; Horiguchi, T. Discovery of a novel type of body scale in the marine dinoflagellate, Amphidinium cupulatisquama sp. nov. (Dinophyceae). Phycol. Res. 2009, 57, 304–312. [Google Scholar] [CrossRef]
- Sekida, S.; Okuda, K.; Katsumata, K.; Horiguchi, T. A novel type of body scale found in two strains of Amphidinium species (Dinophyceae). Phycologia 2003, 42, 661–666. [Google Scholar] [CrossRef]
- Luo, Z.; Wang, N.; Mohamed, H.F.; Liang, Y.; Pei, L.; Huang, S.; Gu, H. Amphidinium stirisquamtum sp. nov. (Dinophyceae), a new marine sand-dwelling dinoflagellate with a novel type of body scale. Algae 2021, 36, 241–261. [Google Scholar] [CrossRef]
- Fraga, S.; Rodríguez, F.; Bravo, I.; Zapata, M.; Marañón, E. Review of the main ecological features affecting benthic dinoflagellate blooms. Cryptogam. Algol. 2012, 33, 171–179. [Google Scholar] [CrossRef]
- Ricklefs, R.E. Estimating diversification rates from phylogenetic information. Trends Ecol. Evol. 2007, 22, 601–610. [Google Scholar] [CrossRef]
- Kolaczkowski, B.; Thornton, J.W. Performance of maximum parsimony and likelihood phylogenetics when evolution is heterogeneous. Nature 2004, 431, 980–984. [Google Scholar] [CrossRef]
- Gribble, K.E.; Anderson, D.M. Molecular phylogeny of the heterotrophic dinoflagellates Protoperidinium, Diplopsalis and Preperidinium (Dinophyceae), inferred from Large Subunit rDNA. J. Phycol. 2006, 42, 1081–1095. [Google Scholar] [CrossRef]
- Stanley, S.M. Earth System History, 4th ed.; Macmillan: New York, NY, USA, 2005. [Google Scholar]
- Avise, J.C. Molecular Markers, Natural History and Evolution; Springer Science & Business Media: New York, NY, USA, 2012; pp. 120–128. [Google Scholar]
- Durán-Riveroll, L.M.; Weber, J.; Krock, B. First identification of amphidinols from Mexican strains and new analogs. Toxins 2023, 15, 163. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
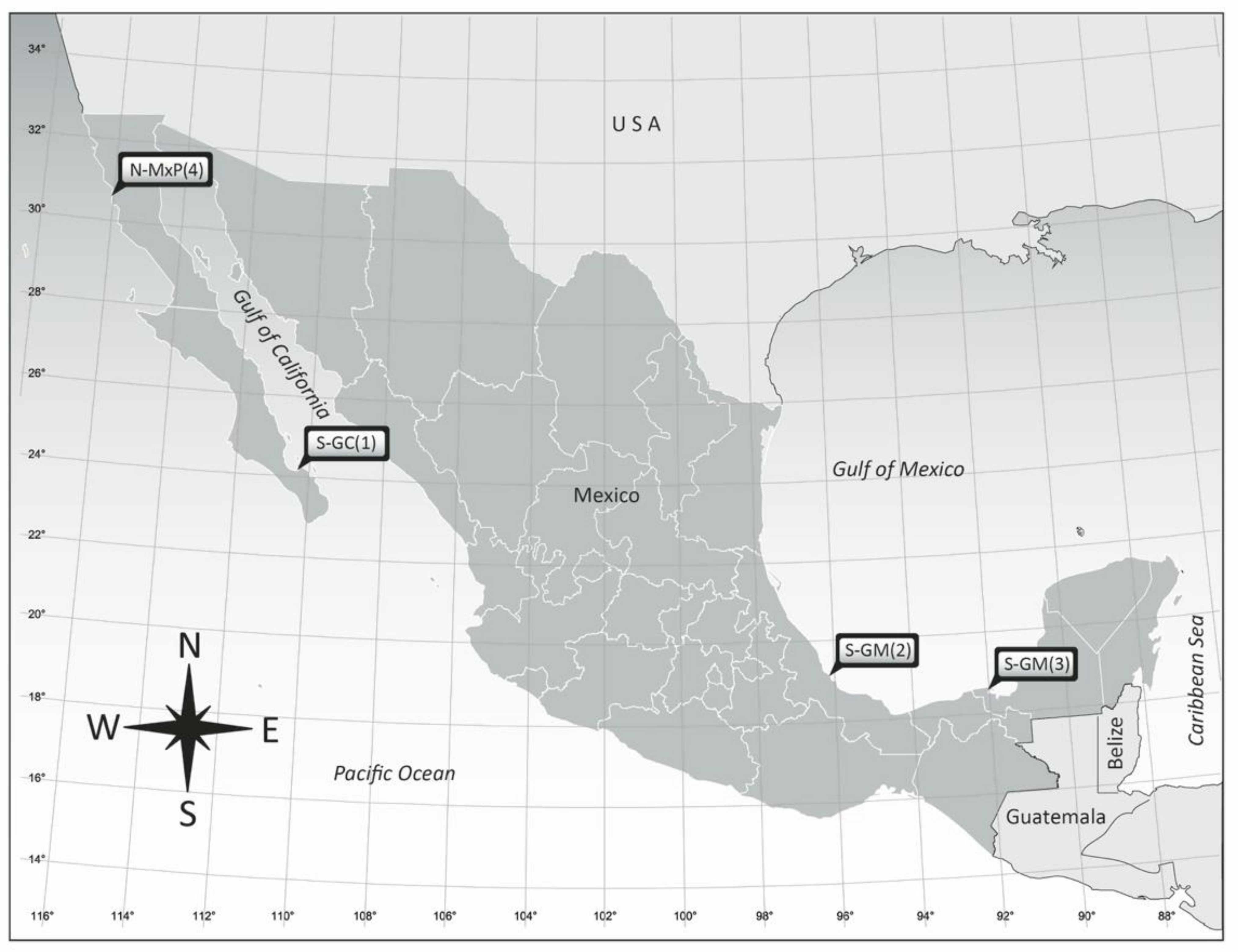
| Isolate | Locality of Origin | Geographical Region | Substrate Type | Provisional Morphospecies |
|---|---|---|---|---|
| AcSAV105 | Veracruz Reef System, Veracruz | S-GM (2) | Anchored buoy | A. carterae Hulburt |
| AeSQ172 | San Quintín, BC | N-MxP (4) | Zostera marina | A. cf. carterae |
| AeSQ177 | San Quintín, BC | N-MxP (4) | Z. marina | A. cf. carterae |
| AeSQ181 | San Quintín, BC | N-MxP (4) | Z. marina | A. cf. carterae |
| AmLT112 | Laguna de Términos, Campeche | S-GM (3) | Thalassia testudinum | A. massartii Biecheler |
| AA60 | Veracruz Reef System, Veracruz | S-GM (2) | Padina sp. | A. operculatum Clap. & J. Lachm. |
| AtLPZ38 | Balandra, La Paz, BCS | S-GC (1) | Floating brown alga | A. theodorei C.R. Tomas & Karafas |
| ITS2 | LSU | |||||||
|---|---|---|---|---|---|---|---|---|
| Taxa | Strain | GenBank Acc. | bp | Authors * | Strain | GenBank Acc. | bp | Authors * |
| A. carterae | DL1100 | FJ907458 | 636 | Zhao et al. | CS-740 | AY460578 | 1359 | Murray et al. |
| A. eilatiensis | CCMP2100 | AJ417900 | 576 | Lee et al. | CCMP2100 | AJ417900 | 886 | Lee et al. |
| A. fijiensis | Amfi0508-1 | KY697937 | 1053 | Karafas et al. | WZD19 | MZ351945 | 1304 | Luo et al. |
| A. gibbosum | D2A1 | MK418355 | 533 | Zhang et al. | CCMP 120 | AY455672 | 1325 | Jörgensen et al. |
| A. magnum | ZS606 | OM177176 | 506 | Xie, H. | Amma0206-1 | KY070341 | 1163 | Karafas et al. |
| A. massartii | NEPCC 802 | FJ823531 | 522 | Stern et al. | CCMP 1821 | AY455670 | 1338 | Jörgensen et al. |
| A. operculatum | SKLMP_S091 | MK605120 | 1499 | Yiu et al. | SM06 | AY455674 | 1376 | Jörgensen et al. |
| A. paucianulatum | Ampa0606-2 | KY697960 | 1056 | Karafas et al. | Ampa0606-1 | KY070345 | 1163 | Karafas et al. |
| A. pseudomassartii | AKLV01 | KY697945 | 1010 | Karafas et al. | AKLSPO1 | AY460588 | 1357 | Murray et al. |
| A. steinii | TIO181 | MZ359142 | 602 | Luo et al. | SM12 | AY460593 | 1426 | Murray et al. |
| A. theodorei | Amth0702-1 | KY697942 | 695 | Karafas et al. | Amth1303-1 | KY697986 | 1317 | Karafas et al. |
| A. thermaeum | Amth00600FA1 | KY697956 | 803 | Karafas et al. | Amth0304-1 | KY070365 | 1159 | Karafas et al. |
| A. tomasii | Amto1412-1 | KY974334 | 1127 | Karafas et al. | Amto1412-2 | KY697984 | 1270 | Karafas et al. |
| Heterocapsa sp. | NA | JN020158 | 640 | Zadabbas et al. | CCMP424 | AY371082 | 860 | De Salas, M. |
| A. carterae | AcSAV105 | OQ448886 | 559 | Present study | AcSAV105 | MT325892 | 949 | Present study |
| A. eilatiensis | AeSQ172 | OQ448892 | 596 | Present study | AeSQ172 | OQ435732 | 860 | Present study |
| A. eilatiensis | AeSQ177 | OQ448891 | 593 | Present study | AeSQ177 | OQ435731 | 857 | Present study |
| A. eilatiensis | AeSQ181 | OQ448890 | 590 | Present study | AeSQ181 | OQ435730 | 857 | Present study |
| A. massartii | AmLT112 | OQ448887 | 715 | Present study | AmLT112 | MT325893 | 926 | Present study |
| A. operculatum | AA60 | OQ448888 | 719 | Present study | AA60 | MT325891 | 874 | Present study |
| A. theodorei | AtLPZ38 | OQ448889 | 732 | Present study | AtLPZ38 | MT325890 | 813 | Present study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Durán-Riveroll, L.M.; Juárez, O.E.; Okolodkov, Y.B.; Mejía-Camacho, A.L.; Ramírez-Corona, F.; Casanova-Gracia, D.; Osorio-Ramírez, M.d.C.; Cervantes-Urieta, V.A.; Cembella, A.D. Morphological and Molecular Characterization of the Benthic Dinoflagellate Amphidinium from Coastal Waters of Mexico. Phycology 2023, 3, 305-324. https://doi.org/10.3390/phycology3020020
Durán-Riveroll LM, Juárez OE, Okolodkov YB, Mejía-Camacho AL, Ramírez-Corona F, Casanova-Gracia D, Osorio-Ramírez MdC, Cervantes-Urieta VA, Cembella AD. Morphological and Molecular Characterization of the Benthic Dinoflagellate Amphidinium from Coastal Waters of Mexico. Phycology. 2023; 3(2):305-324. https://doi.org/10.3390/phycology3020020
Chicago/Turabian StyleDurán-Riveroll, Lorena María, Oscar E. Juárez, Yuri B. Okolodkov, Ana Luisa Mejía-Camacho, Fabiola Ramírez-Corona, Dania Casanova-Gracia, María del Carmen Osorio-Ramírez, Victor A. Cervantes-Urieta, and Allan D. Cembella. 2023. "Morphological and Molecular Characterization of the Benthic Dinoflagellate Amphidinium from Coastal Waters of Mexico" Phycology 3, no. 2: 305-324. https://doi.org/10.3390/phycology3020020