
Entomopathogenic Fungi: Interactions and Applications

 , ,
, , 
Definition
:1. Introduction
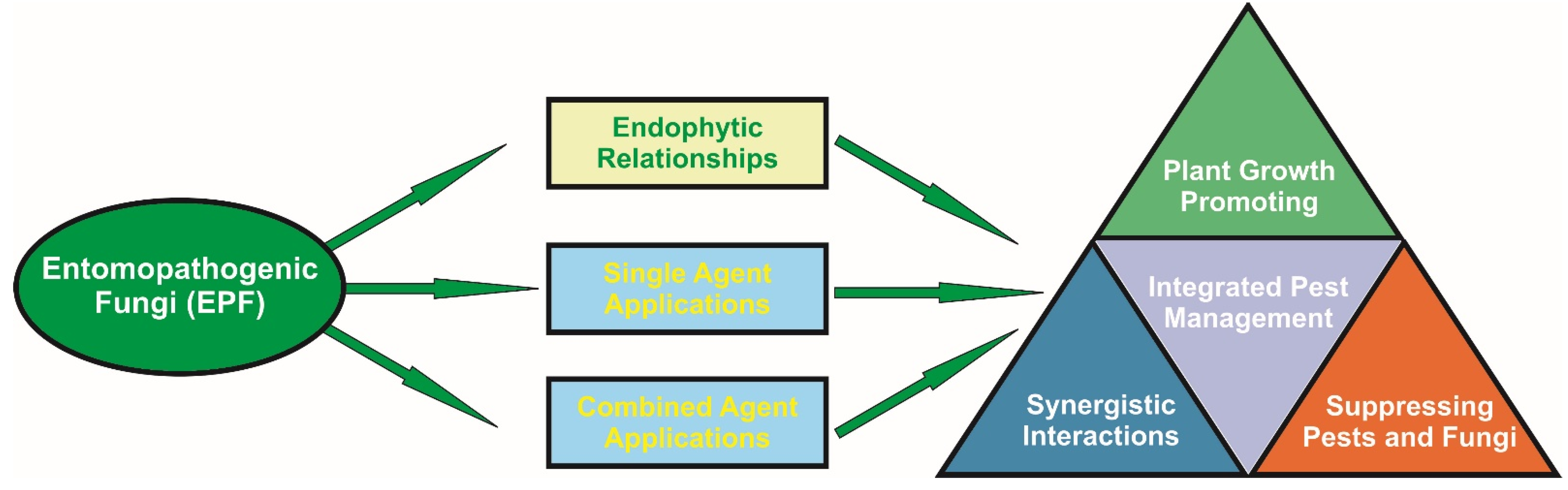
2. Interactions and Applications
2.1. Plant—EPF Interactions
2.2. Single Agent Applications—Mode of Action
2.3. Combined Agent’s Applications
3. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Entry Link on the Encyclopedia Platform
References
- Rehner, S.A.; Vega, F.E.; Blackwell, M. Phylogenetics and Insect Pathogenic Genus Beauveria. In Insect–Fungal Associations: Ecology and Evolution; Oxford University Press, Inc.: New York, NY, USA, 2005; pp. 3–25. [Google Scholar]
- Zimmermann, G.; Papierok, B.; Glare, T. Elias Metschnikoff, Elie Metchnikoff or Ilya Ilich Mechnikov (1845–1916): A Pioneer in Insect Pathology, the First Describer of the Entomopathogenic Fungus Metarhizium anisopliae and How to Translate a Russian Name. Biocontrol Sci. Technol. 1995, 5, 527–530. [Google Scholar] [CrossRef]
- Lord, J.C. From Metchnikoff to Monsanto and beyond: The path of microbial control. J. Invertebr. Pathol. 2005, 89, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Lacey, L.A.; Frutos, R.; Kaya, H.K.; Vail, P. Insect Pathogens as Biological Control Agents: Do They Have a Future? Biol. Control 2001, 21, 230–248. [Google Scholar] [CrossRef]
- Ownley, B.H.; Griffin, M.R.; Klingeman, W.; Gwinn, K.D.; Moulton, J.K.; Pereira, R. Beauveria bassiana: Endophytic colonization and plant disease control. J. Invertebr. Pathol. 2008, 98, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Goettel, M.S.; Koike, M.; Kim, J.J.; Aiuchi, D.; Shinya, R.; Brodeur, J. Potential of Lecanicillium spp. for management of insects, nematodes and plant diseases. J. Invertebr. Pathol. 2008, 98, 256–261. [Google Scholar] [CrossRef]
- Sasan, R.K.; Bidochka, M.J. Antagonism of the endophytic insect pathogenic fungus Metarhizium robertsii against the bean plant pathogen Fusarium solani f. sp. phaseoli. Can. J. Plant Pathol. 2013, 35, 288–293. [Google Scholar] [CrossRef]
- Jaber, L.R.; Salem, N.M. Endophytic colonisation of squash by the fungal entomopathogen Beauveria bassiana (Ascomycota: Hypocreales) for managing Zucchini yellow mosaic virus in cucurbits. Biocontrol Sci. Technol. 2014, 24, 1096–1109. [Google Scholar] [CrossRef]
- Jaber, L.R.; Ownley, B. Can we use entomopathogenic fungi as endophytes for dual biological control of insect pests and plant pathogens? Biol. Control 2018, 116, 36–45. [Google Scholar] [CrossRef]
- Chang, Y.; Xia, X.; Sui, L.; Kang, Q.; Lu, Y.; Li, L.; Liu, W.; Li, Q.; Zhang, Z. Endophytic colonization of entomopathogenic fungi increases plant disease resistance by changing the endophytic bacterial community. J. Basic Microbiol. 2021, 61, 1098–1112. [Google Scholar] [CrossRef] [PubMed]
- Sasan, R.K.; Bidochka, M.J. The insect-pathogenic fungus Metarhizium robertsii (Clavicipitaceae) is also an endophyte that stimulates plant root development. Am. J. Bot. 2012, 99, 101–107. [Google Scholar] [CrossRef]
- Jaber, L.R.; Enkerli, J. Fungal entomopathogens as endophytes: Can they promote plant growth? Biocontrol Sci. Technol. 2017, 27, 28–41. [Google Scholar] [CrossRef]
- Dara, S.K. Non-entomopathogenic roles of entomopathogenic fungi in promoting plant health and growth. Insects 2019, 10, 277. [Google Scholar] [CrossRef]
- Bamisile, B.S.; Siddiqui, J.A.; Akutse, K.S.; Aguila, L.C.R.; Xu, Y. General limitations to endophytic entomopathogenic fungi use as plant growth promoters, pests and pathogens biocontrol agents. Plants 2021, 10, 2119. [Google Scholar] [CrossRef] [PubMed]
- Bruck, D.J. Fungal entomopathogens in the rhizosphere. In The Ecology of Fungal Entomopathogens; Springer: Dordrecht, The Netherlands, 2009; pp. 103–112. [Google Scholar]
- Pava-Ripoll, M.; Angelini, C.; Fang, W.; Wang, S.; Posada, F.J.; Leger, R.S. The rhizosphere-competent entomopathogen Metarhizium anisopliae expresses a specific subset of genes in plant root exudate. Microbiology 2011, 157, 47–55. [Google Scholar] [CrossRef]
- Nelly, N.; Syahrawati, M.; Hamid, H.; Habazar, T.; Gusnia, D.N. Diversity and characterization of entomopathogenic fungi from rhizosphere of maize plants as potential biological control agents. Biodivers. J. Biol. Divers. 2019, 20, 1435–1441. [Google Scholar] [CrossRef]
- De Faria, M.R.; Wraight, S.P. Mycoinsecticides and Mycoacaricides: A comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control 2007, 43, 237–256. [Google Scholar] [CrossRef]
- Quesada-Moraga, E.; López-Díaz, C.; Landa, B.B. The hidden habit of the entomopathogenic fungus Beauveria bassiana: First demonstration of vertical plant transmission. PLoS ONE 2014, 9, e89278. [Google Scholar] [CrossRef]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Vega, F.E.; Goettel, M.S.; Blackwell, M.; Chandler, D.; Jackson, M.A.; Keller, S.; Koike, M.; Maniania, N.K.; Monzón, A.; Ownley, B.; et al. Fungal entomopathogens: New insights on their ecology. Fungal Ecol. 2009, 2, 149–159. [Google Scholar] [CrossRef]
- Inglis, G.D.; Goettel, M.S.; Butt, T.M.; Strasser, H.; Helen, E.R.; Fernando, E.V.; Dave, C.; Mark SGoettel, J.P.; Eric, W. Use of Hyphomycetous Fungi for Managing Insect Pests, Fungi as Biocontrol Agents; Springer: Berlin, Germany, 2001; pp. 23–69. [Google Scholar]
- Akello, J.; Sikora, R. Systemic acropedal influence of endophyte seed treatment on Acyrthosiphon pisum and Aphis fabae offspring development and reproductive fitness. Biol. Control 2012, 61, 215–221. [Google Scholar] [CrossRef]
- Greenfield, M.; Gómez-Jiménez, M.I.; Ortiz, V.; Vega, F.E.; Kramer, M.; Parsa, S. Beauveria bassiana and Metarhizium anisopliae endophytically colonize cassava roots following soil drench inoculation. Biol. Control 2016, 95, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Mantzoukas, S.; Chondrogiannis, C.; Grammatikopoulos, G. Effects of three endophytic entomopathogens on sweet sorghum and on the larvae of the stalk borer Sesamia nonagrioides. Entomol. Exp. Appl. 2015, 154, 78–87. [Google Scholar] [CrossRef]
- Kang, S.C.; Bark, Y.G.; Lee, D.G.; Kim, Y.H. Antifungal activities of Metarhizium anisopliae against Fusarium oxysporum, Botrytis cinerea, and Alternaria solani. Korean J. Mycol. 1996, 24, 49–55. [Google Scholar]
- Shahid, A.; Rao, Q.; Bakhsh, A.; Husnain, T. Entomopathogenic fungi as biological controllers: New insights into their virulence and pathogenicity. Arch. Biol. Sci. 2012, 64, 21–42. [Google Scholar] [CrossRef]
- Wakil, W.; Tahir, M.; Al-Sadi, A.M.; Shapiro-Ilan, D. Interactions between Two Invertebrate Pathogens: An Endophytic Fungus and an Externally Applied Bacterium. Front. Microbiol. 2020, 11, 2624. [Google Scholar] [CrossRef]
- Rodriguez, R.; White, J., Jr.; Arnold, A.; Redman, R. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef]
- Vidal, S.; Jaber, L.R. Entomopathogenic fungi as endophytes: Plant–endophyte–herbivore interactions and prospects for use in biological control. Curr. Sci. 2015, 109, 46–54. [Google Scholar]
- Kambrekar, D.N.; Aruna, J. Screening for endophytic Beauveria bassiana from different plants and its pathogenecity against chickpea pod borer, Helicoverpa armigera (Hübner). J. Exp. Zool. 2018, 21, 727–731. [Google Scholar]
- Gurulingappa, P.; Sword, G.; Murdoch, G.; McGee, P.A. Colonization of crop plants by fungal entomopathogens and their effects on two insect pests when in planta. Biol. Control 2010, 55, 34–41. [Google Scholar] [CrossRef]
- Mantzoukas, S.; Lagogiannis, I. Endophytic colonization of pepper (Capsicum annum) controls aphids (Myzus persicae Sulzer). Appl. Sci. 2019, 9, 2239. [Google Scholar] [CrossRef]
- Bills, G.; Polishook, J.D. Microfungi from Carpinus caroliniana. Can. J. Bot. 1991, 69, 1477–1482. [Google Scholar] [CrossRef]
- Posada, F.; Aime, M.C.; Peterson, S.W.; Rehner, S.A.; Vega, F.E. Inoculation of coffee plants with the fungal entomopathogen Beauveria bassiana (Ascomycota: Hypocreales). Mycol. Res. 2007, 111, 748–757. [Google Scholar] [CrossRef] [PubMed]
- Biswas, C.; Dey, P.; Satpathy, S.; Satya, P. Establishment of the fungal entomopathogen Beauveria bassiana as a season long endophyte in jute (Corchorus olitorius) and its rapid detection using SCAR marker. Entomophaga 2012, 57, 565–571. [Google Scholar] [CrossRef]
- Biswas, C.; Dey, P.; Satpathy, S.; Satya, P.; Mahapatra, B.S. Endophytic colonization of white jute (Corchorus capsularis) plants by different Beauveria bassiana strains for managing stem weevil (Apion corchori). Phytoparasitica 2013, 41, 17–21. [Google Scholar] [CrossRef]
- Guesmi-Jouini, J.; Garrido-Jurado, I.; López-Díaz, C.; Ben Halima-Kamel, M.; Quesada-Moraga, E. Establishment of fungal entomopathogens Beauveria bassiana and Bionectria ochroleuca (Ascomycota: Hypocreales) as endophytes on artichoke Cynara scolymus. J. Invertebr. Pathol. 2014, 119, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.D. Aspects of the Biology and Biological Control of the European Corn Borer in North Carolina. Ph.D. Thesis, Department of Entomology, North Carolina State University, Raleigh, NC, USA, 1994. [Google Scholar]
- Russo, M.L.; Pelizza, S.A.; Cabello, M.N.; Stenglein, S.A.; Scorsetti, A.C. Endophytic colonisation of tobacco, corn, wheat and soybeans by the fungal entomopathogen Beauveria bassiana (Ascomycota, Hypocreales). Biocontrol Sci. Technol. 2014, 25, 475–480. [Google Scholar] [CrossRef]
- Griffin, M.R. Beauveria bassiana, a Cotton Endophyte with Biocontrol Activity against Seedling Disease. Ph.D. Thesis, University of Tennessee, Knoxville, TN, USA, 2007. [Google Scholar]
- Akello, J.; Dubois, T.; Coyne, D.; Kyamanywa, S. Endophytic Beauveria bassiana in banana (Musa spp.) reduces banana weevil (Cosmopolites sordidus) fitness and damage. Crop Prot. 2008, 27, 1437–1441. [Google Scholar] [CrossRef]
- Quesada Moraga, E.; Muñoz-Ledesma, F.J.; Santiago-Álvarez, C. Systemic Protection of Papaver somniferum L. against Iraella luteipes (Hymenoptera: Cynipidae) by an endophytic strain of Beauveria bassiana (Ascomycota: Hypocreales). Environ. Entomol. 2009, 38, 723–730. [Google Scholar] [CrossRef]
- Gómez-Vidal, S.; Lopez-Llorca, L.; Jansson, H.-B.; Salinas, J.; Gómez-Vidal, S.; Lopez-Llorca, L.; Jansson, H.-B.; Salinas, J. Endophytic colonization of date palm (Phoenix dactylifera L.) leaves by entomopathogenic fungi. Micron 2006, 37, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Ganley, R.J.; Newcombe, G. Fungal endophytes in seeds and needles of Pinus monticola. Mycol. Res. 2006, 110, 318–327. [Google Scholar] [CrossRef]
- Reay, S.; Brownbridge, M.; Gicquel, B.; Cummings, N.; Nelson, T. Isolation and characterization of endophytic Beauveria spp. (Ascomycota: Hypocreales) from Pinus radiata in New Zealand forests. Biol. Control 2010, 54, 52–60. [Google Scholar] [CrossRef]
- Donga, T.; Meadow, R.; Meyling, N.; Klingen, I. Natural Occurrence of Entomopathogenic Fungi as Endophytes of Sugarcane (Saccharum officinarum) and in Soil of Sugarcane Fields. Insects 2021, 12, 160. [Google Scholar] [CrossRef]
- Silva, A.C.L.; Silva, G.A.; Abib, P.H.N.; Carolino, A.T.; Samuels, R.I. Endophytic colonization of tomato plants by the entomopathogenic fungus Beauveria bassiana for controlling the South American tomato pinworm, Tuta absoluta. CABI Agric. Biosci. 2020, 1, 3. [Google Scholar] [CrossRef]
- Powell, W.A.; Klingeman, W.; Ownley, B.; Gwinn, K.D. Evidence of endophytic Beauveria bassiana in seed-treated tomato plants acting as a systemic entomopathogen to larval helicoverpa zea (Lepidoptera: Noctuidae). J. Entomol. Sci. 2009, 44, 391–396. [Google Scholar] [CrossRef]
- Toffa, J.; Loko, Y.L.E.; Kpindou, O.K.D.; Zanzana, K.; Adikpeto, J.; Gbenontin, Y.; Koudamiloro, A.; Adandonon, A. Endophytic colonization of tomato plants by Beauveria bassiana Vuillemin (Ascomycota: Hypocreales) and leaf damage in Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) larvae. Egypt. J. Biol. Pest Control 2021, 31, 82. [Google Scholar] [CrossRef]
- Mantzoukas, S. Endophytic colonization of Solanum tuberosum L. (Solanales: Solanaceae) plants can affect the infestation of serious pests. Appl. Microbiol. Theory Technol. 2020, 1, 52–58. [Google Scholar]
- Reddy, N.P.; Khan, A.P.A.; Devi, U.K.; Sharma, H.C.; Reineke, A. Treatment of millet crop plant (Sorghum bicolor) with the entomopathogenic fungus (Beauveria bassiana) to combat infestation by the stem borer, Chilo partellus Swinhoe (Lepidoptera: Pyralidae). J. Asia-Pac. Entomol. 2009, 12, 221–226. [Google Scholar] [CrossRef]
- Posada, F.; Vega, F.E. Establishment of the fungal entomopathogen Beauveria bassiana (Ascomycota: Hypocreales) as an endophyte in cocoa seedlings (Theobroma cacao). Mycologia 2005, 97, 1195–1200. [Google Scholar] [CrossRef]
- Evans, H.C.; Holmes, K.; Thomas, S.E. Endophytes and mycoparasites associated with an indigenous forest tree, Theobroma gileri, in Ecuador and a preliminary assessment of their potential as biocontrol agents of cocoa diseases. Mycol. Prog. 2003, 2, 149–160. [Google Scholar] [CrossRef]
- Jaber, L.R.; Enkerli, J. Effect of seed treatment duration on growth and colonization of Vicia faba by endophytic Beauveria bassiana and Metarhizium brunneum. Biol. Control 2016, 103, 187–195. [Google Scholar] [CrossRef]
- Jaber, L.R. Grapevine leaf tissue colonization by the fungal entomopathogen Beauveria bassiana s.l. and its effect against downy mildew. Entomophaga 2015, 60, 103–112. [Google Scholar] [CrossRef]
- Bing, L.A.; Lewis, L.C. Suppression of Ostrinia nubilalis (Hübner) (Lepidoptera: Pyralidae) by Endophytic Beauveria bassiana (Balsamo) Vuillemin. Environ. Entomol. 1991, 20, 1207–1211. [Google Scholar] [CrossRef]
- Cherry, A.J.; Banito, A.; Djegui, D.; Lomer, C. Suppression of the stem-borer Sesamia calamistis (Lepidoptera; Noctuidae) in maize following seed dressing, topical application and stem injection with African isolates of Beauveria bassiana. Int. J. Pest Manag. 2004, 50, 67–73. [Google Scholar] [CrossRef]
- Mantzoukas, S.; Grammatikopoulos, G. The effect of three entomopathogenic endophytes of the sweet sorghum on the growth and feeding performance of its pest, Sesamia nonagrioides larvae, and their efficacy under field conditions. Crop Prot. 2020, 127, 104952. [Google Scholar] [CrossRef]
- Arnold, A.E.; Lewis, L.C. Ecology and evolution of fungal endophytes, and their roles against insects. In Insect-Fungal Associations: Ecology and Evolution; Oxford University Press: New York, NY, USA, 2005; pp. 74–96. [Google Scholar]
- Akello, J.; Dubois, T.; Coyne, D.; Kyamanywa, S. Effect of endophytic Beauveria bassiana on populations of the banana weevil, Cosmopolites sordidus, and their damage in tissue-cultured banana plants. Entomol. Exp. Appl. 2008, 129, 157–165. [Google Scholar] [CrossRef]
- Vega, F. The use of fungal entomopathogens as endophytes in biological control: A review. Mycologia 2018, 110, 4–30. [Google Scholar] [CrossRef]
- Bamisile, B.S.; Akutse, K.S.; Dash, C.K.; Qasim, M.; Aguila, L.C.R.; Ashraf, H.J.; Huang, W.; Hussain, M.; Chen, S.; Wang, L. Effects of seedling age on colonization patterns of citrus limon plants by endophytic Beauveria bassiana and Metarhizium anisopliae and their influence on seedlings growth. J. Fungi 2020, 6, 29. [Google Scholar] [CrossRef]
- Keller, S.; Zimmermann, G. Mycopathogens of soil. In Insect-Fungus Interactions; Wilding, N., Collins, N.M., Hammond, P.M., Webber, J.F., Eds.; Academic Press: London, UK, 1989; pp. 239–270. [Google Scholar]
- Jaronski, S.T. Ecological factors in the inundative use of fungal entomopathogens. BioControl 2020, 55, 159–185. [Google Scholar] [CrossRef]
- Strasser, H.; Vey, A.; Butt, T.M. Are there any risks in using entomopathogenic fungi for pest control, with particular reference to the bioactive metabolites of metarhizium, tolypocladium and beauveria species? Biocontrol Sci. Technol. 2000, 10, 717–735. [Google Scholar] [CrossRef]
- Zimmermann, G. Review on safety of the entomopathogenic fungi Beauveria bassiana and Beauveria brongniartii. Biocontrol Sci. Technol. 2007, 17, 553–596. [Google Scholar] [CrossRef]
- Zimmermann, G. Review on safety of the entomopathogenic fungus Metarhizium anisopliae. Biocontrol Sci. Technol. 2007, 17, 879–920. [Google Scholar] [CrossRef]
- Vega, F.E. Insect pathology and fungal endophytes. J. Invertebr. Pathol. 2008, 98, 277–279. [Google Scholar] [CrossRef]
- Fargues, J.; Goettel, M.S.; Smits, N.; Ouedraogo, A.; Vidal, C.; Lacey, L.A.; Lomer, C.J.; Rougier, M. Variability in susceptibility to simulated sunlight of conidia among isolates of entomopathogenic Hyphomycetes. Mycopathol. Mycol. Appl. 1996, 135, 171–181. [Google Scholar] [CrossRef]
- Moore, D.; Lord, J.C.; Smith SMSubramanyam, B.; Hagstrum, D.W. Pathogens. In Alternatives to Pesticides in Stored-Product IPM; Springer Science & Business Media: New York, NY, USA, 2000. [Google Scholar]
- Mantzoukas, S.; Lagogiannis, I.; Ntoukas, A.; Eliopoulos, P.A.; Kouretas, D.; Karpouzas, D.G.; Poulas, K. Trapping entomopathogenic fungi from vine terroir soil samples with insect baits for controlling serious pests. Appl. Sci. 2020, 10, 3539. [Google Scholar] [CrossRef]
- Inglis, G.D.; Goettel, M.S.; Erlandson, M.A.; Weaver, D.K. Field Manual of Techniques in Invertebrate Pathology. In Grasshoppers and Locusts; Springer: Dordrecht, The Netherlands, 2007; pp. 627–654. [Google Scholar]
- Zhang, W.; Meng, J.; Ning, J.; Qin, P.; Zhou, J.; Zou, Z.; Wang, Y.; Jiang, H.; Ahmad, F.; Zhao, L.; et al. Differential immune responses of Monochamus alternatus against symbiotic and entomopathogenic fungi. Sci. China Life Sci. 2017, 60, 902–910. [Google Scholar] [CrossRef]
- Mantzoukas, S.; Lagogiannis, I.; Mpekiri, M.; Pettas, I.; Eliopoulos, P.A. Insecticidal action of several isolates of entomopathogenic fungi against the granary weevil Sitophilus granarius. Agriculture 2019, 9, 222. [Google Scholar] [CrossRef]
- Rodrigues, C.; Pratissoli, D. Pathogenicity of Beauveria brongniartii (Sacc.) Petch and Metarhizium anisopliae (Mots.) Sorok and their effect on the corn weevil and the bean beetle. An. Soc. Entomol. Bras. 1990, 19, 301–306. [Google Scholar] [CrossRef]
- Moino, A., Jr.; Alves, S.B.; Pereira, R.M. Efficacy of Beauveria bassiana (Balsamo) Vuillemin isolates for control of stored-grain pests. J. Appl. Entomol. 1998, 122, 301–305. [Google Scholar] [CrossRef]
- Batta, Y. Control of the lesser grain borer (Rhyzopertha dominica (F.), Coleoptera: Bostrichidae) by treatments with residual formulations of Metarhizium anisopliae (Metschnikoff) Sorokin (Deuteromycotina: Hyphomycetes). J. Stored Prod. Res. 2005, 41, 221–229. [Google Scholar] [CrossRef]
- Ignoffo, C.M. The fungus Nomuraea rileyi as a microbial insecticide. Microb. Control Pests Plant Dis. 1970, 1980, 513–538. [Google Scholar]
- Ignoffo, C.M.; Hostetter, D.L.; Sikorowski, P.P.; Sutter, G.; Brooks, W.M. Inactivation of representative species of entomopathogenic viruses, a bacterium, fungus, and protozoan by an ultraviolet light source. Environ. Entomol. 1977, 6, 411–415. [Google Scholar] [CrossRef]
- Meyling, N.V.; Eilenberg, J. Occurrence and distribution of soil borne entomopathogenic fungi within a single organic agroecosystem. Agric. Ecosyst. Environ. 2006, 113, 336–341. [Google Scholar] [CrossRef]
- Butt, T.M.; Goettel, M.S. Bioassays of entomogenous fungi. In Bioassays of Entomopathogenic Microbes and Nematodes; CAB International: Wallingford, UK, 2000; pp. 141–195. [Google Scholar] [CrossRef]
- Fernandes, E.; Rangel, D.E.; Moraes, Á.M.; Bittencourt, V.R.; Roberts, D.W. Cold activity of Beauveria and Metarhizium, and thermotolerance of Beauveria. J. Invertebr. Pathol. 2008, 98, 69–78. [Google Scholar] [CrossRef]
- Bayissa, W.; Ekesi, S.; Mohamed, S.A.; Kaaya, G.P.; Wagacha, J.M.; Hanna, R.; Maniania, N.K. Selection of fungal isolates for virulence against three aphid pest species of crucifers and okra. J. Pest Sci. 2017, 90, 355–368. [Google Scholar] [CrossRef]
- Batta, Y.A.; Kavallieratos, N.G. The use of entomopathogenic fungi for the control of stored-grain insects. Int. J. Pest Manag. 2017, 64, 77–87. [Google Scholar] [CrossRef]
- Batta, Y.A. Control of main stored-grain insects with new formulations of entomopathogenic fungi in diatomaceous earth dusts. Int. J. Food Eng. 2008, 4, 9. [Google Scholar] [CrossRef]
- Bengston, M.; Cooper, L.M.; Grant-Taylor, F.J. A comparison of bioresmethrin, chlorpyrifos-methyl and pirimiphos methyl as grain protectants against malathion-resistant insects in wheat. Qld. J. Agric. Anim. Sci. 1975, 32, 51–78. [Google Scholar]
- Arthur, F.H. Control of Lesser Grain Borer (Coleoptera: Bostrichidae) with Chlorpyrifos-Methyl, Bioresmethrin, and Resmethrin: Effect of Chlorpyrifos-Methyl Resistance and Environmental Degradation. J. Econ. Entomol. 1992, 85, 1471–1475. [Google Scholar] [CrossRef]
- Tyler, P.S.; Taylor, R.W.; Rees, D.P. Insect resistance to phosphine fumigations in food warehouses in Pangladesh. Int. Pest Control. 1983, 25, 10–13. [Google Scholar]
- Zettler, L.J.; Leesch, J.G.; Gill, R.F.; Mackey, B.E. Toxicity of carbonyl sulfide to stored product insects. J. Econ. Entomol. 1997, 90, 832–836. [Google Scholar] [CrossRef]
- Zettler, J.; Arthur, F.H. Chemical control of stored product insects with fumigants and residual treatments. Crop Prot. 2000, 19, 577–582. [Google Scholar] [CrossRef]
- Mantzoukas, S.; Lagogiannis, I.; Karmakolia, K.; Rodi, A.; Gazepi, M.; Eliopoulos, P.A. The effect of grain type on virulence of entomopathogenic fungi against stored product pests. Appl. Sci. 2020, 10, 2970. [Google Scholar] [CrossRef]
- Adane, K.; Moore, D.; Archer, S. Preliminary studies on the use of Beauveria bassiana to control Sitophilus zeamais (Coleoptera: Curculionidae) in the laboratory. J. Stored Prod. Res. 1996, 32, 105–113. [Google Scholar] [CrossRef]
- Padin, S.B.; Bello, G.M.; Vasicek, A.L. Pathogenicity of Beauveria bassiana for adults of Tribolium castaneum (Col.: Tenebrionidae) in stored grains. Entomophaga 1997, 42, 569–574. [Google Scholar] [CrossRef]
- Padín, S.; Bello, G.D.; Fabrizio, M. Grain loss caused by Tribolium castaneum, Sitophilus oryzae and Acanthoscelides obtectus in stored durum wheat and beans treated with Beauveria bassiana. J. Stored Prod. Res. 2002, 38, 69–74. [Google Scholar] [CrossRef]
- Hidalgo, E.; Moore, D.; le Patourel, G. The effect of different formulations of Beauveria bassiana on Sitophilus zeamais in stored maize. J. Stored Prod. Res. 1998, 34, 171–179. [Google Scholar] [CrossRef]
- Rice, W.C.; Cogburn, R.R. Activity of the entomopathogenic fungus Beauveria bassiana (Deuteromycota: Hyphomycetes) against three coleopteran pests of stored grain. J. Econ. Entomol. 1999, 92, 691–694. [Google Scholar] [CrossRef]
- Bello, G.D.; Padin, S.; Lastra, C.L.; Fabrizio, M. Laboratory evaluation of chemical-biological control of the rice weevil (Sitophilus oryzae L.) in stored grains. J. Stored Prod. Res. 2000, 37, 77–84. [Google Scholar] [CrossRef]
- Meikle, W.; Cherry, A.; Holst, N.; Hounna, B.; Markham, R. The Effects of an Entomopathogenic Fungus, Beauveria bassiana (Balsamo) Vuillemin (Hyphomycetes), on Prostephanus truncatus (Horn) (Col.: Bostrichidae), Sitophilus zeamais Motschulsky (Col.: Curculionidae), and Grain Losses in Stored Maize in the Benin Republic. J. Invertebr. Pathol. 2001, 77, 198–205. [Google Scholar] [CrossRef]
- Lord, J.C. Desiccant Dusts Synergize the Effect of Beauveria bassiana (Hyphomycetes: Moniliales) on Stored-Grain Beetles. J. Econ. Entomol. 2001, 94, 367–372. [Google Scholar] [CrossRef]
- Sheeba, G.; Seshadri, S.; Raja, N.; Janarthanan, S.; Ignacimuthu, S. Efficacy of Beauveria bassiana for control of the rice weevil Sitophilus oryzae (L.) (Coleoptera: Curculionidae). Appl. Entomol. Zool. 2001, 36, 117–120. [Google Scholar] [CrossRef]
- Vassilakos, T.; Athanassiou, C.; Kavallieratos, N.; Vayias, B. Influence of temperature on the insecticidal effect of Beauveria bassiana in combination with diatomaceous earth against Rhyzopertha dominica and Sitophilus oryzae on stored wheat. Biol. Control 2006, 38, 270–281. [Google Scholar] [CrossRef]
- Athanassiou, C.; Steenberg, T. Insecticidal effect of Beauveria bassiana (Balsamo) Vuillemin (Ascomycota: Hypocreaes) in combination with three diatomaceous earth formulations against Sitophilus granarius (L.) (Coleoptera: Curculionidae). Biol. Control 2007, 40, 411–416. [Google Scholar] [CrossRef]
- Akbar, W.; Lord, J.C.; Nechols, J.R.; Howard, R.W. Diatomaceous earth increases the efficacy of Beauveria bassiana against Tribolium castaneum larvae and increases conidia attachment. J. Econ. Entomol. 2004, 97, 273–2820. [Google Scholar] [CrossRef]
- Kalita, S.; Hazarika, L.K.; Das, P.; Das, K. Efficacy of Chromolaena odorata leaf powder and Beauveria bassiana against stored grain pests. Indian J. Entomol. 2018, 80, 1005. [Google Scholar] [CrossRef]
- Afandhi, A.; Pratiwi, V.R.; Hadi, M.S.; Setiawan, Y.; Puspitarini, R.D. Suitable Combination Between Beauveria bassiana (Balsamo) Vuillemin and Four Plant Leaf Extracts to Control Spodoptera litura (Fabricius). AGRIVITA J. Agric. Sci. 2020, 42, 341. [Google Scholar] [CrossRef]
- Depieri, R.A.; Martinez, S.S.; Menezes, A.O., Jr. Compatibility of the fungus Beauveria bassiana (Bals.) Vuill. (Deuteromycetes) with extracts of neem seeds and leaves and the emulsible oil. Neotrop. Entomol. 2005, 34, 601–606. [Google Scholar] [CrossRef]
- Mantzoukas, S.; Zikou, A.; Triantafillou, V.; Lagogiannis, I.; Eliopoulos, P.A. Ιnteractions between Beauveria bassiana and Isaria fumosorosea and Their Hosts Sitophilus granarius (L.) and Sitophilus oryzae (L.) (Coleoptera: Curculionidae). Insects 2019, 10, 362. [Google Scholar] [CrossRef]
- Batool, R.; Umer, M.; Wang, Y.; He, K.; Zhang, T.; Bai, S.; Zhi, Y.; Chen, J.; Wang, Z. Synergistic Effect of Beauveria bassiana and Trichoderma asperellum to Induce Maize (Zea mays L.) Defense against the Asian Corn Borer, Ostrinia furnacalis (Lepidoptera, Crambidae) and Larval Immune Response. Int. J. Mol. Sci. 2020, 21, 8215. [Google Scholar] [CrossRef]
- Mantzoukas, S.; Milonas, P.; Kontodimas, D.; Angelopoulos, K. Interaction between the entomopathogenic bacterium Bacillus thuringiensis subsp. kurstaki and two entomopathogenic fungi in biocontrol of Sesamia nonagrioides (Lefebvre) (Lepidoptera: Noctuidae). Ann. Microbiol. 2013, 63, 1083–1091. [Google Scholar] [CrossRef]
- Wraight, S.P.; Ramos, M.E. Synergistic interaction between Beauveria bassiana and Bacillus thuringiensis tenebrionis-based biopesticides applied against field populations of Colorado potato beetle larvae. J. Invertebr. Pathol. 2005, 90, 139–150. [Google Scholar] [CrossRef]
- Pauli, G.; Mascarin, G.M.; Eilenberg, J.; Delalibera, I., Jr. Within-Host Competition between Two Entomopathogenic Fungi and a Granulovirus in Diatraea saccharalis (Lepidoptera: Crambidae). Insects 2018, 9, 64. [Google Scholar] [CrossRef]
- Ansari, M.A.; Shah, F.A.; Tirry, L.; Moens, M. Field trials against Hoplia philanthus (Coleoptera: Scarabaeidae) with a combination of an entomopathogenic nematode and the fungus Metarhizium anisopliae CLO 53. Biol. Control 2006, 39, 453–459. [Google Scholar] [CrossRef]
- Kavallieratos, N.G.; Athanassiou, C.G.; Michalaki, M.P.; Batta, Y.A.; Rigatos, H.A.; Pashalidou, F.G.; Balotis, G.N.; Tomanovic, Z.; Vayias, B.J. Effect of the combined use of Metarhizium anisopliae (Metschinkoff) Sorokin and diatomaceous earth for the control of three stored-product beetle species. Crop Prot. 2006, 25, 1087–1094. [Google Scholar] [CrossRef]
- Dakhel, W.H.; Latchininsky, A.V.; Jaronski, S.T. Efficacy of two entomopathogenic fungi, Metarhizium brunneum, strain F52 alone and combined with Paranosema locustae against the migratory grasshopper, Melanoplus sanguinipes, under laboratory and greenhouse conditions. Insects 2010, 10, 94. [Google Scholar] [CrossRef]
- Uma Maheswara Rao, C.; Uma Devi, K.; Akbar Ali Khan, P. Effect of combination treatment with entomopathogenic fungi Beauveria bassiana and Nomuraea rileyi (Hypocreales) on Spodoptera litura (Lepidoptera: Noctuidaeae). Biocontrol Sci. Technol. 2006, 16, 221–232. [Google Scholar] [CrossRef]


{kind=link}
| Host Plants | Insecticide Activity | Fungicide Activity | References |
|---|---|---|---|
| Brassica napus L. (Brassicaceae) | [30] | ||
| Brassica oleracea L. (Brassicaceae) | Aphis fabae | [31,32] | |
| Capsicum spp. (Solanaceae) | Myzus persicae | [31,33] | |
| Carpinus caroliniana Walter (Betulaceae) | [34] | ||
| Cicer arietinum L. (Fabaceae) | [31] | ||
| Citrus spp. (Rutaceae) | [31] | ||
| Coffea arabica L. (Rubiaceae) | [35] | ||
| Corchorus olitorius L. (Malvaceae) | [36] | ||
| Corchorus capsularis L. (Malvaceae) | Apion corchori | [37] | |
| Cucurbita maxima L. (Cucurbitaceae) | Aphis gossipii | [32] | |
| Cucurbita pepo L. (Cucurbitaceae) | [8] | ||
| Cynara scolymus L. (Asteraceae) | [38] | ||
| Datura stramonium L. (Solanaceae) | [39] | ||
| Glycine max (L.) Merr. (Fabaceae) | [40] | ||
| Gossypium hirsutum L. (Malvaceae) | Aphis gossipii | Pythium myriotylum | [32,39,41] |
| Helianthus annuus L. (Asteraceae) | [31] | ||
| Manihot esculenta Crantz (Euphorbiaceae) | [24] | ||
| Musa spp. (Musaceae) | Cosmopolites sordidus | [31,42] | |
| Nicotiana tabacum L. (Solanaceae); | [39] | ||
| Papaver somniferum L. (Papaveraceae). | Iraella luteipes | [43] | |
| Phaseolus vulgaris L. (Fabaceae) | Helicoverpa armigera | [30,31,32] | |
| Phoenix dactylifera L. (Arecaceae) | [44] | ||
| Pinus monticola Dougl. ex. D. Don (Pinaceae) | [45] | ||
| Pinus radiata D. Don (Pinaceae) | [46] | ||
| Punica granatum L. (Lythraceae) | [31] | ||
| Saccharum officinarum L. (Poaceae) | [47] | ||
| Solanum lycopersicum L. (Solanaceae) | Aphis gossipii, Tuta absoluta, Helicoverpa zea Helicoverpa armigera | X. axonopodis pv. Malvacearum. Rizoctonia solani, Pythium myriotylum | [5,32,40,48,49,50] |
| Solanum melongena L. (Solanaceae) | [31] | ||
| Solanum tuberosum L. (Solanaceae) | Phthorinaea operculla Trialeurodes vaporariorum | [39,51] | |
| Sorghum bicolor L. (Poaceae) | Sesamia nonagrioides, Chilo partelus | [25,52] | |
| Theobroma cacoa L. (Malvaceae) | [53] | ||
| Theobroma gileri Coatrec (Malvaceae) | [54] | ||
| Triticum aestivum L. (Poaceae) | Aphis gossipii | [32] | |
| Vicia faba L. (Fabaceae) | Helicoverpa armigera | [30,55] | |
| Vigna radiata (L.) Wilczek (Fabaceae) | [31] | ||
| Vitis vinifera L. (Vitaceae) | Plasmopara viticola | [56] | |
| Xanthium strumarium L. (Asteraceae) | [39] | ||
| Zea mays L. (Gramineae) | A. gossipii, Sesamia calamistis, Ostrinia nubilalis | [31,32,57,58] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mantzoukas, S.; Kitsiou, F.; Natsiopoulos, D.; Eliopoulos, P.A. Entomopathogenic Fungi: Interactions and Applications. Encyclopedia 2022, 2, 646-656. https://doi.org/10.3390/encyclopedia2020044
Mantzoukas S, Kitsiou F, Natsiopoulos D, Eliopoulos PA. Entomopathogenic Fungi: Interactions and Applications. Encyclopedia. 2022; 2(2):646-656. https://doi.org/10.3390/encyclopedia2020044
Chicago/Turabian StyleMantzoukas, Spiridon, Foteini Kitsiou, Dimitrios Natsiopoulos, and Panagiotis A. Eliopoulos. 2022. "Entomopathogenic Fungi: Interactions and Applications" Encyclopedia 2, no. 2: 646-656. https://doi.org/10.3390/encyclopedia2020044