The Therapeutic Potential of Oxyhydrogen Gas in Oncology: A Study on Epstein–Barr Virus-Immortalised B-Lymphoblastoid (TK6) Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Safety
3. Rationale
4. Materials and Methods
4.1. Sterilization
4.2. Infusion
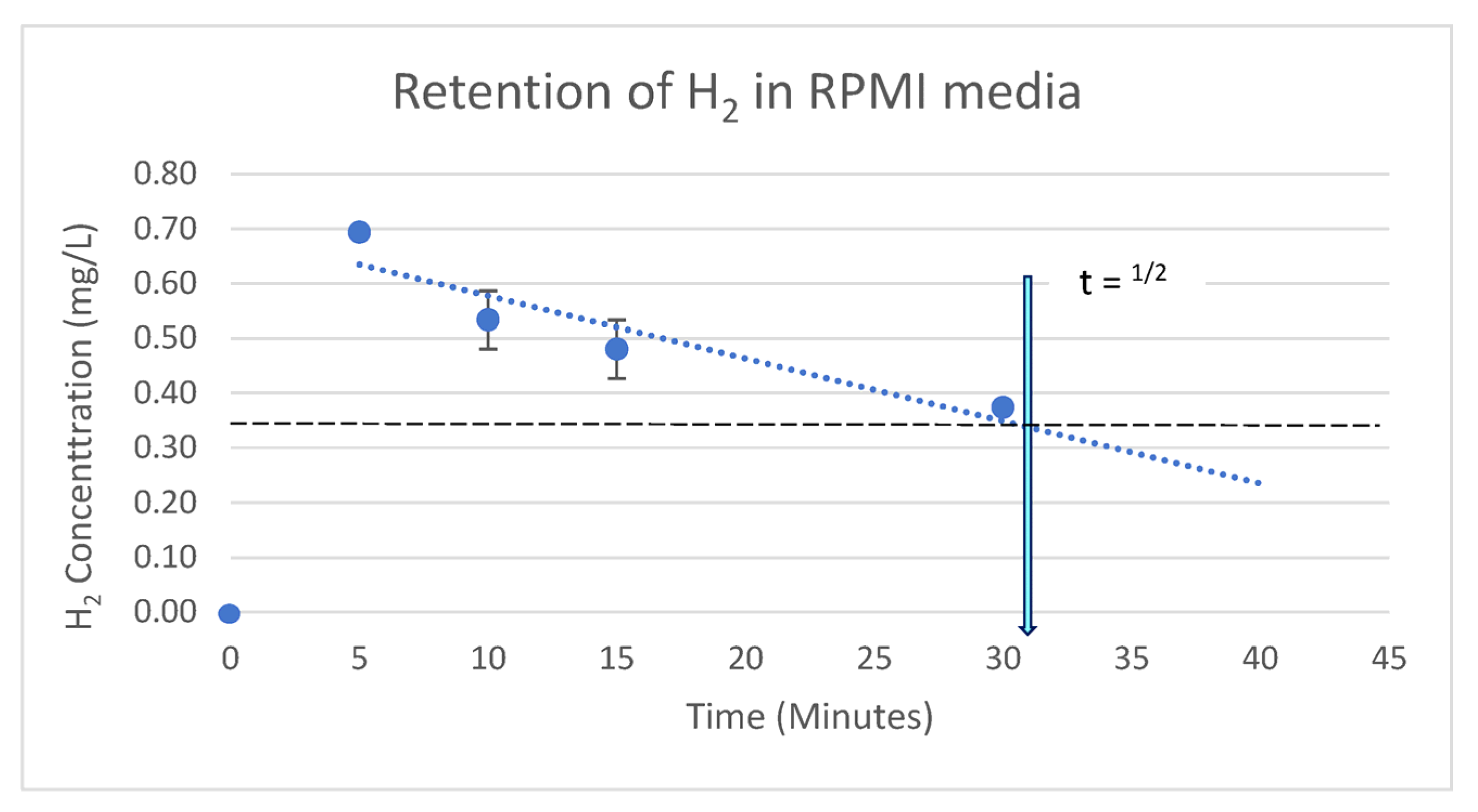
4.3. Retention of H2 in RPMI 1640 Cell Growth Medium
4.4. Experimental Design: Acute (Single) Treatment
4.5. Experimental Design: Chronic (Daily) Treatment
4.6. Cell Numeration Assay
4.7. Mitogen Treatment
4.8. Flow Cytometry
4.9. Statistical Analysis
5. Results
5.1. pH
5.2. Retention of Infused H2 in Cell Media
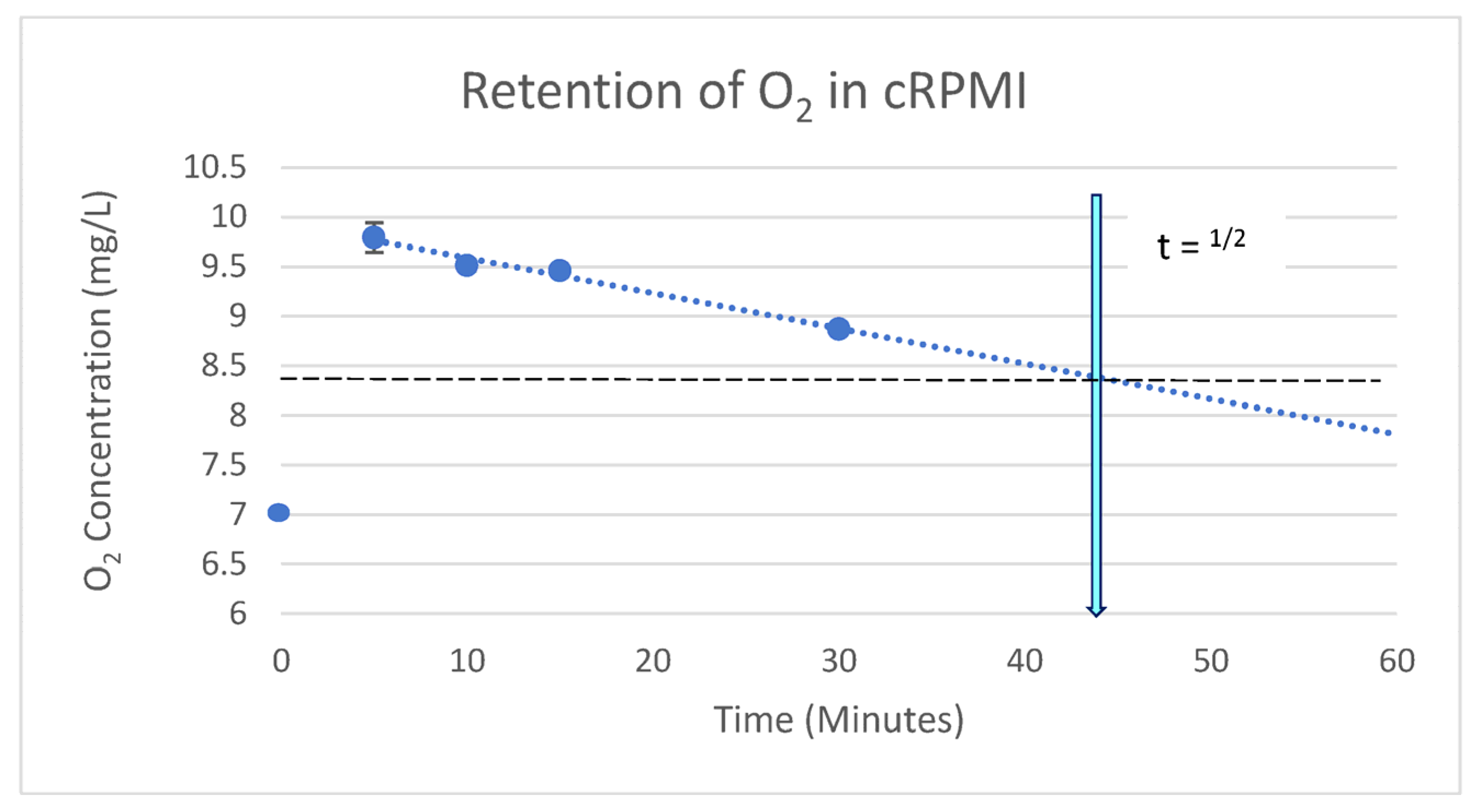
5.3. Retention of Infused O2 in Cell Media
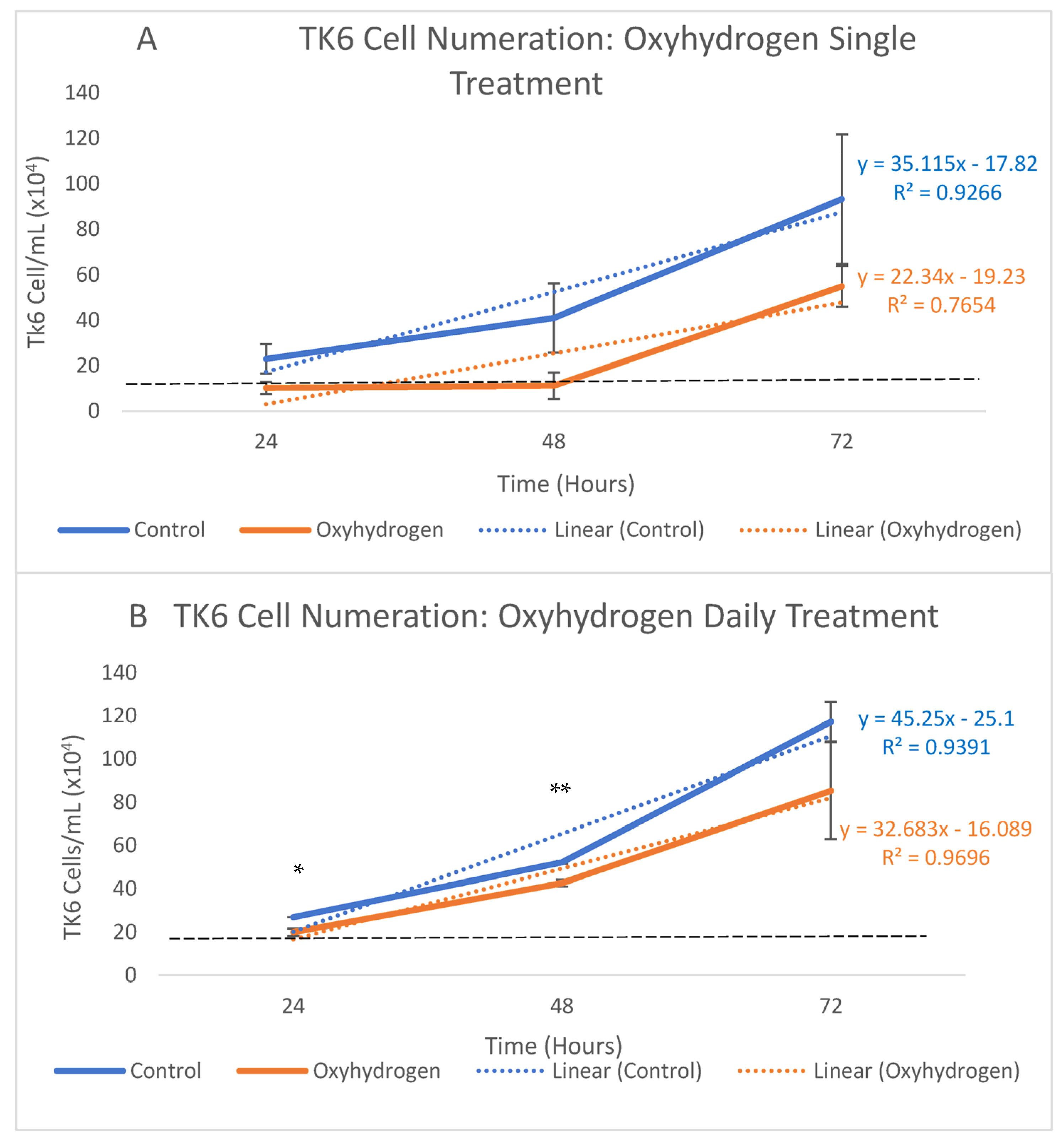
5.4. Cell Proliferation Assays
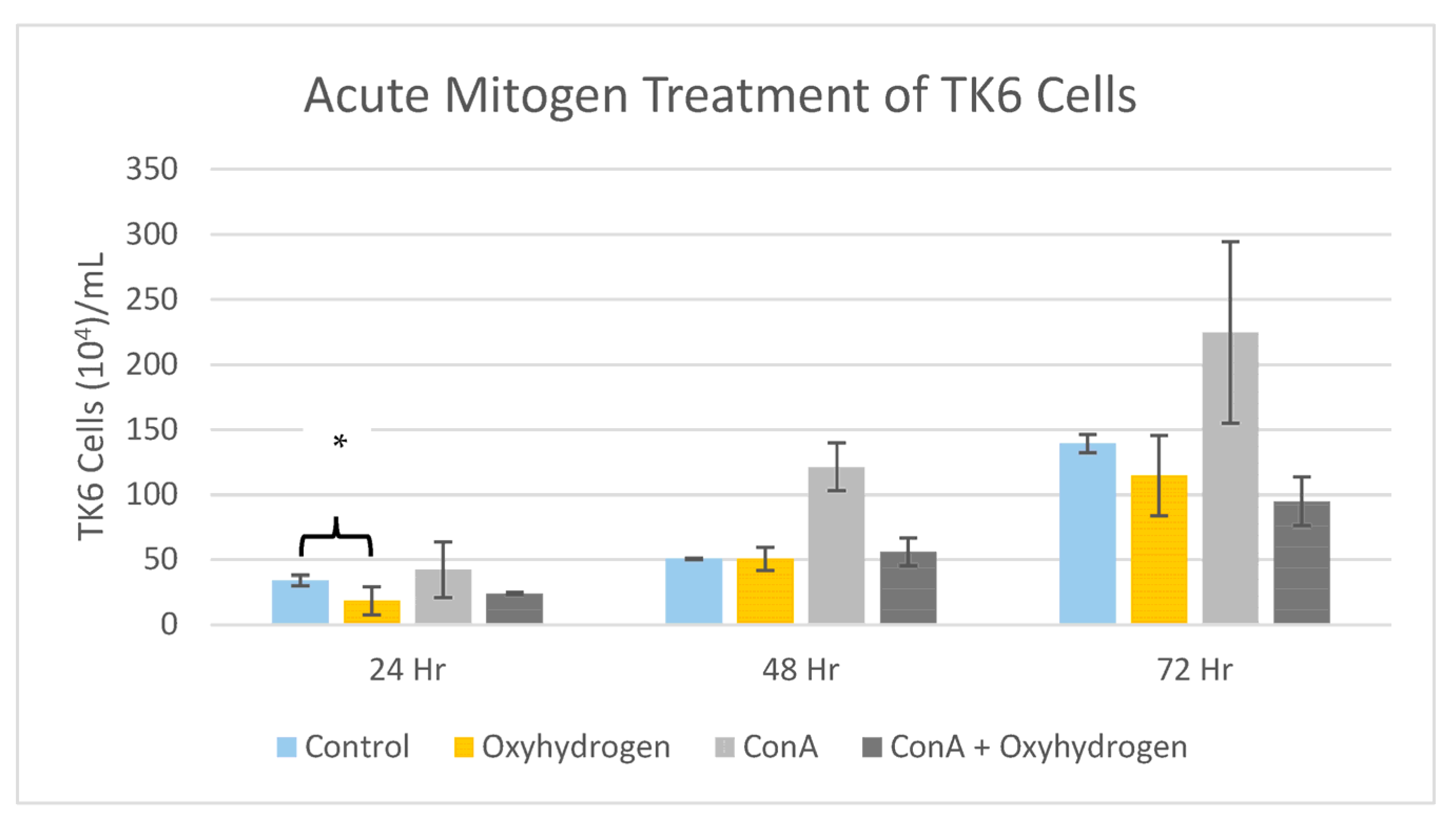
5.5. Mitogen Stimulation Assay
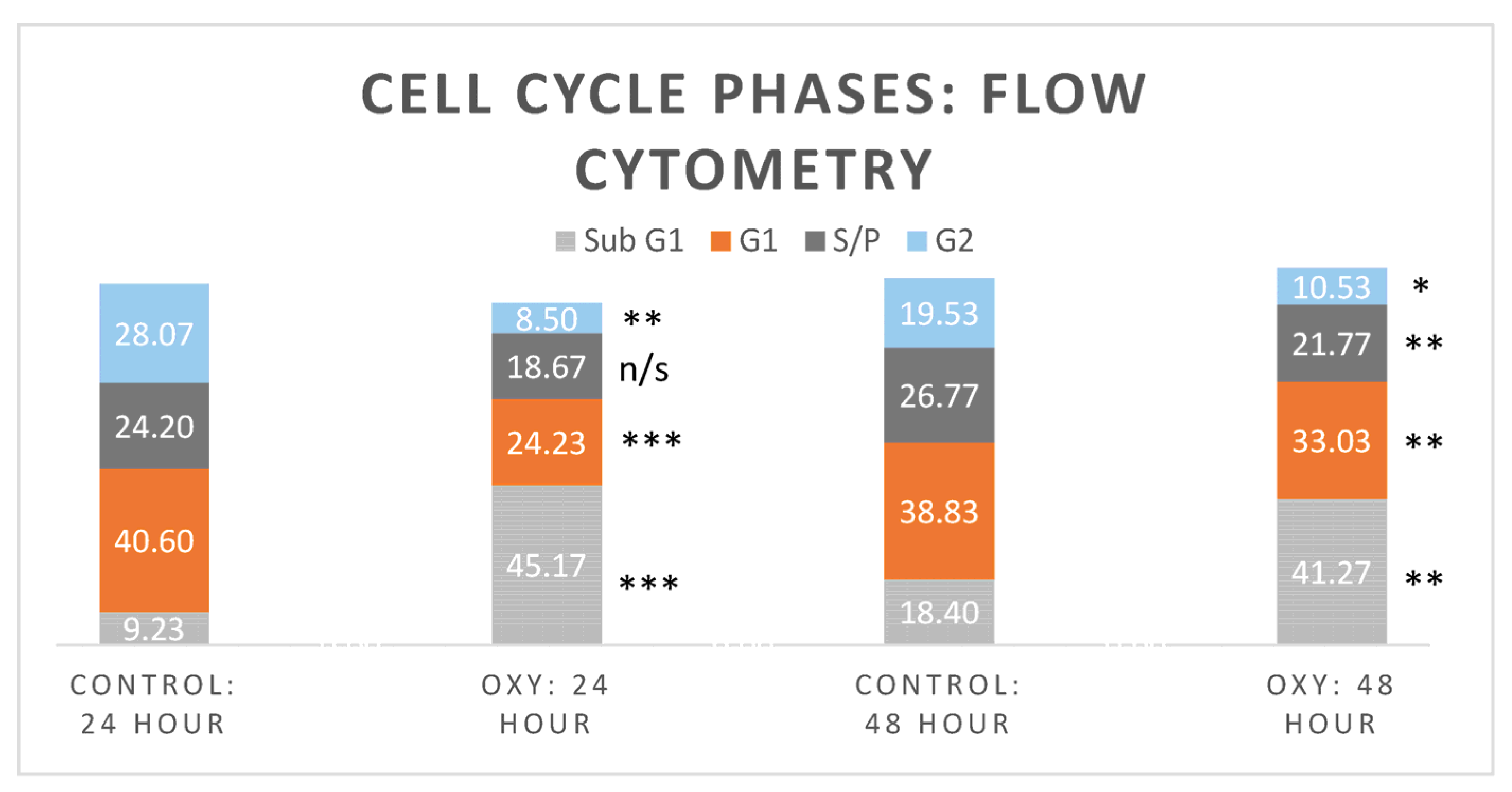
5.6. Flow Cytometry
6. Discussion
7. Summary
8. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| DMEM | Dulbecco’s modified eagle medium |
| EBV | Epstein–Barr Virus |
| HER2 | Human epidermal growth factor receptor-2 |
| HRW | Hydrogen-rich water |
| MAPK | Mitogen-activated protein kinase |
| MCP-1 | Monocyte chemotactic protein-1 |
| NFκB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| PTK | Protein tyrosine kinase |
| ROS/RNS | Reactive oxygen/nitrogen species |
| RPMI | Roswell Park Medical Institute 1640 |
| TNFα | Tumor necrosis factor-α |
| VEGF | Vascular endothelial growth factor-induced |
References
- Ohta, S. Molecular hydrogen as a novel antioxidant: Overview of the advantages of hydrogen for medical applications. Methods Enzymol. 2015, 555, 289–317. [Google Scholar]
- Kura, B.; Bagchi, A.K.; Singal, P.K.; Barancik, M.; LeBaron, T.W.; Valachova, K.; Šoltés, L.; Slezák, J. Molecular hydrogen: Potential in mitigating oxidative-stress-induced radiation injury. Can. J. Physiol. Pharmacol. 2019, 97, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, J.E.; Branco, L.G. Recent advances in molecular hydrogen research reducing exercise-induced oxidative stress and inflammation. Curr. Pharm. Des. 2021, 27, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Deryugina, A.V.; Danilova, D.A.; Brichkin, Y.D.; Taranov, E.V.; Nazarov, E.I.; Pichugin, V.V.; Medvedev, A.P.; Riazanov, M.V.; Fedorov, S.A.; Smorkalov, A.Y.; et al. Molecular hydrogen exposure improves functional state of red blood cells in the early postoperative period: A randomized clinical study. Med. Gas Res. 2023, 13, 59. [Google Scholar]
- Kajiyama, S.; Hasegawa, G.; Asano, M.; Hosoda, H.; Fukui, M.; Nakamura, N.; Kitawaki, J.; Imai, S.; Nakano, K.; Ohta, M.; et al. Supplementation of hydrogen-rich water improves lipid and glucose metabolism in patients with type 2 diabetes or impaired glucose tolerance. Nutr. Res. 2008, 28, 137–143. [Google Scholar] [CrossRef]
- Nishimaki, K.; Asada, T.; Ohsawa, I.; Nakajima, E.; Ikejima, C.; Yokota, T.; Kamimura, N.; Ohta, S. Effects of molecular hydrogen assessed by an animal model and a randomized clinical study on mild cognitive impairment. Curr. Alzheimer Res. 2018, 15, 482–492. [Google Scholar] [CrossRef]
- Akagi, J.; Baba, H. Hydrogen gas restores exhausted CD8+ T cells in patients with advanced colorectal cancer to improve prognosis. Oncol. Rep. 2019, 41, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Kong, X.; Mu, F.; Lu, T.; Du, D.; Xu, K. Hydrogen–oxygen therapy can alleviate radiotherapy-induced hearing loss in patients with nasopharyngeal cancer. Ann. Palliat. Med. 2019, 8, 74651–74751. [Google Scholar] [CrossRef]
- Chen, J.B.; Kong, X.F.; Mu, F.; Lu, T.Y.; Lu, Y.Y.; Xu, K.C. Hydrogen therapy can be used to control tumor progression and alleviate the adverse events of medications in patients with advanced non-small cell lung cancer. Med. Gas Res. 2020, 10, 75. [Google Scholar]
- Itoh, T.; Fujita, Y.; Ito, M.; Masuda, A.; Ohno, K.; Ichihara, M.; Kojima, T.; Nozawa, Y.; Ito, M. Molecular hydrogen suppresses FcεRI-mediated signal transduction and prevents degranulation of mast cells. Biochem. Biophys. Res. Commun. 2009, 389, 651–656. [Google Scholar] [CrossRef]
- Niu, Y.; Nie, Q.; Dong, L.; Zhang, J.; Liu, S.F.; Song, W.; Wang, X.; Wu, G.; Song, D. Hydrogen attenuates allergic inflammation by reversing energy metabolic pathway switch. Sci. Rep. 2020, 10, 1962. [Google Scholar] [CrossRef]
- Rochette, L.; Zeller, M.; Cottin, Y.; Vergely, C. Antitumor activity of protons and molecular hydrogen: Underlying mechanisms. Cancers 2021, 13, 893. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.C.; Li, T.T.; Wan, Q.; Zhang, X.; Sun, L.Y.; Zhang, Y.R.; Lai, P.C.; Li, W.Z. Molecular hydrogen mediates neurorestorative effects after stroke in diabetic rats: The TLR4/NF-κB inflammatory pathway. J. Neuroimmune Pharmacol. 2023, 18, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Slezak, J.; Kura, B.; LeBaron, T.W.; Singal, P.K.; Buday, J.; Barancik, M. Oxidative stress and pathways of molecular hydrogen effects in medicine. Curr. Pharm. Des. 2021, 27, 610–625. [Google Scholar] [CrossRef] [PubMed]
- You, I.S.; Sharma, S.; Fadriquela, A.; Bajgai, J.; Thi, T.T.; Rahman, M.H.; Sung, J.; Kwon, H.U.; Lee, S.Y.; Kim, C.S.; et al. Antioxidant properties of hydrogen gas attenuates oxidative stress in airway epithelial cells. Molecules 2021, 26, 6375. [Google Scholar] [CrossRef]
- Jiang, Y.; Liu, G.; Zhang, L.; Cheng, S.; Luo, C.; Liao, Y.; Guo, S. Therapeutic efficacy of hydrogen-rich saline alone and in combination with PI3K inhibitor in non-small cell lung cancer. Mol. Med. Rep. 2018, 18, 2182–2190. [Google Scholar] [CrossRef]
- Iuchi, K.; Nishimaki, K.; Kamimura, N.; Ohta, S. Molecular hydrogen suppresses free-radical-induced cell death by mitigating fatty acid peroxidation and mitochondrial dysfunction. Can. J. Physiol. Pharmacol. 2019, 97, 999–1005. [Google Scholar] [CrossRef]
- Begum, R.; Kim, C.S.; Fadriquela, A.; Bajgai, J.; Jing, X.; Kim, D.H.; Kim, S.K.; Lee, K.J. Molecular hydrogen protects against oxidative stress-induced RAW 264.7 macrophage cells through the activation of Nrf2 and inhibition of MAPK signaling pathway. Mol. Cell. Toxicol. 2020, 16, 103–118. [Google Scholar] [CrossRef]
- Deryugina, A.V.; Danilova, D.A.; Pichugin, V.V.; Brichkin, Y.D. The Effect of Molecular Hydrogen on Functional States of Erythrocytes in Rats with Simulated Chronic Heart Failure. Life 2023, 13, 418. [Google Scholar] [CrossRef]
- Botek, M.; Krejčí, J.; McKune, A.J.; Sládečková, B.; Naumovski, N. Hydrogen rich water improved ventilatory, perceptual and lactate responses to exercise. Int. J. Sports Med. 2019, 40, 879–885. [Google Scholar] [CrossRef]
- Akagi, J.; Baba, H. Hydrogen gas activates coenzyme Q10 to restore exhausted CD8+ T cells, especially PD-1+ Tim3+ terminal CD8+ T cells, leading to better nivolumab outcomes in patients with lung cancer. Oncol. Lett. 2020, 20, 258. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.J.; Wei, C.H.; Chen, A.L.; Sun, X.C.; Guo, G.Y.; Zou, X.; Shi, J.D.; Lai, P.Z.; Zheng, Z.G.; Zhong, N.S. Hydrogen/oxygen mixed gas inhalation improves disease severity and dyspnea in patients with Coronavirus disease 2019 in a recent multicenter, open-label clinical trial. J. Thorac. Dis. 2020, 12, 3448. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Yuan, M.; Song, J.; Chen, X.; Yang, H. Hydrogen gas from inflammation treatment to cancer therapy. ACS Nano 2019, 13, 8505–8511. [Google Scholar] [CrossRef] [PubMed]
- Radyuk, S.N. Mechanisms underlying the biological effects of molecular hydrogen. Curr. Pharm. Des. 2021, 27, 626–735. [Google Scholar] [CrossRef]
- Mantovani, A.; Marchesi, F.; Porta, C.; Sica, A.; Allavena, P. Inflammation and cancer: Breast cancer as a prototype. Breast 2007, 16, 27–33. [Google Scholar] [CrossRef]
- Jiang, X.; Shapiro, D.J. The immune system and inflammation in breast cancer. Mol. Cell. Endocrinol. 2014, 382, 673–682. [Google Scholar] [CrossRef]
- Ríos-Arrabal, S.; Artacho-Cordón, F.; León, J.; Román-Marinetto, E.; del Mar Salinas-Asensio, M.; Calvente, I.; Núñez, M.I. Involvement of free radicals in breast cancer. SpringerPlus 2013, 2, 404. [Google Scholar] [CrossRef]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in cancer therapy: The bright side of the moon. Exp. Mol. Med. 2020, 52, 192–203. [Google Scholar] [CrossRef]
- Wang, H.; Luo, Y.H.; Shen, G.N.; Piao, X.J.; Xu, W.T.; Zhang, Y.; Wang, J.R.; Feng, Y.C.; Li, J.Q.; Zhang, Y.; et al. Two novel 1,4-naphthoquinone derivatives induce human gastric cancer cell apoptosis and cell cycle arrest by regulating reactive oxygen species-mediated MAPK/Akt/STAT3 signaling pathways. Mol. Med. Rep. 2019, 20, 2571–2582. [Google Scholar] [CrossRef]
- Fan, Q.; Liang, X.; Xu, Z.; Li, S.; Han, S.; Xiao, Y.; Xu, Q.; Yuan, R.; Yang, S.; Gao, H. Pedunculoside inhibits epithelial-mesenchymal transition and overcomes Gefitinib-resistant non-small cell lung cancer through regulating MAPK and Nrf2 pathways. Phytomedicine 2023, 116, 154884. [Google Scholar] [CrossRef]
- Delos Reyes, F.S.L.G.; Mamaril, A.C.C.; Matias, T.J.P.; Tronco, M.K.V.; Samson, G.R.; Javier, N.D.; Fadriquela, A.; Antonio, J.M.; Sajo, M.E.J.V. The search for the elixir of life: On the therapeutic potential of alkaline reduced water in metabolic syndromes. Processes 2021, 9, 1876. [Google Scholar] [CrossRef]
- Asgharzadeh, F.; Tarnava, A.; Mostafapour, A.; Khazaei, M.; LeBaron, T.W. Hydrogen-rich water exerts anti-tumor effects comparable to 5-fluorouracil in a colorectal cancer xenograft model. World J. Gastrointest. Oncol. 2022, 14, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.F.; Hsu, Y.W.; Chen, W.K.; Ho, Y.C.; Lu, F.J. Enhanced induction of mitochondrial damage and apoptosis in human leukemia HL-60 cells due to electrolyzed-reduced water and glutathione. Biosci. Biotechnol. Biochem. 2009, 73, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Al Zahrani, S.M.; Omar, U.M.; Rahimulddin, S.A.; Al-Ghafari, A.B.; Aldahlawi, A.M.; Al Doghaither, H.A. Antiproliferative and apoptotic effects of the natural alkaline water (Zamzam) in breast cancer cell line MCF-7. J. Cancer Res. Ther. 2019, 15, 1098–1104. [Google Scholar] [PubMed]
- LeBaron, T.W.; Kura, B.; Kalocayova, B.; Tribulova, N.; Slezak, J. A new approach for the prevention and treatment of cardiovascular disorders. Molecular hydrogen significantly reduces the effects of oxidative stress. Molecules 2019, 24, 2076. [Google Scholar] [CrossRef]
- Fang, W.; Tang, L.; Wang, G.; Lin, J.; Liao, W.; Pan, W.; Xu, J. Molecular hydrogen protects human melanocytes from oxidative stress by activating Nrf2 signaling. J. Investig. Dermatol. 2020, 140, 2230–2241. [Google Scholar] [CrossRef]
- Frajese, G.V.; Benvenuto, M.; Mattera, R.; Giampaoli, S.; Ambrosin, E.; Bernardini, R.; Giganti, M.G.; Albonici, L.; Dus, I.; Manzari, V.; et al. Electrochemically Reduced Water Delays Mammary Tumors Growth in Mice and Inhibits Breast Cancer Cells Survival In Vitro. Evid. Based Complement. Altern. Med. 2018, 2018, 4753507. [Google Scholar] [CrossRef]
- Chen, J.B.; Kong, X.F.; Lv, Y.Y.; Qin, S.C.; Sun, X.J.; Mu, F.; Lu, T.Y.; Xu, K.C. “Real world survey” of hydrogen-controlled cancer: A follow-up report of 82 advanced cancer patients. Med. Gas Res. 2019, 9, 115. [Google Scholar]
- Yang, Y.; Zhu, Y.; Xi, X. Anti-inflammatory and antitumor action of hydrogen via reactive oxygen species. Oncol. Lett. 2018, 16, 2771–2776. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, P.Y.; Bao, W.; Chen, S.J.; Wu, F.S.; Zhu, P.Y. Hydrogen inhibits endometrial cancer growth via a ROS/NLRP3/caspase-1/GSDMD-mediated pyroptotic pathway. BMC Cancer 2020, 20, 28. [Google Scholar] [CrossRef]
- Dole, M.; Wilson, F.R.; Fife, W.P. Hyperbaric hydrogen therapy: A possible treatment for cancer. Science 1975, 190, 152–154. [Google Scholar] [CrossRef] [PubMed]
- Runtuwene, J.; Amitani, H.; Amitani, M.; Asakawa, A.; Cheng, K.C.; Inui, A. Hydrogen–water enhances 5-fluorouracil-induced inhibition of colon cancer. PeerJ 2015, 3, e859. [Google Scholar] [CrossRef]
- Meng, J.; Liu, L.; Wang, D.; Yan, Z.; Chen, G. Hydrogen gas represses the progression of lung cancer via down-regulating CD47. Biosci. Rep. 2020, 40, BSR20192761. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.Y.; Xie, F.; Zhang, Y.; Wang, T.T.; Ma, S.N.; Zhao, P.X.; Zhang, X.; Lebaron, T.W.; Yan, X.L.; Ma, X.M. Molecular hydrogen suppresses glioblastoma growth via inducing the glioma stem-like cell differentiation. Stem Cell Res. Ther. 2019, 10, 145. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.; Gao, J.; Wang, J.; Li, L.; Chen, G.; Dang, J.; Wang, Z.; Jin, Z.; Liu, X. Mechanism of hydrogen on cervical cancer suppression revealed by high-throughput RNA sequencing. Oncol. Rep. 2021, 46, 141. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Cui, H.; Xu, W. Hydrogen inhibits the proliferation and migration of gastric cancer cells by modulating lncRNA MALAT1/miR-124-3p/EZH2 axis. Cancer Cell Int. 2021, 21, 70. [Google Scholar] [CrossRef]
- Wong, Y.; Meehan, M.T.; Burrows, S.R.; Doolan, D.L.; Miles, J.J. Estimating the global burden of Epstein–Barr virus-related cancers. J. Cancer Res. Clin. Oncol. 2022, 148, 31–46. [Google Scholar] [CrossRef]
- Hsu, J.Y.; Chang, C.J.; Cheng, J.S. Survival, treatment regimens and medical costs of women newly diagnosed with metastatic triple-negative breast cancer. Sci. Rep. 2022, 12, 729. [Google Scholar] [CrossRef]
- McCloskey, C.; Ortega, M.T.; Nair, S.; Garcia, M.J.; Manevy, F. A systematic review of time and resource use costs of subcutaneous versus intravenous administration of oncology biologics in a hospital setting. PharmacoEcon. Open 2023, 7, 3–36. [Google Scholar] [CrossRef]
- Ryu, A.H.; Eckalbar, W.L.; Kreimer, A.; Yosef, N.; Ahituv, N. Use antibiotics in cell culture with caution: Genome-wide identification of antibiotic-induced changes in gene expression and regulation. Sci. Rep. 2017, 7, 7533. [Google Scholar] [CrossRef]
- Xu, X.; Wei, H.; Dong, Z.; Chen, Y.; Tian, Z. The differential effects of low dose and high dose concanavalin A on cytokine profile and their importance in liver injury. Inflamm. Res. 2006, 55, 144–152. [Google Scholar] [CrossRef]
- Wei, X.; Wang, S.; Zheng, W.; Wang, X.; Liu, X.; Jiang, S.; Pi, J.; Zheng, Y.; He, G.; Qu, W. Drinking water disinfection byproduct iodoacetic acid induces tumorigenic transformation of NIH3T3 cells. Environ. Sci. Technol. 2013, 47, 5913–5920. [Google Scholar] [CrossRef] [PubMed]
- Zhan, L.; Sakamoto, H.; Sakuraba, M.; Wu, D.S.; Zhang, L.S.; Suzuki, T.; Hayashi, M.; Honma, M. Genotoxicity of microcystin-LR in human lymphoblastoid TK6 cells. Mutat. Res./Genet. Toxicol. Environ. Mutagen. 2004, 557, 1–6. [Google Scholar] [CrossRef]
- Darzynkiewicz, Z.; Huang, X.; Zhao, H. Analysis of cellular DNA content by flow cytometry. Curr. Protoc. Immunol. 2017, 119, 5–7. [Google Scholar] [CrossRef] [PubMed]
- Manohar, S.M.; Shah, P.; Nair, A. Flow cytometry: Principles, applications and recent advances. Bioanalysis 2021, 13, 181–198. [Google Scholar] [CrossRef]
- Saitoh, Y.; Okayasu, H.; Xiao, L.; Harata, Y.; Miwa, N. Neutral pH hydrogen-enriched electrolyzed water achieves tumor-preferential clonal growth inhibition over normal cells and tumor invasion inhibition concurrently with intracellular oxidant repression. Oncol. Res. 2008, 17, 247–255. [Google Scholar] [CrossRef]
- Saitoh, Y.; Harata, Y.; Mizuhashi, F.; Nakajima, M.; Miwa, N. Biological safety of neutral-pH hydrogen-enriched electrolyzed water upon mutagenicity, genotoxicity and subchronic oral toxicity. Toxicol. Ind. Health 2010, 26, 203–216. [Google Scholar] [CrossRef]
- Wang, D.; Wang, L.; Zhang, Y.; Zhao, Y.; Chen, G. Hydrogen gas inhibits lung cancer progression through targeting SMC3. Biomed. Pharmacother. 2018, 104, 788–797. [Google Scholar] [CrossRef] [PubMed]
- Zan, R.; Wang, H.; Cai, W.; Ni, J.; Luthringer-Feyerabend, B.J.; Wang, W.; Peng, H.; Ji, W.; Yan, J.; Xia, J.; et al. Controlled release of hydrogen by implantation of magnesium induces P53-mediated tumor cells apoptosis. Bioact. Mater. 2022, 9, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, T.; Ito, M.; Hasegawa, S.; Teranishi, M.; Takeda, K.; Negishi, S.; Nishiwaki, H.; Takeda, J.I.; LeBaron, T.W.; Ohno, K. Molecular Hydrogen Enhances Proliferation of Cancer Cells That Exhibit Potent Mitochondrial Unfolded Protein Response. Int. J. Mol. Sci. 2022, 23, 2888. [Google Scholar] [CrossRef]
- Noor, M.N.Z.M.; Alauddin, A.S.; Wong, Y.H.; Looi, C.Y.; Wong, E.H.; Madhavan, P.; Yeong, C.H. A Systematic Review of Molecular Hydrogen Therapy in Cancer Management. Asian Pac. J. Cancer Prev. 2023, 24, 37. [Google Scholar] [CrossRef] [PubMed]
- Sabri, S.M.; Annuar, N.; Rahman, N.L.A.; Musairah, S.K.; Mutalib, H.A.; Subagja, I.K. Major Trends in Ageing Population Research: A Bibliometric Analysis from 2001 to 2021. Proceedings 2022, 82, 19. [Google Scholar]
- NHS Backlog Data Analysis. Available online: https://www.bma.org.uk/advice-and-support/nhs-delivery-and-workforce/pressures/nhs-backlog-data-analysis (accessed on 31 August 2023).






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Russell, G.; Thomas, A.D.; Nenov, A.; Mannings, G.; Hancock, J.T. The Therapeutic Potential of Oxyhydrogen Gas in Oncology: A Study on Epstein–Barr Virus-Immortalised B-Lymphoblastoid (TK6) Cells. Hydrogen 2023, 4, 746-759. https://doi.org/10.3390/hydrogen4040047
Russell G, Thomas AD, Nenov A, Mannings G, Hancock JT. The Therapeutic Potential of Oxyhydrogen Gas in Oncology: A Study on Epstein–Barr Virus-Immortalised B-Lymphoblastoid (TK6) Cells. Hydrogen. 2023; 4(4):746-759. https://doi.org/10.3390/hydrogen4040047
Chicago/Turabian StyleRussell, Grace, Adam D. Thomas, Alexander Nenov, Georgia Mannings, and John T. Hancock. 2023. "The Therapeutic Potential of Oxyhydrogen Gas in Oncology: A Study on Epstein–Barr Virus-Immortalised B-Lymphoblastoid (TK6) Cells" Hydrogen 4, no. 4: 746-759. https://doi.org/10.3390/hydrogen4040047