
Considering Phytosphingosine-Based Ceramide Formulations for Atopic Skin Care
 , , and
, , and 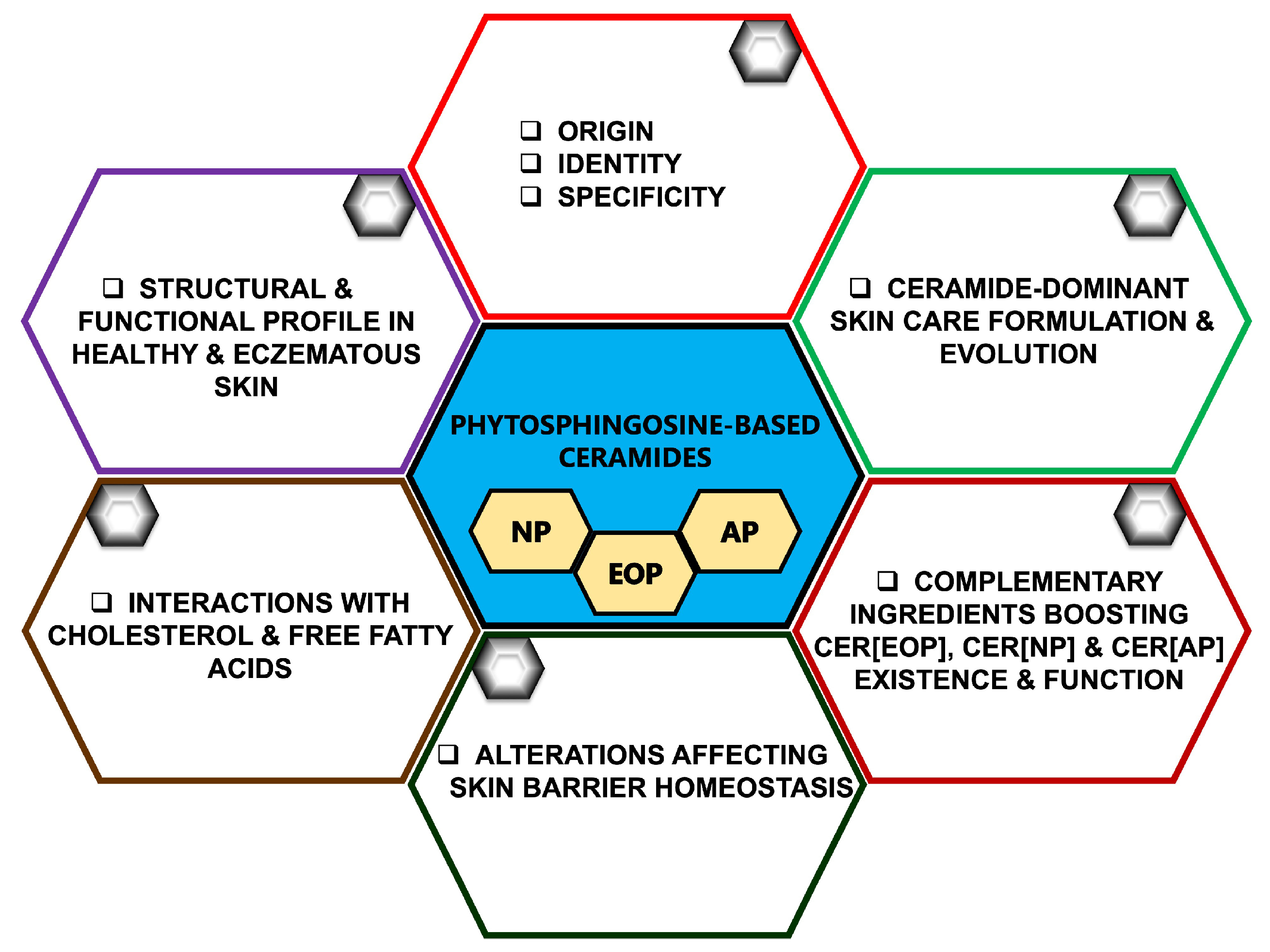{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Composition of the Stratum corneum and the Importance of Ceramides in Skin Health
3. Characterizing the Key Skin Ceramides
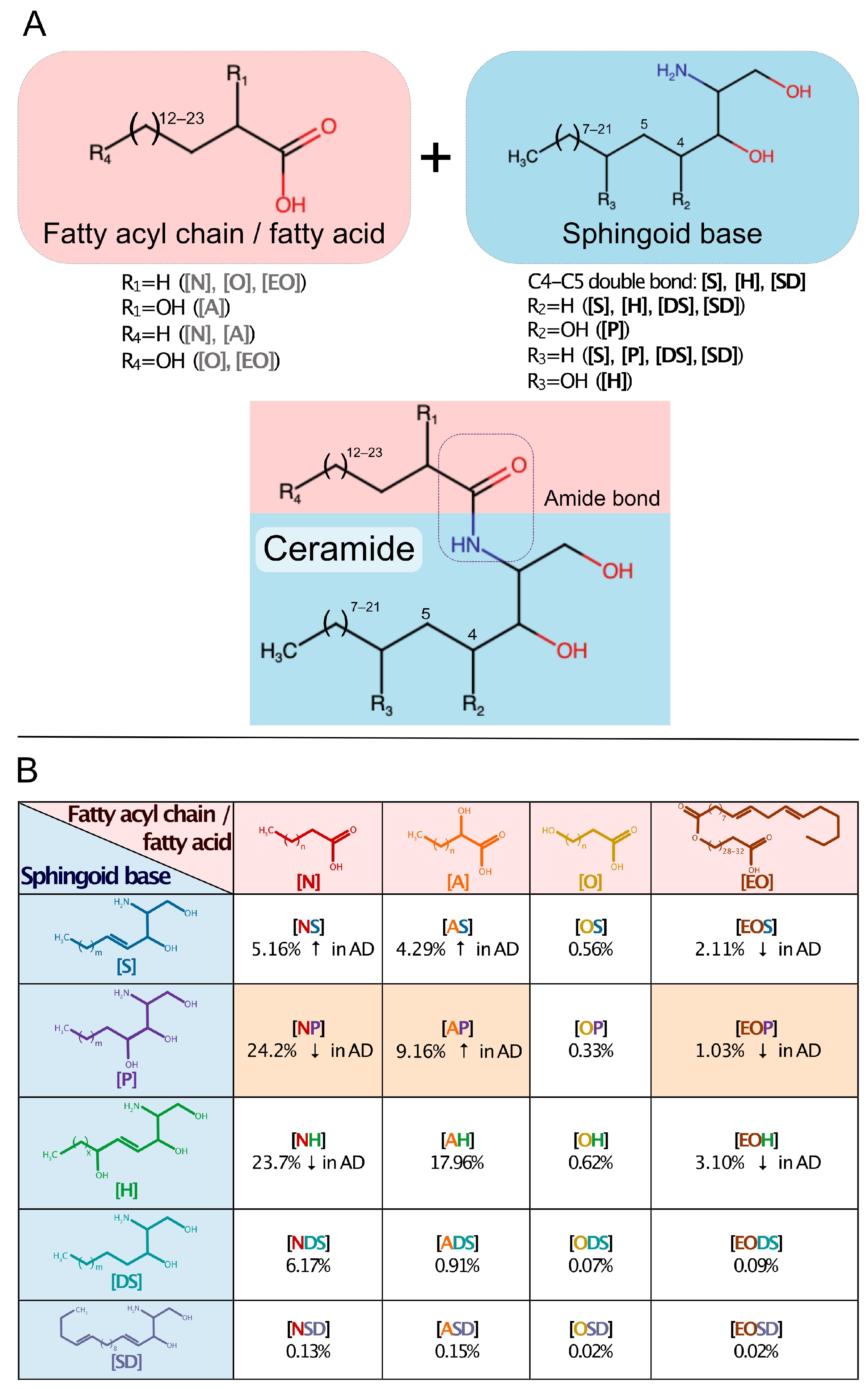
3.1. Origin and Nomenclature

3.2. Ceramide Identity, Functional Diversity, Complexity, and Specificity
3.3. Ceramide Conformational Features and Interactions with Cholesterol and Free Fatty Acids
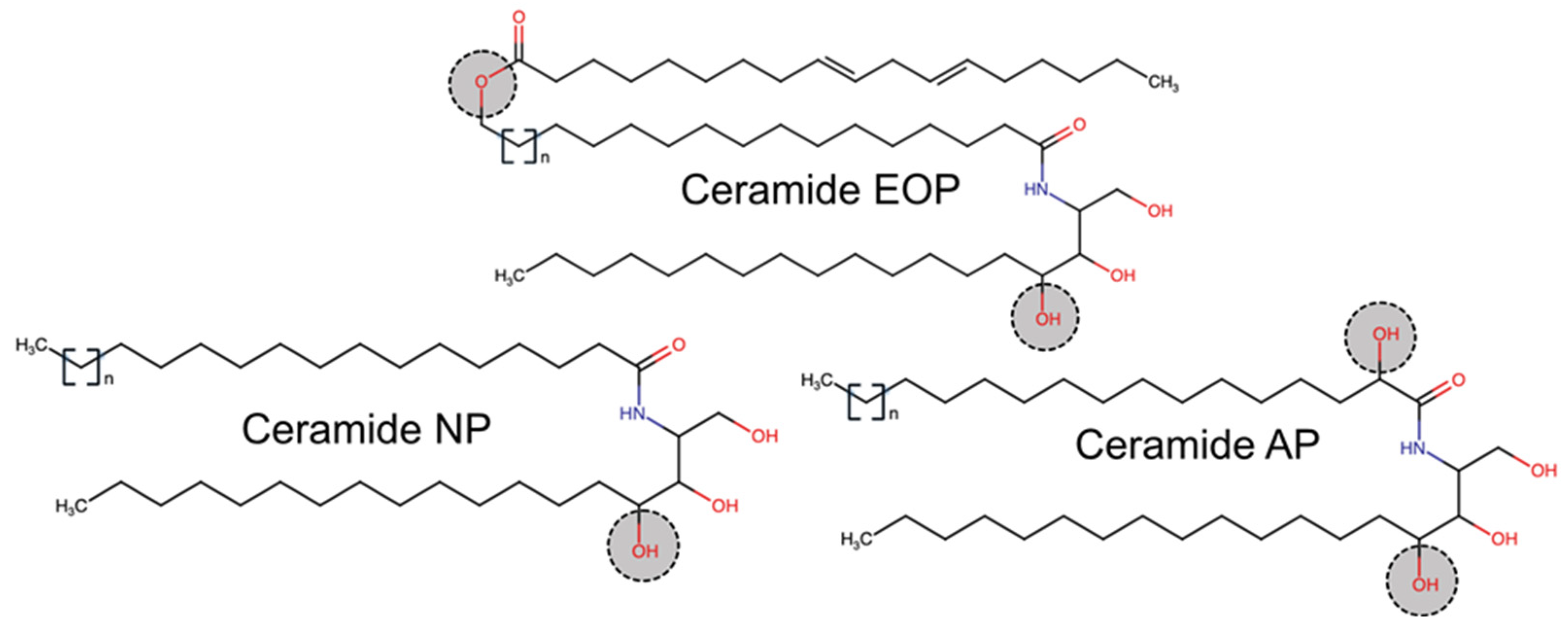
3.4. CER[EOP]
3.5. CER[NP]
3.6. CER[AP]
4. Altered Ceramide Levels in Eczematous Skin
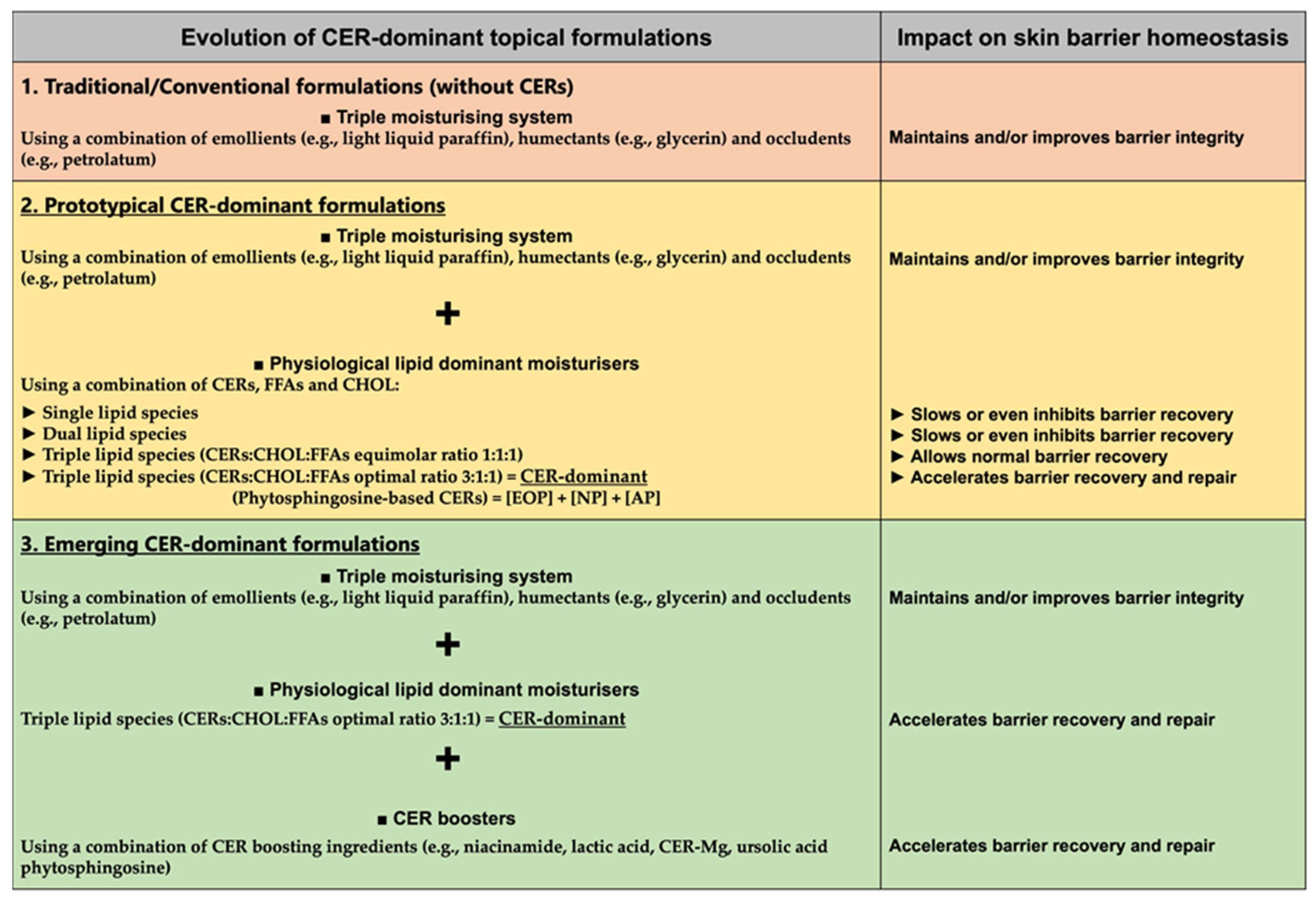
5. Ceramide-Dominant Skin Care Evolution for Skin Barrier Maintenance, Recovery and Repair
6. Considering Ceramide-Dominant Topical Skin Care Formulations
6.1. Niacinamide
6.2. Lactic Acid
6.3. Ceramide–Magnesium (CER-Mg)
6.4. Phytosphingosine
6.5. Ursolic Acid
7. Conclusions and Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Elias, P.M. Stratum corneum defensive functions: An integrated view. J. Investig. Dermatol. 2005, 125, 183–200. [Google Scholar] [CrossRef]
- Sahle, F.F.; Gebre-Mariam, T.; Dobner, B.; Wohlrab, J.; Neubert, R.H. Skin diseases associated with the depletion of stratum corneum lipids and stratum corneum lipid substitution therapy. Skin Pharmacol. Physiol. 2015, 28, 42–55. [Google Scholar] [CrossRef]
- Almeida, C.; Filipe, P.; Rosado, C.; Pereira-Leite, C. Nanodelivery strategies for skin diseases with barrier impairment: Focusing on ceramides and glucocorticoids. Nanomaterials 2022, 12, 275. [Google Scholar] [CrossRef] [PubMed]
- Yadav, N.; Madke, B.; Das, A. Ceramides: Where do we stand? CosmoDerma 2021, 1, 44. [Google Scholar] [CrossRef]
- Knox, S.; O’Boyle, N.M. Skin lipids in health and disease: A review. Chem. Phys. Lipids 2021, 236, 105055. [Google Scholar] [CrossRef] [PubMed]
- Drakou, K.; Tsianni, A.; Vrani, F.; Kefala, V.; Rallis, E. Revealing the correlation between altered skin lipids composition and skin disorders. Cosmetics 2021, 8, 88. [Google Scholar] [CrossRef]
- Zeb, A.; Arif, S.T.; Malik, M.; Shah, F.A.; Din, F.U.; Qureshi, O.S.; Lee, E.-S.; Lee, G.-Y.; Kim, J.-K. Potential of nanoparticulate carriers for improved drug delivery via skin. J. Pharm. Investig. 2019, 49, 485–517. [Google Scholar] [CrossRef]
- Parrado, C.; Mercado-Saenz, S.; Perez-Davo, A.; Gilaberte, Y.; Gonzalez, S.; Juarranz, A. Environmental stressors on skin aging. Mechanistic insights. Front. Pharmacol. 2019, 10, 759. [Google Scholar] [CrossRef] [PubMed]
- Molina-García, M.; Granger, C.; Trullàs, C.; Puig, S. Exposome and skin: Part 1. Bibliometric analysis and review of the impact of exposome approaches on dermatology. Dermatol. Ther. 2022, 12, 345–359. [Google Scholar] [CrossRef] [PubMed]
- Évora, A.S.; Adams, M.J.; Johnson, S.A.; Zhang, Z. Corneocytes: Relationship between structural and biomechanical properties. Skin Pharmacol. Physiol. 2021, 34, 146–161. [Google Scholar] [CrossRef]
- Hatfield, R.M.; Fung, L.W. Molecular properties of a stratum corneum model lipid system: Large unilamellar vesicles. Biophys. J. 1995, 68, 196–207. [Google Scholar] [CrossRef]
- Coderch, L.; López, O.; de la Maza, A.; Parra, J.L. Ceramides and skin function. Am. J. Clin. Dermatol. 2003, 4, 107–129. [Google Scholar] [CrossRef] [PubMed]
- Bouwstra, J.A.; Gooris, G.S. The lipid organisation in human stratum corneum and model systems. Open Dermatol. J. 2010, 4, 10–13. Available online: https://opendermatologyjournal.com/contents/volumes/V4/TODJ-4-10/TODJ-4-10.pdf (accessed on 19 May 2023). [CrossRef]
- Elias, P.M.; Man, M.Q.; Darmstad, G.L. Optimised emollient mixture for skin barrier repair: Applications to global child health. J. Glob. Health 2022, 12, 03019. [Google Scholar] [CrossRef] [PubMed]
- Bonté, F. Skin lipids: Their origin and function. Recent Res. Devel. Lipid Res. 1999, 3, 43–62. Available online: https://hal.science/hal-03316280/document (accessed on 19 May 2023).
- Feingold, K.R. Thematic review series: Skin lipids. The role of epidermal lipids in cutaneous permeability barrier homeostasis. J. Lipid Res. 2007, 48, 2531–2546. [Google Scholar] [CrossRef] [PubMed]
- Haftek, M.; Roy, D.C.; Liao, I.C. Evolution of skin barrier science for healthy and compromised skin. J. Drugs Dermatol. 2021, 20, s3–s9. [Google Scholar] [CrossRef]
- Kahraman, E.; Kaykın, M.; Şahin Bektay, H.; Güngör, S. Recent advances on topical application of ceramides to restore barrier function of skin. Cosmetics 2019, 6, 52. [Google Scholar] [CrossRef]
- Uche, L.E.; Gooris, G.S.; Bouwstra, J.A.; Beddoes, C.M. Increased levels of short-chain ceramides modify the lipid organization and reduce the lipid barrier of skin model membranes. Langmuir 2021, 37, 9478–9489. [Google Scholar] [CrossRef]
- Hatta, I. Stratum corneum structure and function studied by X-ray diffraction. Dermato 2022, 2, 79–108. [Google Scholar] [CrossRef]
- Engelbrecht, T.N.; Schroeter, A.; Hauß, T.; Demé, B.; Scheidt, H.A.; Huster, D.; Neubert, R.H.H. The impact of ceramides NP and AP on the nanostructure of stratum corneum lipid bilayer. Part I: Neutron diffraction and 2H NMR studies on multilamellar models based on ceramides with symmetric alkyl chain length distribution. Soft Matter 2012, 8, 2599–2607. [Google Scholar] [CrossRef]
- Tessema, E.N.; Gebre-Mariam, T.; Neubert, R.H.H.; Wohlrab, J. Potential applications of phyto-derived ceramides in improving epidermal barrier function. Skin Pharmacol. Physiol. 2017, 30, 115–138. [Google Scholar] [CrossRef]
- Schachner, L.A.; Blume-Peytavi, U.; Andriessen, A.; Izakovic, J.; Maruani, A.; Micali, G.; Murashkin, N.; Salavastru, C.; Torrelo, A. Expert consensus on ceramides containing skincare in newborns and infants and potential mitigation of atopic dermatitis. Ital. J. Dermatol. Venerol. 2022, 157, 23–32. [Google Scholar] [CrossRef]
- Iwai, I.; Han, H.; den Hollander, L.; Svensson, S.; Ofverstedt, L.G.; Anwar, J.; Brewer, J.; Bloksgaard, M.; Laloeuf, A.; Nosek, D.; et al. The human skin barrier is organized as stacked bilayers of fully extended ceramides with cholesterol molecules associated with the ceramide sphingoid moiety. J. Investig. Dermatol. 2012, 132, 2215–2225. [Google Scholar] [CrossRef]
- Wollenweber, U.; Farwick, M. Application of skin-identical ceramide 3 for enhanced skin moisturization and smoothness: Latest results. Euro. Cosmet. 2006, 2006, 1–5. Available online: https://www.researchgate.net/publication/291273298_Applicaion_of_skin-identical_ceramide_3_for_enhanced_skin_moisturization_and_smoothness_latest_results (accessed on 19 May 2023).
- Breathnach, A.S.; Goodman, T.; Stolinski, C.; Gross, M. Freeze-fracture replication of cells of stratum corneum of human epidermis. J. Anat. 1973, 114, 65–81. Available online: https://pubmed.ncbi.nlm.nih.gov/4736654/ (accessed on 19 May 2023). [PubMed]
- Madison, K.C.; Swartzendruber, D.C.; Wertz, P.W.; Downing, D.T. Presence of intact intercellular lipid lamellae in the upper layers of the stratum corneum. J. Investig. Dermatol. 1987, 88, 714–718. [Google Scholar] [CrossRef] [PubMed]
- Nădăban, A.; Gooris, G.S.; Beddoes, C.M.; Dalgliesh, R.M.; Bouwstra, J.A. Phytosphingosine ceramide mainly localizes in the central layer of the unique lamellar phase of skin lipid model systems. J. Lipid Res. 2022, 63, 100258. [Google Scholar] [CrossRef] [PubMed]
- Beddoes, C.M.; Gooris, G.S.; Barlow, D.J.; Lawrence, M.J.; Dalgliesh, R.M.; Malfois, M.; Demé, B.; Bouwstra, J.A. The importance of ceramide headgroup for lipid localisation in skin lipid models. Biochim. Biophys. Acta Biomembr. 2022, 1864, 183886. [Google Scholar] [CrossRef] [PubMed]
- Bouwstra, J.A.; Ponec, M. The skin barrier in healthy and diseased state. Biochim. Biophys. Acta 2006, 1758, 2080–2095. [Google Scholar] [CrossRef]
- van Smeden, J.; Janssens, M.; Gooris, G.S.; Bouwstra, J.A. The important role of stratum corneum lipids for the cutaneous barrier function. Biochim. Biophys. Acta 2014, 1841, 295–313. [Google Scholar] [CrossRef]
- Schmitt, T.; Neubert, R.H.H. State of the art in stratum corneum research. Part II: Hypothetical stratum corneum lipid matrix models. Skin Pharmacol. Physiol. 2020, 33, 213–230. [Google Scholar] [CrossRef]
- Shamaprasad, P.; Frame, C.O.; Moore, T.C.; Yang, A.; Iacovella, C.R.; Bouwstra, J.A.; Bunge, A.L.; McCabe, C. Using molecular simulation to understand the skin barrier. Prog. Lipid Res. 2022, 88, 101184. [Google Scholar] [CrossRef]
- Elias, P.M.; Wakefield, J.S.; Man, M.Q. Moisturizers versus current and next-generation barrier repair therapy for the management of atopic dermatitis. Skin Pharmacol. Physiol. 2019, 32, 1–7. [Google Scholar] [CrossRef]
- Madison, K.C. Barrier function of the skin: “la raison d’être” of the epidermis. J. Investig. Dermatol. 2003, 121, 231–241. [Google Scholar] [CrossRef]
- Draelos, Z.D. The effect of ceramide-containing skin care products on eczema resolution duration. Cutis 2008, 81, 87–91. [Google Scholar]
- Spada, F.; Barnes, T.M.; Greive, K.A. Skin hydration is significantly increased by a cream formulated to mimic the skin’s own natural moisturizing systems. Clin. Cosmet. Investig. Dermatol. 2018, 11, 491–497. [Google Scholar] [CrossRef]
- Draelos, Z.D.; Baalbaki, N.H.; Raab, S.; Colón, G. The effect of a ceramide-containing product on stratum corneum lipid levels in dry legs. J. Drugs Dermatol. 2020, 19, 372–376. [Google Scholar] [CrossRef]
- Draelos, Z.D.; Hall, S.; Munsick, C. A 14-day controlled study assessing qualitative improvement with 15% lactic acid and ceramides in skin moisturization and desquamation. J. Clin. Aesthet. Dermatol. 2020, 13, E54–E58. Available online: https://pubmed.ncbi.nlm.nih.gov/33178384/ (accessed on 19 May 2023).
- Spada, F.; Harrison, I.P.; Barnes, T.M.; Greive, K.A.; Daniels, D.; Townley, J.P.; Mostafa, N.; Fong, A.T.; Tong, P.H.; Shumack, S. A daily regimen of a ceramide-dominant moisturizing cream and cleanser restores the skin permeability barrier in adults with moderate eczema: A randomized trial. Dermatol. Ther. 2021, 34, e14970. [Google Scholar] [CrossRef]
- Koppes, S.A.; Charles, F.; Lammers, L.; Frings-Dresen, M.; Kezic, S.; Rustemeyer, T. Efficacy of a cream containing ceramides and magnesium in the treatment of mild to moderate atopic dermatitis: A randomized, double-blind, emollient- and hydrocortisone-controlled trial. Acta Derm. Venereol. 2016, 96, 948–953. [Google Scholar] [CrossRef] [PubMed]
- Cha, H.J.; He, C.; Zhao, H.; Dong, Y.; An, I.S.; An, S. Intercellular and intracellular functions of ceramides and their metabolites in skin (Review). Int. J. Mol. Med. 2016, 38, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Ge, F.; Sun, K.; Hu, Z.; Dong, X. Role of omega-hydroxy ceramides in epidermis: Biosynthesis, barrier integrity and analyzing method. Int. J. Mol. Sci. 2023, 24, 5035. [Google Scholar] [CrossRef] [PubMed]
- Kendall, A.C.; Kiezel-Tsugunova, M.; Brownbridge, L.C.; Harwood, J.L.; Nicolaou, A. Lipid functions in skin: Differential effects of n-3 polyunsaturated fatty acids on cutaneous ceramides, in a human skin organ culture model. Biochim. Biophys. Acta Biomembr. 2017, 1859, 1679–1689. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.J.; Rawlings, A.V. The chemistry, function and (patho)physiology of stratum corneum barrier ceramides. Int. J. Cosmet. Sci. 2017, 39, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Fujii, M. The pathogenic and therapeutic implications of ceramide abnormalities in atopic dermatitis. Cells 2021, 10, 2386. [Google Scholar] [CrossRef] [PubMed]
- Zwara, A.; Wertheim-Tysarowska, K.; Mika, A. Alterations of ultra long-chain fatty acids in hereditary skin diseases—Review article. Front. Med. 2021, 8, 730855. [Google Scholar] [CrossRef]
- Farwick, M.; Lersch, P.; Santonnat, B. Developments in ceramide identification, synthesis, function and nomenclature. Cosmet. Toilet. Mag. 2009, 124, 63–72. [Google Scholar]
- Kessner, D.; Brezesinski, G.; Funari, S.S.; Dobner, B.; Neubert, R.H.H. Impact of the long chain omega-acylceramides on the stratum corneum lipid nanostructure. Part 1: Thermotropic phase behaviour of CER[EOS] and CER[EOP] studied using X-ray powder diffraction and FT-Raman spectroscopy. Chem. Phys. Lipids 2010, 163, 42–50. [Google Scholar] [CrossRef]
- van Smeden, J.; Hoppel, L.; van der Heijden, R.; Hankemeier, T.; Vreeken, R.J.; Bouwstra, J.A. LC/MS analysis of stratum corneum lipids: Ceramide profiling and discovery. J. Lipid Res. 2011, 52, 1211–1221. [Google Scholar] [CrossRef]
- Kawana, M.; Miyamoto, M.; Ohno, Y.; Kihara, A. Comparative profiling and comprehensive quantification of stratum corneum ceramides in humans and mice by LC/MS/MS. J. Lipid Res. 2020, 61, 884–895. [Google Scholar] [CrossRef]
- Masukawa, Y.; Narita, H.; Shimizu, E.; Kondo, N.; Sugai, Y.; Oba, T.; Homma, R.; Ishikawa, J.; Takagi, Y.; Kitahara, T.; et al. Characterization of overall ceramide species in human stratum corneum. J. Lipid Res. 2008, 49, 1466–1476. [Google Scholar] [CrossRef] [PubMed]
- Boncheva, M. The physical chemistry of the stratum corneum lipids. Int. J. Cosmet. Sci. 2014, 36, 505–515. [Google Scholar] [CrossRef]
- Bouwstra, J.A.; Helder, R.W.J.; El Ghalbzouri, A. Human skin equivalents: Impaired barrier function in relation to the lipid and protein properties of the stratum corneum. Adv. Drug Deliv. Rev. 2021, 175, 113802. [Google Scholar] [CrossRef]
- Norlén, L.; Nicander, I.; Lundsjö, A.; Cronholm, T.; Forslind, B. A new HPLC-based method for the quantitative analysis of inner stratum corneum lipids with special reference to the free fatty acid fraction. Arch. Dermatol. Res. 1998, 290, 508–516. [Google Scholar] [CrossRef]
- Novotný, J.; Janůsová, B.; Novotný, M.; Hrabálek, A.; Vávrová, K. Short-chain ceramides decrease skin barrier properties. Skin Pharmacol. Physiol. 2009, 22, 22–30. [Google Scholar] [CrossRef]
- Pascher, I. Molecular arrangements in sphingolipids. Conformation and hydrogen bonding of ceramide and their implication on membrane stability and permeability. Biochim. Biophys. Acta 1976, 455, 433–451. [Google Scholar] [CrossRef]
- Snyder, P.W.; Mecinovic, J.; Moustakas, D.T.; Thomas, S.W., 3rd; Harder, M.; Mack, E.T.; Lockett, M.R.; Héroux, A.; Sherman, W.; Whitesides, G.M. Mechanism of the hydrophobic effect in the biomolecular recognition of arylsulfonamides by carbonic anhydrase. Proc. Natl. Acad. Sci. USA 2011, 108, 17889–17894. [Google Scholar] [CrossRef] [PubMed]
- Damien, F.; Boncheva, M. The extent of orthorhombic lipid phases in the stratum corneum determines the barrier efficiency of human skin in vivo. J. Investig. Dermatol. 2010, 130, 611–614. [Google Scholar] [CrossRef] [PubMed]
- Roussel, L.; Atrux-Tallau, N.; Pirot, F. Glycerol as a skin barrier influencing humectant. In Treatment of Dry Skin Syndrome: The Art and Science of Moisturizers, 1st ed.; Lodén, M., Maibach, H.I., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 473–480. [Google Scholar] [CrossRef]
- Garmy, N.; Taïeb, N.; Yahi, N.; Fantini, J. Interaction of cholesterol with sphingosine: Physicochemical characterization and impact on intestinal absorption. J. Lipid Res. 2005, 46, 36–45. [Google Scholar] [CrossRef]
- Shamaprasad, P.; Moore, T.C.; Xia, D.; Iacovella, C.R.; Bunge, A.L.; McCabe, C. Multiscale simulation of ternary stratum corneum lipid mixtures: Effects of cholesterol composition. Langmuir 2022, 38, 7496–7511. [Google Scholar] [CrossRef]
- Macheleidt, O.; Sandhoff, K.; Kaiser, H.W. Deficiency of epidermal protein-bound Ω-hydroxyceramides in atopic dermatitis. J. Investig. Dermatol. 2002, 119, 166–173. [Google Scholar] [CrossRef]
- Ishikawa, J.; Narita, H.; Kondo, N.; Hotta, M.; Takagi, Y.; Masukawa, Y.; Kitahara, T.; Takema, Y.; Koyano, S.; Yamazaki, S.; et al. Changes in the ceramide profile of atopic dermatitis patients. J. Investig. Dermatol. 2010, 130, 2511–2514. [Google Scholar] [CrossRef]
- de Jager, M.; Gooris, G.; Ponec, M.; Bouwstra, J. Acylceramide head group architecture affects lipid organization in synthetic ceramide mixtures. J. Investig. Dermatol. 2004, 123, 911–916. [Google Scholar] [CrossRef]
- Rawlings, A.V.; Lane, M.E. Letter to the Editor regarding ‘An investigation of the skin barrier restoring effects of a cream and lotion containing ceramides in a multi-vesicular emulsion in people with dry, eczema-prone skin: The RESTORE study phase 1’. Dermatol. Ther. 2021, 11, 2245–2248. [Google Scholar] [CrossRef]
- Lee, J.Y.; Jeon, S.; Han, S.; Liu, K.H.; Cho, Y.; Kim, K.P. Positive correlation of triacylglycerols with increased chain length and unsaturation with ω-O-acylceramide and ceramide-NP as well as acidic pH in the skin surface of healthy Korean adults. Metabolites 2022, 13, 31. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.C.; Chang, T.M. Ceramide 1 and ceramide 3 act synergistically on skin hydration and the transepidermal water loss of sodium lauryl sulfate-irritated skin. Int. J. Dermatol. 2008, 47, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Yokose, U.; Ishikawa, J.; Morokuma, Y.; Naoe, A.; Inoue, Y.; Yasuda, Y.; Tsujimura, H.; Fujimura, T.; Murase, T.; Hatamochi, A. The ceramide [NP]/[NS] ratio in the stratum corneum is a potential marker for skin properties and epidermal differentiation. BMC Dermatol. 2020, 20, 6. [Google Scholar] [CrossRef] [PubMed]
- Jungersted, J.M.; Scheer, H.; Mempel, M.; Baurecht, H.; Cifuentes, L.; Høgh, J.K.; Hellgren, L.I.; Jemec, G.B.E.; Agner, T.; Weidinger, S. Stratum corneum lipids, skin barrier function and filaggrin mutations in patients with atopic eczema. Allergy 2010, 65, 911–918. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.H.; Kim, E.J.; Lee, C.H.; Park, G.H.; Yoo, K.M.; Nam, S.J.; Shin, K.O.; Park, K.; Choi, E.H. A Lipid mixture enriched by ceramide NP with fatty acids of diverse chain lengths contributes to restore the skin barrier function impaired by topical corticosteroid. Skin Pharmacol. Physiol. 2022, 35, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Uche, L.E.; Gooris, G.S.; Beddoes, C.M.; Bouwstra, J.A. New insight into phase behavior and permeability of skin lipid models based on sphingosine and phytosphingosine ceramides. Biochim. Biophys. Acta Biomembr. 2019, 1861, 1317–1328. [Google Scholar] [CrossRef]
- Danby, S.G.; Andrew, P.V.; Kay, L.J.; Pinnock, A.; Chittock, J.; Brown, K.; Williams, S.F.; Cork, M.J. Enhancement of stratum corneum lipid structure improves skin barrier function and protects against irritation in adults with dry, eczema-prone skin. Br. J. Dermatol. 2022, 186, 875–886. [Google Scholar] [CrossRef]
- Eichenfield, L.F.; Tom, W.L.; Chamlin, S.L.; Feldman, S.R.; Hanifin, J.M.; Simpson, E.L.; Berger, T.G.; Bergman, J.N.; Cohen, D.E.; Cooper, K.D.; et al. Guidelines of care for the management of atopic dermatitis. Part 1: Diagnosis and assessment of atopic dermatitis. J. Am. Acad. Dermatol. 2014, 70, 338–351. [Google Scholar] [CrossRef]
- Nutten, S. Atopic dermatitis: Global epidemiology and risk factors. Ann. Nutr. Metab. 2015, 66, 8–16. [Google Scholar] [CrossRef]
- Bieber, T. Atopic dermatitis. Ann. Dermatol. 2010, 22, 125–137. [Google Scholar] [CrossRef]
- Proksch, E.; Fölster-Holst, R.; Bräutigam, M.; Sepehrmanesh, M.; Pfeiffer, S.; Jensen, J.M. Role of the epidermal barrier in atopic dermatitis. J. Dtsch. Dermatol. Ges. 2009, 7, 899–910. [Google Scholar] [CrossRef]
- Sugarman, J.L.; Parish, L.C. Efficacy of a lipid-based barrier repair formulation in moderate-to-severe pediatric atopic dermatitis. J. Drugs Dermatol. 2009, 8, 1106–1111. Available online: https://pubmed.ncbi.nlm.nih.gov/20027938/ (accessed on 19 May 2023). [PubMed]
- Lodén, M. The increase in skin hydration after application of emollients with different amounts of lipids. Acta Derm. Venereol. 1992, 72, 327–330. Available online: https://pubmed.ncbi.nlm.nih.gov/1361276/ (accessed on 19 May 2023). [CrossRef] [PubMed]
- Leung, D.Y.; Boguniewicz, M.; Howell, M.D.; Nomura, I.; Hamid, Q.A. New insights into atopic dermatitis. J. Clin. Investig. 2004, 113, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.N.; Irvine, A.D.; Terron-Kwiatkowski, A.; Zhao, Y.; Liao, H.; Lee, S.P.; Goudie, D.R.; Sandilands, A.; Campbell, L.A.; Smith, F.J.D.; et al. Common loss-of-function variants of the epidermal barrier protein filaggrin are a major predisposing factor for atopic dermatitis. Nat. Genet. 2006, 38, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.P.; Zhao, M.T.; Jia, Z.X.; Zhang, J.L.; Jiao, L.; Ma, L. Skin ceramide profile in children with atopic dermatitis. Dermatitis 2018, 29, 219–222. [Google Scholar] [CrossRef]
- Imokawa, G.; Abe, A.; Jin, K.; Higaki, Y.; Kawashima, M.; Hidano, A. Decreased level of ceramides in stratum corneum of atopic dermatitis: An etiologic factor in atopic dry skin? J. Investig. Dermatol. 1991, 96, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Sator, P.G.; Schmidt, J.B.; Hönigsmann, H. Comparison of epidermal hydration and skin surface lipids in healthy individuals and in patients with atopic dermatitis. J. Am. Acad. Dermatol. 2003, 48, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Bleck, O.; Abeck, D.; Ring, J.; Hoppe, U.; Vietzke, J.P.; Wolber, R.; Brandt, O.; Schreiner, V. Two ceramide subfractions detectable in Cer(AS) position by HPTLC in skin surface lipids of non-lesional skin of atopic eczema. J. Investig. Dermatol. 1999, 113, 894–900. [Google Scholar] [CrossRef] [PubMed]
- Di Nardo, A.; Wertz, P.; Giannetti, A.; Seidenari, S. Ceramide and cholesterol composition of the skin of patients with atopic dermatitis. Acta Derm. Venereol. 1998, 78, 27–30. [Google Scholar] [CrossRef]
- Yamamoto, A.; Serizawa, S.; Ito, M.; Sato, Y. Stratum corneum lipid abnormalities in atopic dermatitis. Arch. Dermatol. Res. 1991, 283, 219–223. [Google Scholar] [CrossRef]
- Janssens, M.; van Smeden, J.; Gooris, G.S.; Bras, W.; Portale, G.; Caspers, P.J.; Vreeken, R.J.; Hankemeier, T.; Kezic, S.; Wolterbeek, R.; et al. Increase in short-chain ceramides correlates with an altered lipid organization and decreased barrier function in atopic eczema patients. J. Lipid Res. 2012, 53, 2755–2766. [Google Scholar] [CrossRef] [PubMed]
- Vávrová, K.; Kováčik, A.; Opálka, L. Ceramides in the skin barrier. Euro. Pharm. J. 2017, 64, 28–35. [Google Scholar] [CrossRef]
- Snyder, R.G.; Liang, G.L.; Strauss, H.L.; Mendelsohn, R. IR spectroscopic study of the structure and phase behavior of long-chain diacylphosphatidylcholines in the gel state. Biophys. J. 1996, 71, 3186–3198. [Google Scholar] [CrossRef]
- Eichenfield, L.F.; Tom, W.L.; Berger, T.G.; Krol, A.; Paller, A.S.; Schwarzenberger, K.; Bergman, J.N.; Chamlin, S.L.; Cohen, D.E.; Cooper, K.D.; et al. Guidelines of care for the management of atopic dermatitis: Section 2. Management and treatment of atopic dermatitis with topical therapies. J. Am. Acad. Dermatol. 2014, 71, 116–132. [Google Scholar] [CrossRef]
- Schachner, L.A.; Andriessen, A.; Benjamin, L.; Bree, A.F.; Lechman, P.A.; Pinera-Llano, A.A.; Kircik, L. A consensus about the importance of ceramide containing skincare for normal and sensitive skin conditions in neonates and infants. J. Drugs Dermatol. 2020, 19, 769–776. [Google Scholar] [CrossRef]
- Correa, M.C.M.; Nebus, J. Management of patients with atopic dermatitis: The role of emollient therapy. Dermatol. Res. Pract. 2012, 2012, 836931. [Google Scholar] [CrossRef]
- Nicol, N.H.; Rippke, F.; Weber, T.M.; Hebert, A.A. Daily moisturization for atopic dermatitis: Importance, recommendations, and moisturizer choices. J. Nurse Pract. 2021, 17, 920–925. Available online: https://www.researchgate.net/publication/353432758_Daily_Moisturization_for_Atopic_Dermatitis_Importance_Recommendations_and_Moisturizer_Choices (accessed on 19 May 2023). [CrossRef]
- Sirikudta, W.; Kulthanan, K.; Varothai, S.; Nuchkull, P. Moisturizers for patients with atopic dermatitis: An overview. J. Allergy Ther. 2013, 4, 143. [Google Scholar] [CrossRef]
- Mawazi, S.M.; Ann, J.; Othman, N.; Khan, J.; Alolayan, S.O.; Al thagfan, S.S.; Kaleemullah, M. A review of moisturizers: History, preparation, characterization and applications. Cosmetics 2022, 9, 61. [Google Scholar] [CrossRef]
- Kang, S.Y.; Um, J.Y.; Chung, B.Y.; Lee, S.Y.; Park, J.S.; Kim, J.C.; Park, C.W.; Kim, H.O. Moisturizer in patients with inflammatory skin diseases. Medicina 2022, 58, 888. [Google Scholar] [CrossRef] [PubMed]
- Harrison, I.P.; Spada, F. Breaking the itch-scratch cycle: Topical options for the management of chronic cutaneous itch in atopic dermatitis. Medicines 2019, 6, 76. [Google Scholar] [CrossRef] [PubMed]
- Danby, S.G.; Andrew, P.V.; Brown, K.; Chittock, J.; Kay, L.J.; Cork, M.J. An investigation of the skin barrier restoring effects of a cream and lotion containing ceramides in a multi-vesicular emulsion in people with dry, eczema-prone, skin: The RESTORE Study Phase 1. Dermatol. Ther. 2020, 10, 1031–1041. [Google Scholar] [CrossRef]
- Kono, T.; Miyachi, Y.; Kawashima, M. Clinical significance of the water retention and barrier function-improving capabilities of ceramide-containing formulations: A qualitative review. J. Dermatol. 2021, 48, 1807–1816. [Google Scholar] [CrossRef]
- Tanno, O.; Ota, Y.; Kitamura, N.; Katsube, T.; Inoue, S. Nicotinamide increases biosynthesis of ceramides as well as other stratum corneum lipids to improve the epidermal permeability barrier. Br. J. Dermatol. 2000, 143, 524–531. [Google Scholar] [CrossRef]
- Soma, Y.; Kashima, M.; Imaizumi, A.; Takahama, H.; Kawakami, T.; Mizoguchi, M. Moisturizing effects of topical nicotinamide on atopic dry skin. Int. J. Dermatol. 2005, 44, 197–202. [Google Scholar] [CrossRef]
- Berson, D.S.; Osborne, R.; Oblong, J.E.; Hakozaki, T.; Johnson, M.B.; Bissett, D.L. Niacinamide: A topical vitamin with wide-ranging skin appearance benefits. In Cosmeceuticals and Cosmetic Practice, 1st ed.; Farris, P.K., Ed.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2013; pp. 103–112. [Google Scholar]
- Christman, J.C.; Fix, D.K.; Lucus, S.C.; Watson, D.; Desmier, E.; Wilkerson, R.J.; Fixler, C. Two randomized, controlled, comparative studies of the stratum corneum integrity benefits of two cosmetic niacinamide/glycerin body moisturizers vs. conventional body moisturizers. J. Drugs Dermatol. 2012, 11, 22–29. [Google Scholar]
- Rawlings, A.V.; Davies, A.; Carlomusto, M.; Pillai, S.; Zhang, K.; Kosturko, R.; Verdejo, P.; Feinberg, C.; Nguyen, L.; Chandar, P. Effect of lactic acid isomers on keratinocyte ceramide synthesis, stratum corneum lipid levels and stratum corneum barrier function. Arch. Dermatol. Res. 1996, 288, 383–390. [Google Scholar] [CrossRef]
- Dahl, M.V.; Dahl, A.C. 12% lactate lotion for the treatment of xerosis. A double-blind clinical evaluation. Arch. Dermatol. 1983, 119, 27–30. Available online: https://pubmed.ncbi.nlm.nih.gov/6336928/ (accessed on 19 May 2023). [CrossRef] [PubMed]
- Hebert, A.A.; Rippke, F.; Weber, T.M.; Nicol, N.H. Efficacy of nonprescription moisturizers for atopic dermatitis: An updated review of clinical evidence. Am. J. Clin. Dermatol. 2020, 21, 641–655. [Google Scholar] [CrossRef]
- Clarke, C.J.; Wu, B.X.; Hannun, Y.A. The neutral sphingomyelinase family: Identifying biochemical connections. Advn. Enzyme Reg. 2011, 51, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Polefka, T.G.; Bianchini, R.J.; Shapiro, S. Interaction of mineral salts with the skin: A literature survey. Int. J. Cosmet. Sci. 2012, 34, 416–423. [Google Scholar] [CrossRef]
- Oh, M.J.; Cho, Y.H.; Cha, S.Y.; Lee, E.O.; Kim, J.W.; Kim, S.K.; Park, C.S. Novel phytoceramides containing fatty acids of diverse chain lengths are better than a single C18-ceramide N-stearoyl phytosphingosine to improve the physiological properties of human stratum corneum. Clin. Cosmet. Investig. Dermatol. 2017, 10, 363–371. [Google Scholar] [CrossRef]
- Both, D.M.; Goodtzova, K.; Yarosh, D.B.; Brown, D.A. Liposome-encapsulated ursolic acid increases ceramides and collagen in human skin cells. Arch. Dermatol. Res. 2002, 293, 569–575. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mijaljica, D.; Townley, J.P.; Hondros, A.; Hewson, C.; Harrison, I.P.; Spada, F. Considering Phytosphingosine-Based Ceramide Formulations for Atopic Skin Care. Dermato 2024, 4, 5-22. https://doi.org/10.3390/dermato4010002
Mijaljica D, Townley JP, Hondros A, Hewson C, Harrison IP, Spada F. Considering Phytosphingosine-Based Ceramide Formulations for Atopic Skin Care. Dermato. 2024; 4(1):5-22. https://doi.org/10.3390/dermato4010002
Chicago/Turabian StyleMijaljica, Dalibor, Joshua P. Townley, Angelina Hondros, Caroline Hewson, Ian P. Harrison, and Fabrizio Spada. 2024. "Considering Phytosphingosine-Based Ceramide Formulations for Atopic Skin Care" Dermato 4, no. 1: 5-22. https://doi.org/10.3390/dermato4010002