
Ecotonic Communities of Diatoms in the Southeastern Part of the Kamchatka Peninsula




Abstract
:1. Introduction
2. Materials and Methods
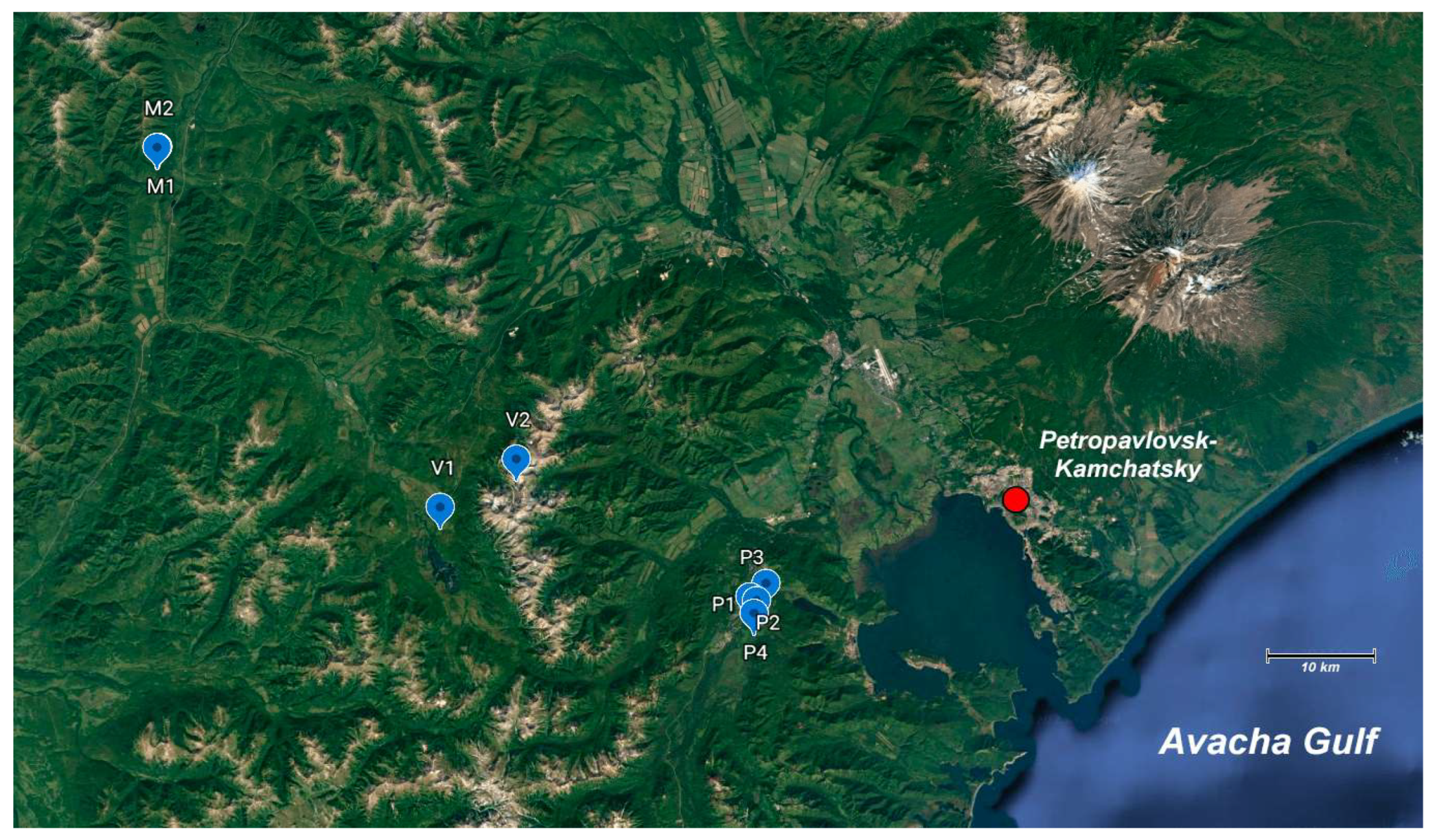
2.1. Studied Area

2.2. Sample Collection and Data Analysis
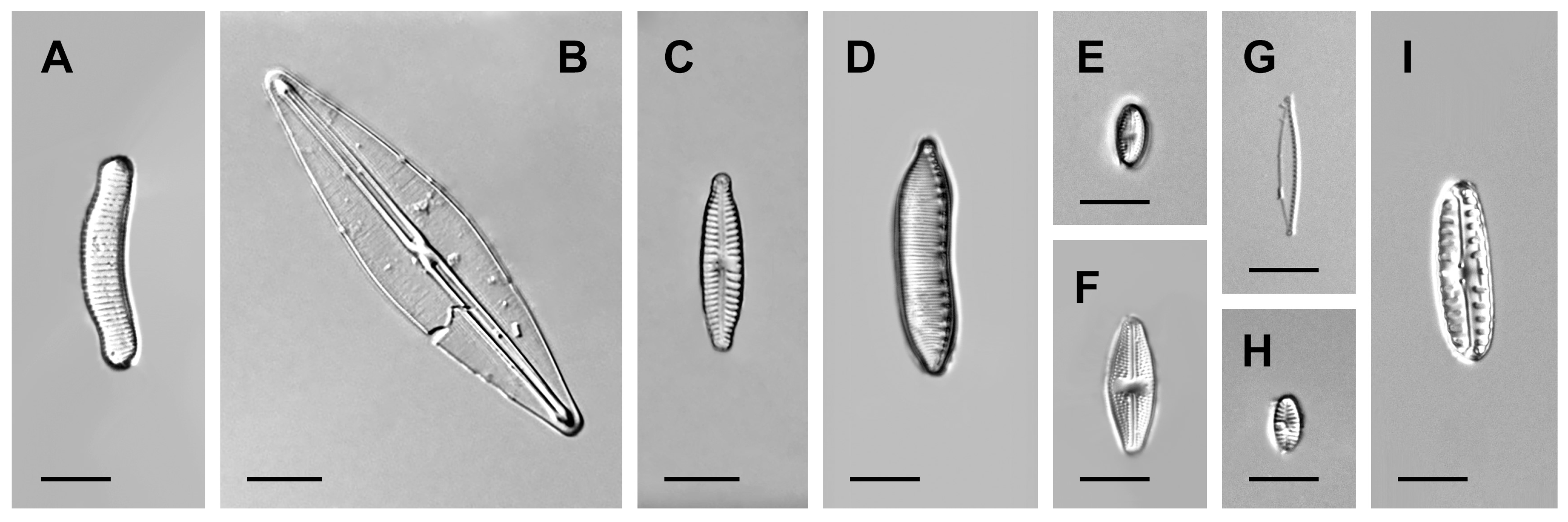
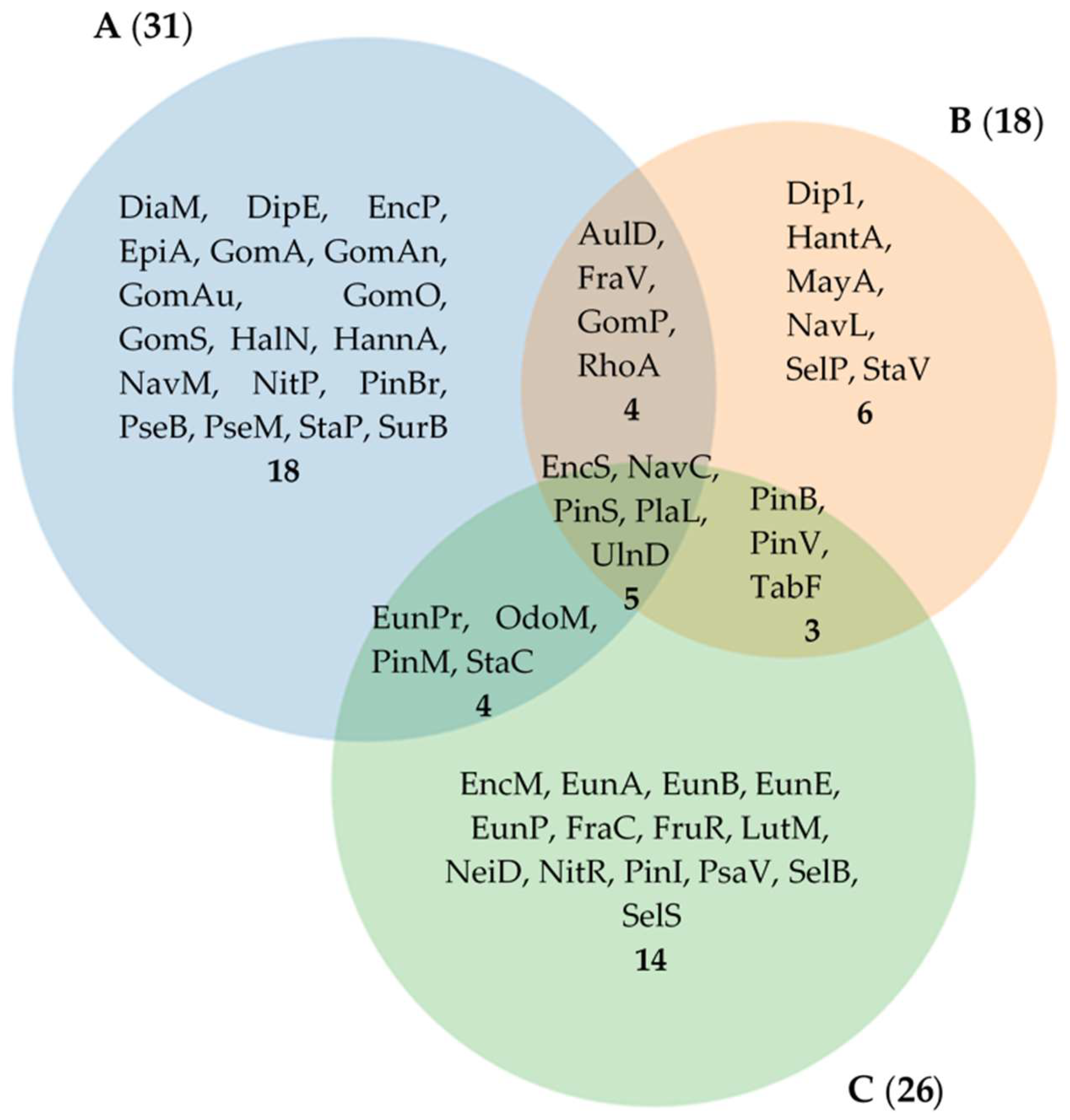
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bezais, E.K. Conditions of soil formation in Kamchatka. Mater. Study Russ. Soils 1911, 20, 89–148. (In Russian) [Google Scholar]
- Liverovsky, Y.A. Soils of Kamchatka; Nauka: Moscow-Leningrad, Russia, 1940; pp. 127–156. (In Russian) [Google Scholar]
- Liverovsky, Y.A. Soils of the Plains of the Kamchatka Peninsula; Nauka: Moscow-Leningrad, Russia, 1959; p. 126. (In Russian) [Google Scholar]
- Zonn, S.V.; Karpachevsky, L.O.; Stefin, V.V. Forest Soils of Kamchatka; Publishing House of the Academy of Sciences of the USSR: Moscow, Russia, 1963; pp. 182–198. (In Russian) [Google Scholar]
- Sokolov, I.A. Peculiarities of geochemistry of landscapes from Kamchatka in connection with modern volcanic activity. In Geochemistry of Landscapes; Nauka: Moscow, Russia, 1967; pp. 72–95. (In Russian) [Google Scholar]
- Sokolov, I.A. Volcanic Activity and Soil Generation; Nauka: Moscow, Russia, 1973; p. 224. (In Russian) [Google Scholar]
- Karpachevsky, L.O.; Alyabyina, I.O.; Zakharikhina, L.V.; Makeev, A.O.; Merechek, M.S.; Radyukin, A.Y.U.; Shoba, S.A. Kamchatka Soils; GEOS: Moscow, Russia, 2009; p. 224. (In Russian) [Google Scholar]
- Afanasyeva, T.V.; Vasilenko, V.I.; Tereshina, T.V.; Sheremet, B.V. Soils of the USSR; Mysl: Moscow, Russia, 1979; p. 380. (In Russian) [Google Scholar]
- Kondratyuk, V.I. Climate of Kamchatka; Hydrometeoizdat: Moscow, Russia, 1974; p. 204. (In Russian) [Google Scholar]
- Manko, Y.I.; Sidelnikov, A.N. Effect of Volcanism on Vegetation; DVO AN SSSR: Vladivostok, Russia, 1989; p. 188. (In Russian) [Google Scholar]
- Duggen, S.; Portnyagin, M.; Baker, J.; Ulfbeck, D.; Hoernle, K.; Garbe-Schönberg, D.; Grassineau, N. Drastic shift in lava geochemistry in the volcanic-front to rear-arc region of the Southern Kamchatkan subduction zone: Evidence for the transition from slab surface dehydration to sediment melting. Geochim. Cosmochim. Acta 2007, 71, 452–480. [Google Scholar] [CrossRef]
- Chashchin, A.A.; Martynov, Y.A.; Perepelov, A.B.; Ekimova, N.I.; Vladimirova, T.P. Physical and chemical conditions of the formation and evolution of late pleistocene-holocene magmas of the Gorely and Mutnovsky volcanoes, southern Kamchatka. Russ. J. Pac. Geol. 2011, 5, 348–367. [Google Scholar] [CrossRef]
- Neshataeva, V.Y. Vegetation of the Kamchatka Peninsula; KMK: Moscow, Russia, 2009; p. 537. (In Russian) [Google Scholar]
- Zakharikhina, L.V.; Litvinenko, Y.S. Volcanism and Geochemistry of Soil and Vegetation Cover of Kamchatka. Communication 1. Geochemical features of volcanic surface ashes. Volcanol. Seismol. 2019, 2, 34–44. [Google Scholar]
- Zakharikhina, L.; Litvinenko, Y.S. Volcanism and Geochemistry of Soil and Vegetation Cover of Kamchatka. Communication 2. Specificity of forming the elemental composition of volcanic soil in cold and humid conditions. Volcanol. Seismol. 2019, 3, 25–33. [Google Scholar]
- Koloskov, A.V.; Flerov, G.B.; Perepelov, A.B.; Melekestsev, I.V.; Puzankov, M.Y.; Filosofova, T.M. Evolution stages and petrology of the Kekuknai volcanic massif as reflecting the Magmatismin Backarc Zone of Kuril-Kamchatka Island Arc System. Part 1. Geological position and geochemistry of volcanic rocks. J. Volcanol. Seismol. 2011, 5, 312–334. [Google Scholar] [CrossRef] [Green Version]
- Prytkov, A.S.; Vasilenko, N.F.; Frolov, D.I. Recent geodynamics of the Kuril subduction zone. Russ. J. Pac. Geol. 2017, 11, 19–24. [Google Scholar] [CrossRef]
- Prijdun, V.I.; Karpenko, V.I.; Mikhaylova, E.G. Natural resources of Kamchatka territory and prospects for their rational use. In Proceedings of the IX All-Russian Scientific and Practical Conference “Natural Resources, Their Current State, Protection, Commercial and Technical Use”, Petropavlovsk-Kamchatsky, Russia, 20–22 March 2018. [Google Scholar]
- Sharipova, M.Y.; Dubovik, I.E.; Putenikhin, V.P.; Akhmedyanov, D.I.; Gabidullina, G.F.; Gulamanova, G.A.; Teltsova, L.Z.; Turyanova, R.R. Algological assessment of oil pollution in ecotonic ecosystems. IOP Conf. Ser. Earth Environ. Sci. 2020, 548, 062079. [Google Scholar] [CrossRef]
- Zaletaev, V.S. Ecotones in the Biosphere; Publishing House of the Russian Academy of Agricultural Sciences: Moscow, Russia, 1997; pp. 1–329. (In Russian) [Google Scholar]
- Hansen, A.J.; diCastri, F.; Naiman, R.J. Ecotones: What and why? Biol. Intl. 1988, 17, 9–46. [Google Scholar]
- Fleituch, T.; Starzecka, A.; Bednarz, T. Spatial trends in sediment structure, bacteria, and periphyton communities across a freshwater ecotone. Hydrobiologia 2001, 464, 165–174. [Google Scholar] [CrossRef]
- Izaguirre, I.; O’Farrell, I.; Tell, G. Variation in phytoplankton composition and limnological features in a water-water ecotone of the lower Paraná Basin (Argentina). Freshwat. Biol. 2001, 46, 63–74. [Google Scholar]
- Komulainen, S.F. Structural and functional organization of river periphyton as an ecotone community. In Proceedings of the 8th Congress of the Hydrobiological Society of the Russian Academy of Science, Kaliningrad, Russia, 16–23 September 2001; pp. 182–183. (In Russian). [Google Scholar]
- Sharipova, M.Y. Algae of Ecotone Communities; RIO BashGU: Ufa, Russia, 2006; pp. 1–182. (In Russian) [Google Scholar]
- Stenger-Kovács, C.; Buczko, K.; Hajnal, E.; Padisak, J. Epiphytic, littoral diatoms as bioindicators of shallow lake trophic status: Trophic Diatom Index for Lakes (TDIL) developed in Hungary. Hydrobiologia 2007, 589, 141–154. [Google Scholar] [CrossRef]
- Lobo, E.A.; Heinrich, C.G.; Schuch, M.; Wetzel, C.E.; Ector, L. Diatoms as Bioindicators in Rivers. In River Algae; Necchi, O., Jr., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 245–271. [Google Scholar] [CrossRef]
- Wanner, M.; Birkhofer, K.; Fischer, T.; Shimizu, M.; Shimano, S.; Puppe, D. Soil testate amoebae and diatoms as bioindicators of an old heavy metal contaminated floodplain in Japan. Microb. Ecol. 2020, 79, 123–133. [Google Scholar] [CrossRef]
- Genkal, S.I.; Lepskaya, E.V. Centric diatom algae of Volcanic Verkhneavachinsk Lakes (Kamchatka). Inland Water Biol. 2014, 7, 1–9. [Google Scholar] [CrossRef]
- Genkal, S.I.; Lepskaya, E.V. Materials to the Flora of Bacillariophyta of Lake Kronotskoye (the Kamchatka Peninsula, Russia). Int. J. Algae 2015, 17, 14–22. [Google Scholar] [CrossRef]
- Genkal, S.I.; Lepskaya, E.V. Diatoms (Bacillariophyta) of the Tolmachev reservoir (Kamchatka, Russia). Bot. Pac. A J. Plant Sci. Conserv. 2023; in press. [Google Scholar] [CrossRef]
- Solovieva, N.; Klimaschewski, A.; Self, A.E.; Jones, V.J.; Andren, E.; Andreev, A.A.; Hammarlund, D.; Lepskaya, E.V.; Nazarova, L. Holocene environmental history of a small coastal lake from north-eastern Kamchatka Peninsula. Glob. Planet. Chang. 2015, 134, 55e66. [Google Scholar] [CrossRef] [Green Version]
- Nikulina, T.V.; Kalitina, E.G.; Vakh, E.A.; Kharitonova, N.A. List of diatoms from three hot springs from Kamchatka—Malkinskiye, Nachikinskiye and Verhne-Paratunskiye (Russia). In Freshwater Life; Bogatov, V.V., Ed.; Dalnauka: Vladivostok, Russia, 2016; Volume 2, pp. 108–115. (In Russian) [Google Scholar]
- Nikulina, T.V.; Grishchenko, O.V. Diatom flora of Dachnye Thermal Springs (Kamchatka Peninsula, Russia). In Vladimir Ya. Levanidov’s Biennial Memorial Meetings; FNC of Biodiversity FEB RAS: Vladivostok, Russia, 2017; Volume 7, pp. 185–193. Available online: http://www.ibss.febras.ru/files/00015192.pdf (accessed on 25 December 2022). (In Russian)
- Fazlutdinova, A.I.; Gabidullin, Y.Z.; Allaguvatova, R.Z.; Gaysina, L.A. Diatoms in Kamchatka’s Hot Spring Soils. Diversity 2020, 12, 435. [Google Scholar] [CrossRef]
- Shtina, E.A.; Andreyeva, V.M.; Kuzyakina, T.I. Algae settlement of volcanic substrates. Bot. Zhurnal 1992, 8, 33–42. [Google Scholar]
- Fazlutdinova, A.I.; Gabidullin, Y.; Allaguvatova, R.; Gaysina, L. Diatoms in Volcanic Soils of Mutnovsky and Gorely Volcanoes (Kamchatka Peninsula, Russia). Microorganisms 2021, 9, 1851. [Google Scholar] [CrossRef]
- Litvinenko, Y.S.; Zakharikhina, L.V. Zoning and geochemical characterization of volcanic soils on Kamchatka. Geochem. Int. 2009, 47, 463–475. [Google Scholar] [CrossRef]
- Dumont, H.J.; Haritonov, A.Y.; Kosterin, O.E.; Malikova, E.I.; Popova, O.G. A review of the Odonata of Kamchatka Peninsula, Russia. Odonatologica 2005, 34, 131–153. [Google Scholar]
- Khomutov, S.Y. Magnetic observations at Geophysical Observatory Paratunka IKIR FEB RAS: Tasks, possibilities and future prospects. In Proceedings of the Solar-Terrestrial Relations and Physics of Earthquake Precursors, Paratunka, Russia, 25–29 September 2017; p. 02002. [Google Scholar] [CrossRef]
- Hydrometeorological data, Leningrad, Russia. In Surface Water Resources of the USSR; Hydrometeoisdat: Kamchatka, Russia, 1973; Volume 20, p. 368. (In Russian)
- Barragán, C.; Wetzel, C.E.; Ector, L. A standard method for the routine sampling of terrestrial diatom communities for soil quality assessment. J. Appl. Phycol. 2018, 30, 1095–1113. [Google Scholar] [CrossRef]
- Acker, F.; Russell, B.; Morales, E. Preparation of Diatom Slides Using NaphraxTM Mounting Medium; Protocol P-13-49; Academy of Natural Sciences of Philadelphia, Patrick Center for Environmental Research: Philadelphia, PA, USA, 1999; pp. 13–42. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae, Naviculaceae. Süβwasserflora von Mitteleuropa; Band 2/1; Spectrum Academiche Verlag: Berlin, Germany, 1986; p. 876. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae, Teil 2. Epithemiaceae, Bacillariaceae, Surirellaceae. In Süsswasserflora von Mitteleuropa; Pascher, A., Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; VEB Gustav Fisher Verlag: Stuttgart, Germany, 1988; p. 596. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae, Teil 3. Centrales, Fragilariaceae, Eunotiaceae, Achnanthaceae. In Süβwasserflora von Mitteleuropa; Pascher, A., Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; VEB Gustav Fisher Verlag: Stuttgart, Germany, 1991; p. 576. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae, Teil 4. Achnanthaceae, Kritische Erganzungen zu Navicula (Lineolatae) und Gomphonema. In Süsswasserflora von Mitteleuropa; Pascher, A., Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; VEB Gustav Fisher Verlag: Stuttgart, Germany, 1991; p. 436. [Google Scholar]
- Hofmann, G.; Werum, M.; Lange-Bertalot, H. Diatomeen im Süßwasser—Benthos von Mitteleuropa. In Bestimmungsflora Kieselalgen für die ökologische Praxis; Über 700 der häufigsten Arten und ihre Ökologie; Koeltz Scientific Books: Königstein, Germany, 2013; p. 908. [Google Scholar]
- Paula, C.F.; Lowe, R.L.; Johansen, J.R. Teratology in Eunotia taxa in The Great Smoky Mountains National Park and description of Eunotia macroglossa spp. nov. Diatom Res. 2009, 24, 273–290. [Google Scholar]
- Solak, C.; Wojtal, A. Diatoms in springs and streams of Turkmen Mt (Sakarya River Basin) common in Turkish Inland waters. Pol. Bot. J. 2012, 57, 345–425. [Google Scholar]
- Wetzel, C.E.; Ector, L.; Vijver, B.; Compère, P.; Mann, D.G. Morphology, typification and critical analysis of some ecologically important small naviculoid species (Bacillariophyta). Fottea 2015, 15, 203–234. [Google Scholar] [CrossRef] [Green Version]
- Potapova, M.G. Diatoms of Bering Island, Kamchatka, Russia. Nova Hedwig. 2014, 143, 63–102. [Google Scholar]
- Beauger, A.; Wetzel, C.E.; Voldoire, O.; Garreau, A.; Ector, L. Sellaphora labernardierei (Sellaphoraceae, Bacillariophyta), a new epilithic species from French spring and four new combinations within the genus Sellaphora. Phytotaxa 2016, 260, 235–246. [Google Scholar] [CrossRef]
- Kostikov, I.Y.; Romanenko, P.O.; Demchenko, E.M.; Darienko, T.M.; Mikhailyuk, T.I.; Rybchinsky, O.V.; Solonenko, A.M. Algae in Soils of Ukraine: History and Methods of Studies, System, and List of the Algal Flora; Fitosotsiotsentr: Kiev, Ukraine, 2001; p. 300. (In Ukrainian) [Google Scholar]
- Venn, J. On the diagrammatic and mechanical representation of propositions and reasonings. Philos. Mag. J. Sci. 1880, 9, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Hulsen, T.; de Vlieg, J.; Alkema, W. BioVenn—A web application for the comparison and visualization of biological lists using area-proportional Venn diagrams. BMC Genom. 2008, 9, 488. [Google Scholar] [CrossRef] [Green Version]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef] [Green Version]
- Kabirov, R.R.; Safiulina, L.M. Peculiarities of ecology and distribution of unicellular soil alga Eustigmatos magnus (J.B. Petersen) Hibberd in Southern Ural (Russia). Int. J. Algae 2008, 10, 105–116. [Google Scholar]
- Misra, A.A.; Mukherjee, S. Introduction to “Atlas of Structural Geological and Geomorphological Interpretation of Remote Sensing Images”. In Atlas of Structural Geological and Geomorphological Interpretation of Remote Sensing Images; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2022; pp. 1–5. [Google Scholar] [CrossRef]
- Nikulina, T.V.; Kociolek, J.P. Diatoms from hot springs from Kuril and Sakhalin Islands (Far East, Russia). In Cellular Origin, Life in Extreme Habitats and Astrobiology; The DiatomWorld, Part 3; Seckbach, J., Kociolek, J.P., Eds.; Springer: Dordrecht, The Netherlands, 2011; Volume 19, pp. 333–363. [Google Scholar]
- Zaitsev, Y.P. Ecotones of the Black Sea. In Ecotones in the Biosphere; Publishing House of the Russian Academy of Agricultural Sciences: Moscow, Russia, 1997; pp. 240–258. (In Russian) [Google Scholar]
- Foets, J.; Stanek-Tarkowska, J.; Teuling, A.J.; Van de Vijver, B.; Wetzel, C.E.; Pfister, L. Autecology of terrestrial diatoms under anthropic disturbance and across climate zones. Ecol. Indic. 2021, 122, 107248. [Google Scholar] [CrossRef]
- Ilchibaeva, K.V.; Kunsbaeva, D.F.; Allaguvatova, R.Z.; Fazlutdinova, A.I.; Polokhin, O.V.; Sibirina, L.A.; Gontcharov, A.A.; Singh, P.; Gaysina, L.A. Preliminary data about algae and cyanobacteria of volcanic soils on Kuril islands. Theor. Appl. Ecol. 2018, 4, 119–126. [Google Scholar] [CrossRef]
- Lai, G.G.; Beauger, A.; Wetzel, C.E.; Padedda, B.M.; Voldoire, O.; Lugliè, A.; Allain, E.; Ector, L. Diversity, ecology and distribution of benthic diatoms in thermo-mineral springs in Auvergne (France) and Sardinia (Italy). PeerJ 2019, 7, e7238. [Google Scholar] [CrossRef]
- Ishikawa, S.; Kashima, K. Diatoms in Bekanbeushi Wetland, Eastern Hokkaido. Diatom 2009, 25, 106–110. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Site Number | Site Description | Site Name | GPS | The Name of the Area | Sample Description |
|---|---|---|---|---|---|
| 1 | Central part of the Paratunka river valley | P1 | 52°56′59.0″ N 158°16′18.0″ E | Paratunka river | Soil from a swampy area |
| 2 | Abandoned pioneer camp (on the lake shore 500 m from the highway in the forest) | P2 | 52°57′53.0″ N 158°17′07.0″ E | Paratunka river | Soil 0.3 m from the water’s edge |
| 3 | Paratunka village | P3 | 52°57′09.0″ N 158°16′08.0″ E | Paratunka river | Soil from a dried-up swamp |
| 4 | Paratunka river (2 km from the village of Paratunka) | P4 | 52°57′07.0″ N 158°16′10.0″ E | Paratunka river | Soil 0.2 m from the water’s edge |
| 5 | The foot of the Vachkazhets volcano (3 km from the volcano) | V1 | 53°02′54.0″ N 157°52′57.0″ E | Vachkazhets volcano | Soil 10 m from the lake |
| 6 | “Veronica’s Hair” Waterfall | V2 | 53°04′07.0″ N 157°56′28.0″ E | Vachkazhets volcano | Soil at the base of the waterfall bowl |
| 7 | Village Malki (the bank of the Bystraya river) | M1 | 53°19′43.0″ N 157°27′55.0″ E | Bystraya river | Soil 0.3 m from the river bank |
| 8 | Village Malki (the bank of the Bystraya river) | M2 | 53°19′43.0″ N 157°27′55.0″ E | Bystraya river | Soil 15 m from the river bank |
| Taxon | A | B | C | Habitat * | Salinity Tolerance * | pH * | Distribution * | Typical Environment * | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P1 | P2 | P3 | P4 | V1 | V2 | M1 | M2 | ||||||
| Aulacoseira distans (Ehrenberg) Simonsen | 8 | 4 | 4 | 2 | P B | i | ac | b | Marine, freshwater | ||||
| Diatoma moniliformis (Kützing) D.M. Williams | 6 | 5 | P B | hl | al | c | Marine, freshwater | ||||||
| Diploneis elliptica (Kützing) Cleve | 2 | 6 | 1 | 2 | B | i | al | c | Freshwater | ||||
| Diploneis sp.1 | 1 | B | i | al | c | Freshwater | |||||||
| Encyonema minutum (Hilse) D.G. Mann | 4 | 4 | B Ep | i | al | c | Marine, freshwater | ||||||
| Encyonema perpusillum (A. Cleve) D.G. Mann | 5 | 7 | B Ep | hb | ac | c | Freshwater | ||||||
| Encyonema silesiacum (Bleisch) D.G. Mann | 3 | 5 | 4 | 3 | 2 | 3 | 1 | B Ep | i | al | b | Freshwater, terrestrial | |
| Epithemia adnata (Kützing) Brébisson | 10 | 12 | 8 | Ep | i | al | c | Freshwater, terrestrial | |||||
| Eunotia arcus var. fallax Hustedt | 4 | Ep | hb | i | c | Freshwater | |||||||
| Eunotia bilunaris (Ehrenberg) Schaarschmidt | 6 | 8 | L Ep | hb | ac | c | Freshwater | ||||||
| Eunotia exigua (Brébisson ex Kützing) Rabenhorst | 15 | 15 | L Ep | hb | ac | c | Freshwater | ||||||
| Eunotia paratridentula Lange-Bertalot and Kulikovskiy | 2 | 4 | L Ep | hb | ac | c | Freshwater | ||||||
| Eunotia praerupta Ehrenberg | 12 | 5 | 3 | 8 | 8 | 10 | L Ep | hb | ac | c | Freshwater | ||
| Fragilaria capucina Desmazières | 2 | 1 | L P | i | al | c | Marine, freshwater | ||||||
| Fragilariforma virescens (Ralfs) D.M. Williams and Round | 15 | 15 | 15 | 15 | 15 | 15 | L Ep | i | i | aa | Freshwater | ||
| Frustulia rhomboides (Ehrenberg) De Toni | 3 | B | hb | ac | aa | Freshwater, terrestrial | |||||||
| Gomphonema acuminatum Ehrenberg | 13 | 10 | 6 | 4 | Ep | i | i | c | Freshwater | ||||
| Gomphonema angustatum (Kützing) Rabenhorst | 10 | 8 | Ep | i | al | c | Freshwater | ||||||
| Gomphonema augur Ehrenberg | 1 | 1 | Ep | i | i | c | Freshwater | ||||||
| Gomphonema olivaceum (Hornemann) Ehrenberg | 7 | 5 | 4 | Ep | i | al | c | Freshwater | |||||
| Gomphonema parvulum (Kützing) Kützing | 8 | 13 | 10 | 8 | 5 | 4 | Ep | i | i | c | Freshwater | ||
| Gomphonema subclavatum (Grunow) Grunow | 7 | 6 | 9 | Ep | i | al | c | Freshwater | |||||
| Halamphora normanii (Rabenhorst) Levkov | 15 | 15 | 15 | B | i | al | c | Freshwater, terrestrial | |||||
| Hannaea arcus (Ehrenberg) R.M. Patrick | 3 | 1 | 1 | L | hb | al | aa | Freshwater | |||||
| Hantzschia cf. abundans Lange-Bertalot | 4 | 2 | B | i | al | c | Freshwater | ||||||
| Luticola mutica (Kützing) D.G. Mann | 10 | 13 | B Ep | hl | al | c | Marine, brackish, freshwater, terrestrial | ||||||
| Mayamaea atomus (Kützing) Lange-Bertalot | 10 | 8 | B Ep | i | al | c | Marine, brackish, freshwater, terrestrial | ||||||
| Navicula cincta (Ehrenberg) Ralfs | 3 | 7 | 5 | 4 | 5 | 5 | 2 | B Ep | hl | al | c | Marine, brackish, freshwater, terrestrial | |
| Navicula leptostriata Jørgensen | 3 | B | hb | ac | c | Freshwater | |||||||
| Navicula minima Grunow | 10 | 12 | 5 | B Ep | i | al | c | Freshwater | |||||
| Neidium dubium (Ehenberg) Cleve | 3 | B | i | al | c | Marine | |||||||
| Nitzschia palea (Kützing) W. Smith | 15 | 15 | 15 | B | i | al | c | Freshwater | |||||
| N. recta Hantzsch ex Rabenhorst | 2 | B | i | al | c | Freshwater | |||||||
| Odontidium mesodon Kützing (Ehrenberg) | 3 | 1 | 1 | B Ep | hb | al | aa | Freshwater | |||||
| Pinnularia brevicostata Cleve | 12 | 3 | B | hb | i | b | Freshwater, terrestrial | ||||||
| Pinnularia borealis Ehrenberg | 6 | 8 | 10 | B | i | ac | c | Freshwater, terrestrial | |||||
| Pinnularia intermedia (Lagerstedt) Cleve | 3 | 12 | B | i | i | aa | Freshwater | ||||||
| Pinnularia microstauron (Ehrenberg) Cleve | 10 | 6 | 6 | 4 | 6 | B | i | i | c | Freshwater, terrestrial | |||
| Pinnularia cf. subcapitata W. Gregory | 15 | 15 | 15 | 8 | 15 | 14 | B | hb | ac | c | Freshwater, terrestrial | ||
| Pinnularia viridis (Nitzsch) Ehrenberg | 2 | 4 | B | i | i | c | Freshwater, terrestrial | ||||||
| Planothidium lanceolatum (Brébisson ex Kützing) Lange-Bertalot | 15 | 15 | 15 | 15 | 15 | 10 | 8 | Ep | i | al | c | Freshwater | |
| Psammothidium ventrale (Krasske) Bukhtiyarova and Round | 3 | 1 | Ep | hb | ac | aa | Freshwater | ||||||
| Pseudostaurosira brevistriata (Grunow) D.M. Williams and Round | 5 | 7 | 1 | L Ep | i | al | c | Freshwater | |||||
| Pseudostaurosira medliniae D.M. Williams and E.A Morales | 1 | 1 | L Ep | i | al | c | Freshwater | ||||||
| Rhopalodia acuminata Krammer in Lange-Bertalot and Krammer | 4 | 2 | 15 | 15 | Ep | i | al | b | Freshwater | ||||
| Sellaphora bacillum (Ehrenberg) D.G. Mann | 5 | 3 | B | i | al | c | Freshwater | ||||||
| Sellaphora pupula (Kützing) Mereschkovsky | 3 | 6 | B | hl | i | c | Freshwater | ||||||
| Sellaphora seminulum (Grunow) D.G. Mann | 5 | 2 | B Ep | i | ac | c | Freshwater | ||||||
| Staurosira construens Ehrenberg | 8 | 10 | 10 | 5 | 8 | 6 | L Ep | i | al | c | Freshwater | ||
| Staurosira venter (Ehrenberg) Cleve and J.D. Möller | 3 | L Ep | i | al | c | Freshwater | |||||||
| Staurosirella pinnata (Ehrenberg) D.M. Williams and Round | 2 | 2 | 2 | L | hl | al | c | Marine, freshwater | |||||
| Surirella brebissonii Krammer and Lange-Bertalot | 4 | 1 | B | i | al | c | Freshwater | ||||||
| Tabellaria flocculosa (Roth) Kützing | 5 | 2 | P Ep | hb | ac | aa | Freshwater, terrestrial | ||||||
| Tabellaria fenestrata (Lyngbye) Kützing | 4 | 8 | P | i | i | b | Freshwater | ||||||
| Ulnaria danica (Kützing) Compère and Bukhtiyarova | 1 | 3 | 2 | 2 | 1 | 1 | Ep L | i | al | c | Freshwater | ||
| Total number: 55 | 31 | 20 | 20 | 20 | 14 | 15 | 26 | 18 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fazlutdinova, A.I.; Allaguvatova, R.Z.; Gaysina, L.A. Ecotonic Communities of Diatoms in the Southeastern Part of the Kamchatka Peninsula. Earth 2023, 4, 209-222. https://doi.org/10.3390/earth4020011
Fazlutdinova AI, Allaguvatova RZ, Gaysina LA. Ecotonic Communities of Diatoms in the Southeastern Part of the Kamchatka Peninsula. Earth. 2023; 4(2):209-222. https://doi.org/10.3390/earth4020011
Chicago/Turabian StyleFazlutdinova, Alfiya I., Rezeda Z. Allaguvatova, and Lira A. Gaysina. 2023. "Ecotonic Communities of Diatoms in the Southeastern Part of the Kamchatka Peninsula" Earth 4, no. 2: 209-222. https://doi.org/10.3390/earth4020011