
Elucidating the Influence of Lipid Composition on Bilayer Perturbations Induced by the N-Terminal Region of the Huntingtin Protein

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
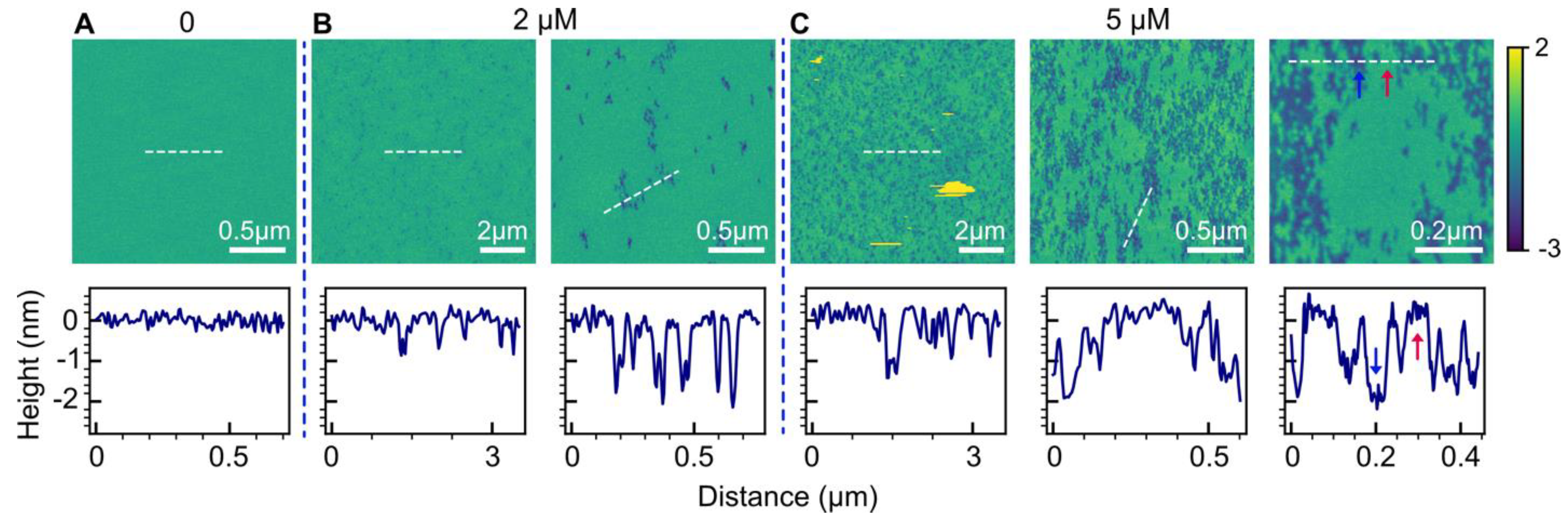
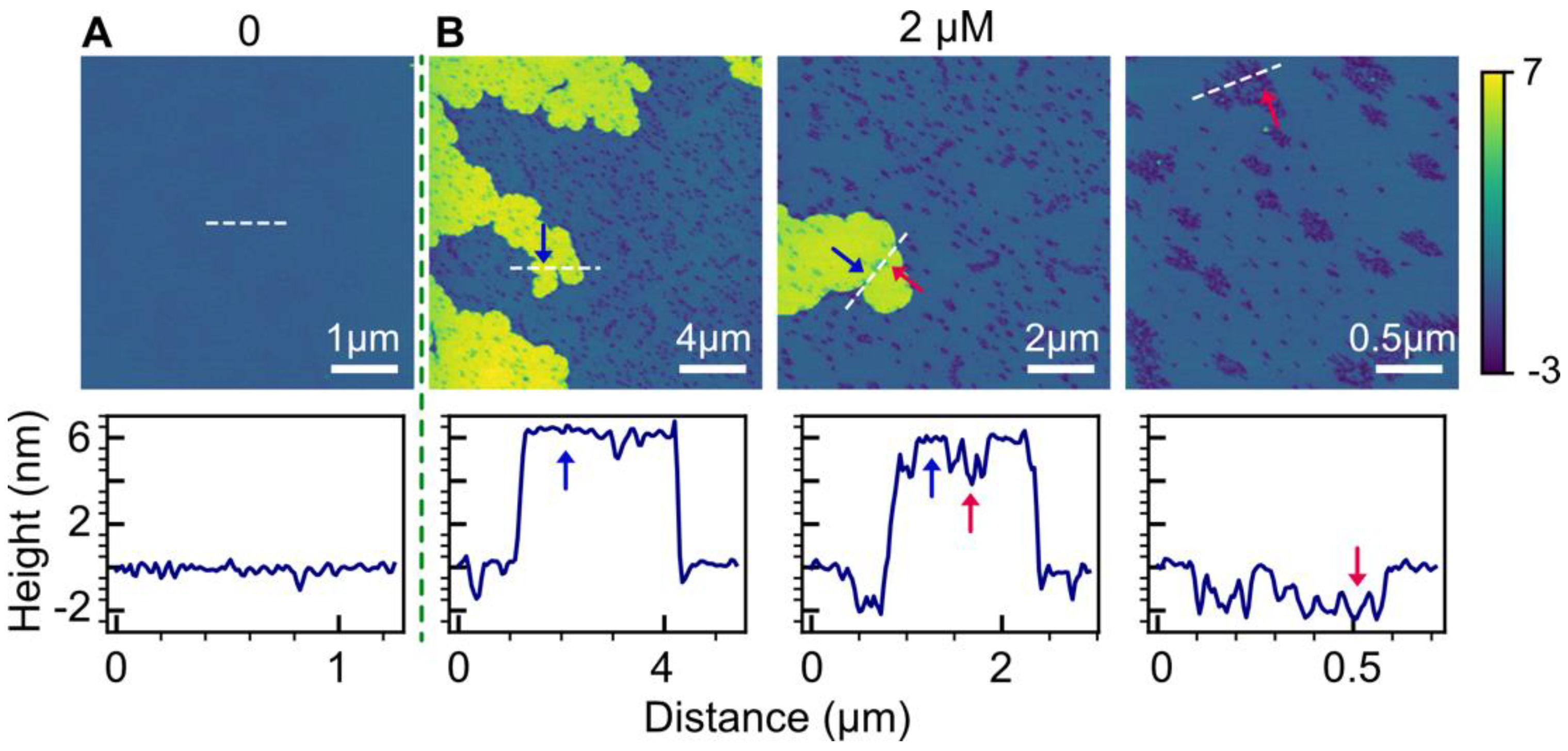
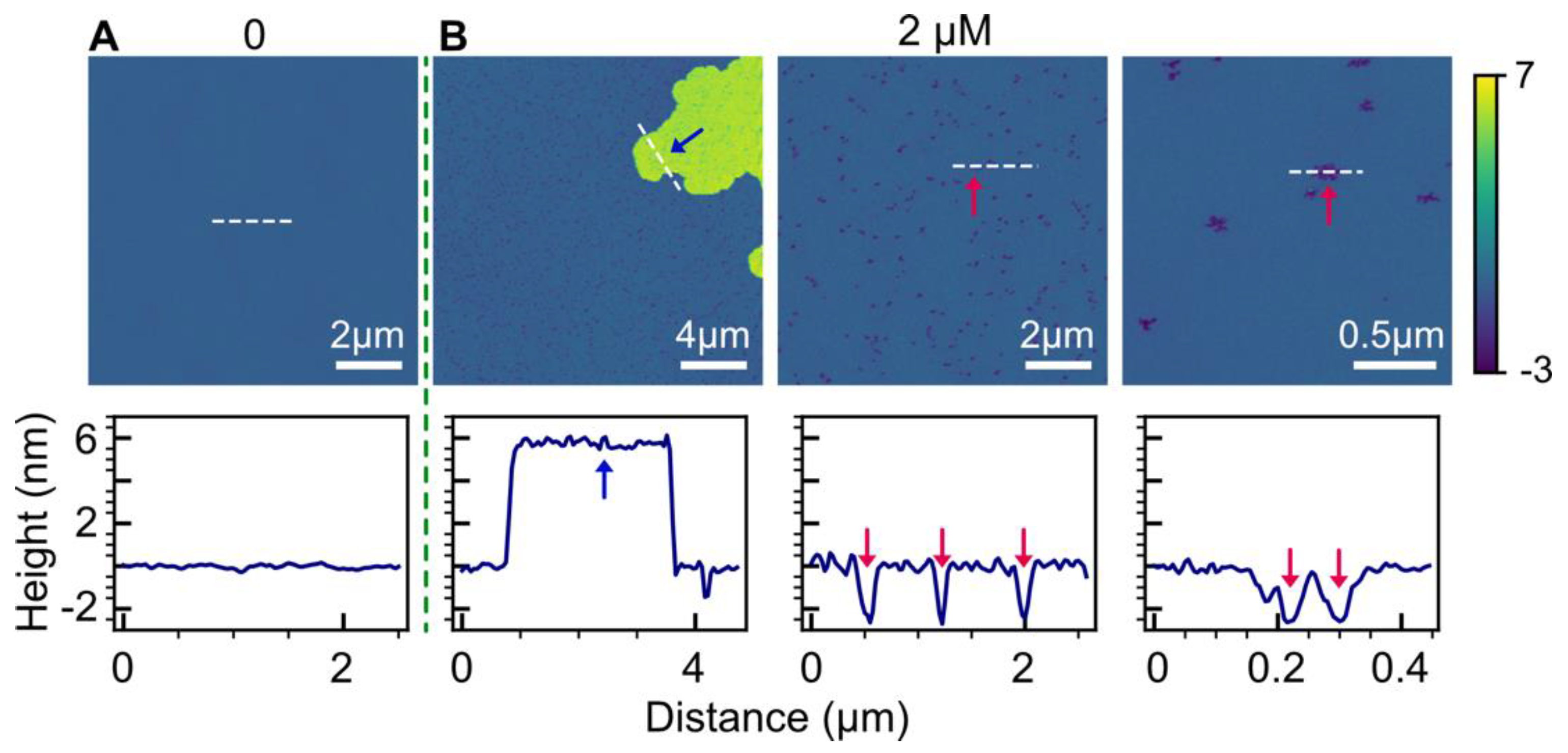
2.1. POPC Bilayers
2.2. Effect of Anionic POPG Lipids
2.3. Effect of Anionic Cardiolipin Lipids
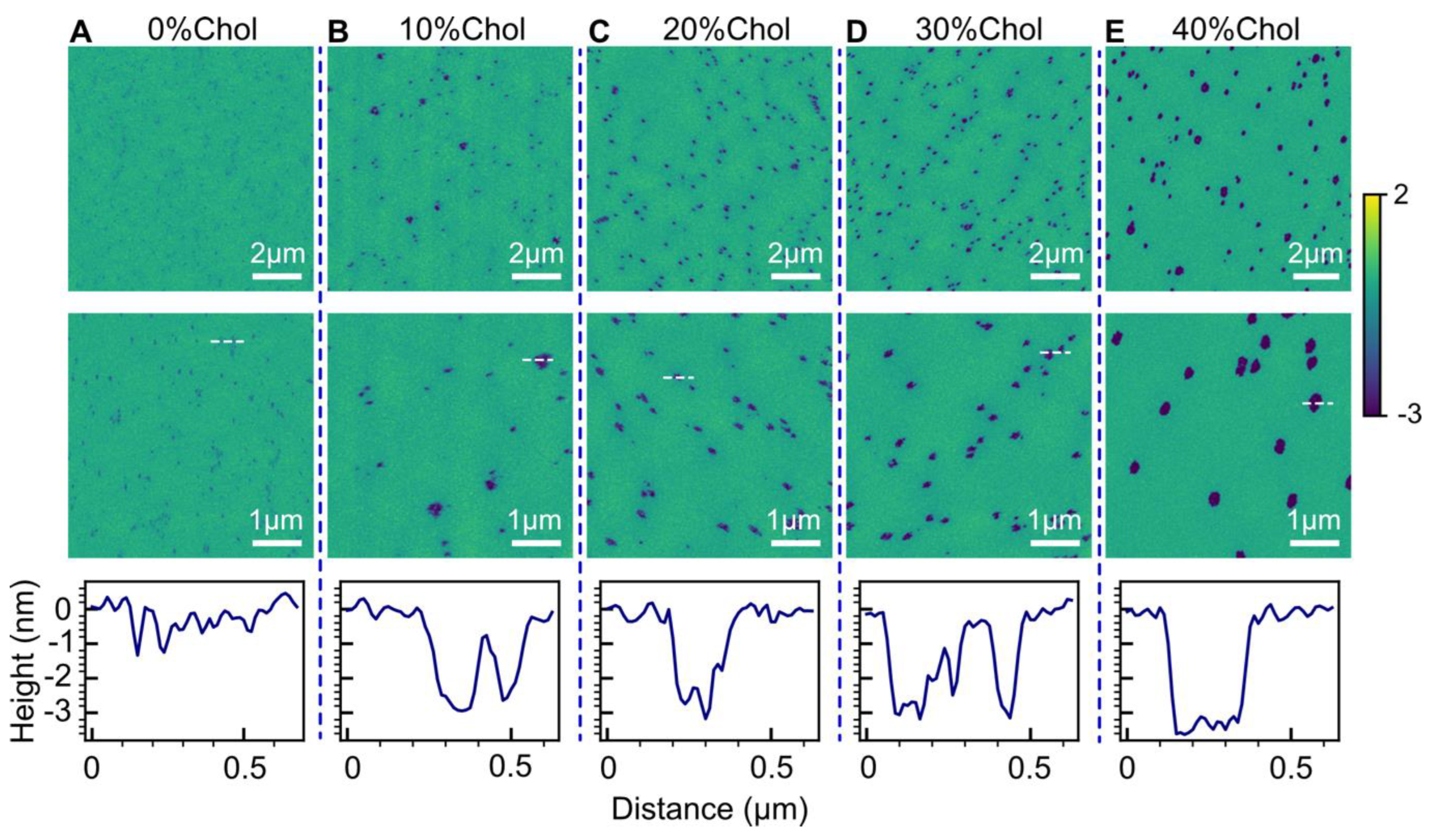
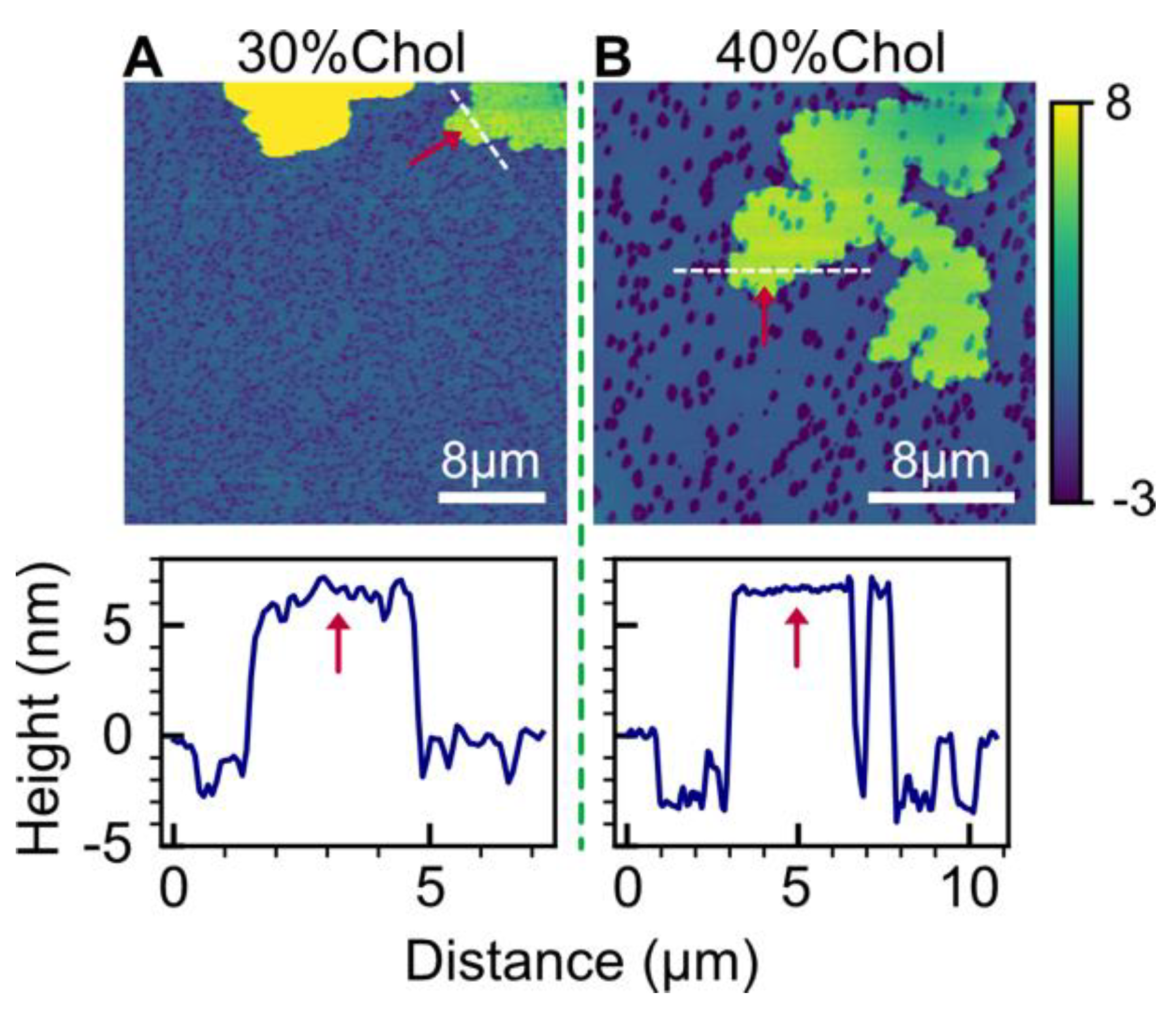
2.4. Effect of Cholesterol
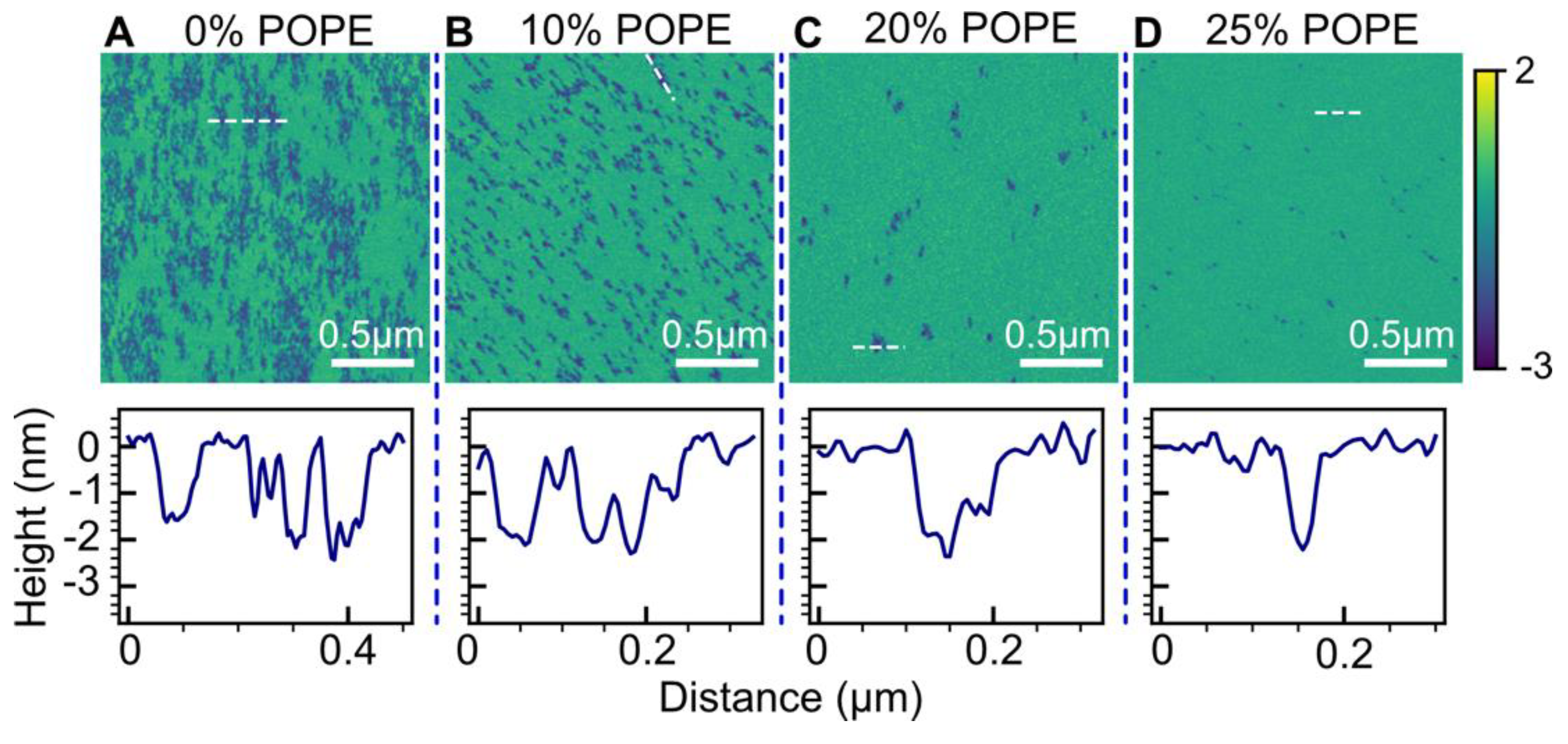
2.5. Effect of POPE
3. Discussion
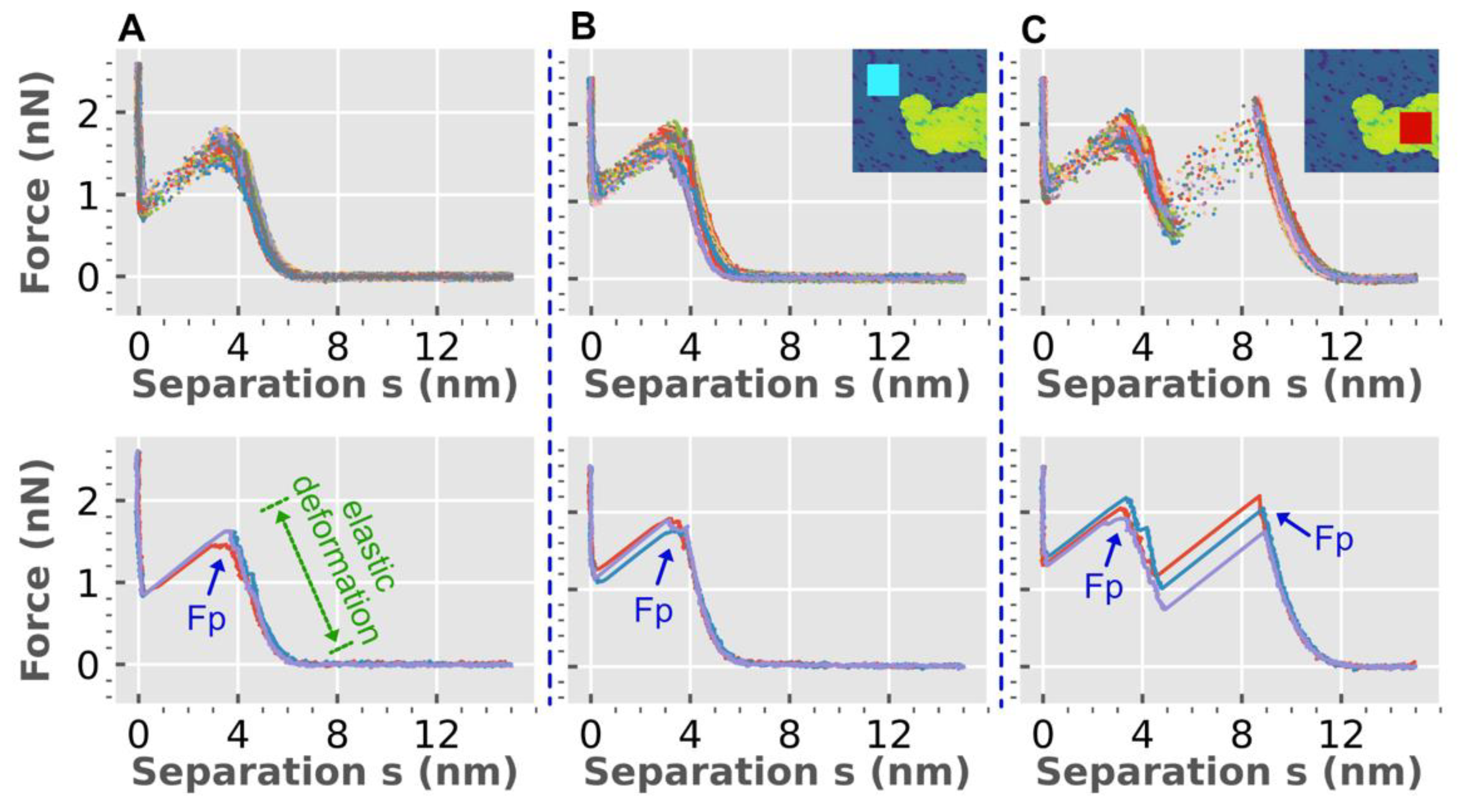
3.1. Bilayer Hole Formation
3.2. Implications of Force Spectroscopy
3.3. Effect of Cholesterol
3.4. Effect of POPE
3.5. Double-Bilayer Patches
3.6. Implications of Bilayer Fusion
3.7. Implications of Lipid Specificity
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Vijayvargia, R.; Epand, R.; Leitner, A.; Jung, T.Y.; Shin, B.; Jung, R.; Lloret, A.; Atwal, R.S.; Lee, H.; Lee, J.M.; et al. Huntingtin’s spherical solenoid structure enables polyglutamine tract-dependent modulation of its structure and function. eLife 2016, 5, e11184. [Google Scholar] [CrossRef]
- Saudou, F.; Humbert, S. The Biology of Huntingtin. Neuron 2016, 89, 910–926. [Google Scholar]
- Zuccato, C.; Valenza, M.; Cattaneo, E. Molecular Mechanisms and Potential Therapeutical Targets in Huntington’s Disease. Physiol. Rev. 2010, 90, 905–981. [Google Scholar] [CrossRef]
- Jurcau, A. Molecular Pathophysiological Mechanisms in Huntington’s Disease. Biomedicines 2022, 10, 1432. [Google Scholar] [CrossRef]
- Zoghbi, H.Y.; Orr, H.T. Glutamine repeats and neurodegeneration. Annu. Rev. Neurosci. 2000, 23, 217–247. [Google Scholar] [CrossRef]
- Bauer, P.O.; Nukina, N. The pathogenic mechanisms of polyglutamine diseases and current therapeutic strategies. J. Neurochem. 2009, 110, 1737–1765. [Google Scholar] [CrossRef]
- Gusella, J.F.; MacDonald, M.E. Molecular genetics: Unmasking polyglutamine triggers in neurodegenerative disease. Nat. Rev. Neurosci. 2000, 1, 109–115. [Google Scholar]
- Takahashi, T.; Katada, S.; Onodera, O. Polyglutamine Diseases: Where does Toxicity Come from? What is Toxicity? Where are We Going? J. Mol. Cell Biol. 2010, 2, 180–191. [Google Scholar] [CrossRef]
- Wetzel, R. Physical Chemistry of Polyglutamine: Intriguing Tales of a Monotonous Sequence. J. Mol. Biol. 2012, 421, 466–490. [Google Scholar]
- Rockabrand, E.; Slepko, N.; Pantalone, A.; Nukala, V.N.; Kazantsev, A.; Marsh, J.L.; Sullivan, P.G.; Steffan, J.S.; Sensi, S.L.; Thompson, L.M. The first 17 amino acids of Huntingtin modulate its sub-cellular localization, aggregation and effects on calcium homeostasis. Hum. Mol. Genet. 2007, 16, 61–77. [Google Scholar] [CrossRef]
- Ehrnhoefer, D.E.; Sutton, L.; Hayden, M.R. Small Changes, Big Impact: Posttranslational Modifications and Function of Huntingtin in Huntington Disease. Neuroscientist 2011, 17, 475–492. [Google Scholar] [CrossRef] [PubMed]
- Thakur, A.K.; Jayaraman, M.; Mishra, R.; Thakur, M.; Chellgren, V.M.; Byeon, I.J.L.; Anjum, D.H.; Kodali, R.; Creamer, T.P.; Conway, J.F.; et al. Polyglutamine disruption of the huntingtin exon 1 N terminus triggers a complex aggregation mechanism. Nat. Struct. Mol. Biol. 2009, 16, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.W.; Chelliah, Y.; Kim, S.W.; Otwinowski, Z.; Bezprozvanny, I. Secondary Structure of Huntingtin Amino-Terminal Region. Structure 2009, 17, 1205–1212. [Google Scholar] [CrossRef] [PubMed]
- Rossetti, G.; Cossio, P.; Laio, A.; Carloni, P. Conformations of the Huntingtin N-term in aqueous solution from atomistic simulations. FEBS Lett. 2011, 585, 3086–3089. [Google Scholar] [CrossRef] [PubMed]
- Dlugosz, M.; Trylska, J. Secondary Structures of Native and Pathogenic Huntingtin N-Terminal Fragments. J. Phys. Chem. B 2011, 115, 11597–11608. [Google Scholar] [CrossRef] [PubMed]
- Binette, V.; Cote, S.; Mousseau, N. Free-Energy Landscape of the Amino-Terminal Fragment of Huntingtin in Aqueous Solution. Biophys. J. 2016, 110, 1075–1088. [Google Scholar] [CrossRef]
- Michalek, M.; Salnikov, E.S.; Bechinger, B. Structure and Topology of the Huntingtin 1–17 Membrane Anchor by a Combined Solution and Solid-State NMR Approach. Biophys. J. 2013, 105, 699–710. [Google Scholar] [CrossRef]
- Cote, S.; Binette, V.; Salnikov, E.S.; Bechinger, B.; Mousseau, N. Probing the Huntingtin 1–17 Membrane Anchor on a Phospholipid Bilayer by Using All-Atom Simulations. Biophys. J. 2015, 108, 1187–1198. [Google Scholar] [CrossRef]
- Tao, M.X.; Pandey, N.K.; Barnes, R.; Han, S.; Langen, R. Structure of Membrane-Bound Huntingtin Exon 1 Reveals Membrane Interaction and Aggregation Mechanisms. Structure 2019, 27, 1570–1580. [Google Scholar] [CrossRef]
- Kim, M. Beta conformation of polyglutamine track revealed by a crystal structure of Huntingtin N-terminal region with insertion of three histidine residues. Prion 2013, 7, 221–228. [Google Scholar] [CrossRef]
- Cariulo, C.; Azzollini, L.; Verani, M.; Martufi, P.; Boggio, R.; Chiki, A.; Deguire, S.M.; Cherubini, M.; Gines, S.; Marsh, J.L.; et al. Phosphorylation of huntingtin at residue T3 is decreased in Huntington’s disease and modulates mutant huntingtin protein conformation. Proc. Natl. Acad. Sci. USA 2017, 114, E10809–E10818. [Google Scholar] [CrossRef] [PubMed]
- DeGuire, S.M.; Ruggeri, F.S.; Fares, M.B.; Chiki, A.; Cendrowska, U.; Dietler, G.; Lashuel, H.A. N-terminal Huntingtin (Htt) phosphorylation is a molecular switch regulating Htt aggregation, helical conformation, internalization, and nuclear targeting. J. Biol. Chem. 2018, 293, 18540–18558. [Google Scholar] [CrossRef] [PubMed]
- Monsellier, E.; Redeker, V.; Ruiz-Arlandis, G.; Bousset, L.; Melki, R. Molecular Interaction between the Chaperone Hsc70 and the N-terminal Flank of Huntingtin Exon 1 Modulates Aggregation. J. Biol. Chem. 2015, 290, 2560–2576. [Google Scholar] [CrossRef] [PubMed]
- Chaibva, M.; Jawahery, S.; Pilkington, A.W.; Arndt, J.R.; Sarver, O.; Valentine, S.; Matysiak, S.; Legleiter, J. Acetylation within the First 17 Residues of Huntingtin Exon 1 Alters Aggregation and Lipid Binding. Biophys. J. 2016, 111, 349–362. [Google Scholar] [CrossRef]
- Kegel, K.B.; Sapp, E.; Yoder, J.; Cuiffo, B.; Sobin, L.; Kim, Y.J.; Qin, Z.H.; Hayden, M.R.; Aronin, N.; Scott, D.L.; et al. Huntingtin associates with acidic phospholipids at the plasma membrane. J. Biol. Chem. 2005, 280, 36464–36473. [Google Scholar] [CrossRef]
- Kegel, K.B.; Sapp, E.; Alexander, J.; Valencia, A.; Reeves, P.; Li, X.Y.; Masso, N.; Sobin, L.; Aronin, N.; DiFiglia, M. Polyglutamine expansion in huntingtin alters its interaction with phospholipids. J. Neurochem. 2009, 110, 1585–1597. [Google Scholar] [CrossRef]
- Nagarajan, A.; Jawahery, S.; Matysiak, S. The Effects of Flanking Sequences in the Interaction of Polyglutamine Peptides with a Membrane Bilayer. J. Phys. Chem. B 2014, 118, 6368–6379. [Google Scholar] [CrossRef]
- Duennwald, M.L.; Jagadish, S.; Muchowski, P.J.; Lindquist, S. Flanking sequences profoundly alter polyglutamine toxicity in yeast. Proc. Natl. Acad. Sci. USA 2006, 103, 11045–11050. [Google Scholar] [CrossRef]
- Atwal, R.S.; Xia, J.; Pinchev, D.; Taylor, J.; Epand, R.M.; Truant, R. Huntingtin has a membrane association signal that can modulate huntingtin aggregation, nuclear entry and toxicity. Hum. Mol. Genet. 2007, 16, 2600–2615. [Google Scholar] [CrossRef]
- Jayaraman, M.; Kodali, R.; Sahoo, B.; Thakur, A.K.; Mayasundari, A.; Mishra, R.; Peterson, C.B.; Wetzel, R. Slow Amyloid Nucleation via alpha-Helix-Rich Oligomeric Intermediates in Short Polyglutamine-Containing Huntingtin Fragments. J. Mol. Biol. 2012, 415, 881–899. [Google Scholar] [CrossRef]
- Sivanandam, V.N.; Jayaraman, M.; Hoop, C.L.; Kodali, R.; Wetzel, R.; van der Wel, P.C.A. The Aggregation-Enhancing Huntingtin N-Terminus Is Helical in Amyloid Fibrils. J. Am. Chem. Soc. 2011, 133, 4558–4566. [Google Scholar] [CrossRef] [PubMed]
- Hoop, C.L.; Lin, H.K.; Kar, K.; Hou, Z.P.; Poirier, M.A.; Wetzel, R.; van der Wel, P.C.A. Polyglutamine Amyloid Core Boundaries and Flanking Domain Dynamics in Huntingtin Fragment Fibrils Determined by Solid-State Nuclear Magnetic Resonance. Biochemistry 2014, 53, 6653–6666. [Google Scholar] [CrossRef] [PubMed]
- Williamson, T.E.; Vitalis, A.; Crick, S.L.; Pappu, R.V. Modulation of Polyglutamine Conformations and Dimer Formation by the N-Terminus of Huntingtin. J. Mol. Biol. 2010, 396, 1295–1309. [Google Scholar] [CrossRef] [PubMed]
- Veldman, M.B.; Rios-Galdamez, Y.; Lu, X.H.; Gu, X.F.; Qin, W.; Li, S.; Yang, X.W.; Lin, S. The N17 domain mitigates nuclear toxicity in a novel zebrafish Huntington’s disease model. Mol. Neurodegener. 2015, 10, 67. [Google Scholar] [CrossRef]
- Gu, X.F.; Cantle, J.P.; Greiner, E.R.; Lee, C.Y.D.; Barth, A.M.; Gao, F.Y.; Park, C.S.; Zhang, Z.Q.; Sandoval-Miller, S.; Zhang, R.L.; et al. N17 Modifies Mutant Huntingtin Nuclear Pathogenesis and Severity of Disease in HD BAC Transgenic Mice. Neuron 2015, 85, 726–741. [Google Scholar] [CrossRef]
- Pandey, N.K.; Isas, J.M.; Rawat, A.; Lee, R.V.; Langen, J.; Pandey, P.; Langen, R. The 17-residue-long N terminus in huntingtin controls stepwise aggregation in solution and on membranes via different mechanisms. J. Biol. Chem. 2018, 293, 2597–2605. [Google Scholar] [CrossRef]
- Lakhani, V.V.; Ding, F.; Dokholyan, N.V. Polyglutamine Induced Misfolding of Huntingtin Exon1 is Modulated by the Flanking Sequences. PLoS Comput. Biol. 2010, 6, e1000772. [Google Scholar] [CrossRef]
- Huynen, C.; Willet, N.; Buell, A.K.; Duwez, A.S.; Jerome, C.; Dumoulin, M. Influence of the protein context on the polyglutamine length-dependent elongation of amyloid fibrils. BBA Proteins Proteom. 2015, 1854, 239–248. [Google Scholar] [CrossRef]
- Burke, K.A.; Kauffman, K.J.; Umbaugh, C.S.; Frey, S.L.; Legleiter, J. The Interaction of Polyglutamine Peptides with Lipid Membranes Is Regulated by Flanking Sequences Associated with Huntingtin. J. Biol. Chem. 2013, 288, 14993–15005. [Google Scholar] [CrossRef]
- Michalek, M.; Salnikov, E.S.; Werten, S.; Bechinger, B. Membrane Interactions of the Amphipathic Amino Terminus of Huntingtin. Biochemistry 2013, 52, 847–858. [Google Scholar] [CrossRef]
- Michalek, M.; Aisenbrey, C.; Bechinger, B. Investigation of membrane penetration depth and interactions of the amino-terminal domain of huntingtin: Refined analysis by tryptophan fluorescence measurement. Eur. Biophys. J. Biophy. 2014, 43, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Cote, S.; Wei, G.H.; Mousseau, N. Atomistic mechanisms of huntingtin N-terminal fragment insertion on a phospholipid bilayer revealed by molecular dynamics simulations. Proteins 2014, 82, 1409–1427. [Google Scholar] [CrossRef] [PubMed]
- Ceccon, A.; Schmidt, T.; Tugarinov, V.; Kotler, S.A.; Schwieters, C.D.; Clore, G.M. Interaction of Huntingtin Exon-1 Peptides with Lipid-Based Micellar Nanoparticles Probed by Solution NMR and Q-Band Pulsed EPR. J. Am. Chem. Soc. 2018, 140, 6199–6202. [Google Scholar] [CrossRef] [PubMed]
- Karanji, A.K.; Beasley, M.; Sharif, D.; Ranjbaran, A.; Legleiter, J.; Valentine, S.J. Investigating the interactions of the first 17 amino acid residues of Huntingtin with lipid vesicles using mass spectrometry and molecular dynamics. J. Mass Spectrom. 2020, 55, e4470. [Google Scholar] [CrossRef]
- Goksu, E.I.; Vanegas, J.M.; Blanchette, C.D.; Lin, W.C.; Longo, M.L. AFM for structure and dynamics of biomembranes. BBA Biomembr. 2009, 1788, 254–266. [Google Scholar] [CrossRef] [PubMed]
- El Kirat, K.; Morandat, S.; Dufrêne, Y.F. Nanoscale analysis of supported lipid bilayers using atomic force microscopy. BBA Biomembr. 2010, 1798, 750–765. [Google Scholar] [CrossRef]
- Picas, L.; Milhiet, P.E.; Hernández-Borrell, J. Atomic force microscopy: A versatile tool to probe the physical and chemical properties of supported membranes at the nanoscale. Chem. Phys. Lipids 2012, 165, 845–860. [Google Scholar] [CrossRef]
- Mescola, A.; Ragazzini, G.; Facci, P.; Alessandrini, A. The potential of AFM in studying the role of the nanoscale amphipathic nature of (lipo)-peptides interacting with lipid bilayers. Nanotechnology 2022, 33, 432001. [Google Scholar] [CrossRef]
- Burke, K.A.; Godbey, J.; Legleiter, J. Assessing mutant huntingtin fragment and polyglutamine aggregation by atomic force microscopy. Methods 2011, 53, 275–284. [Google Scholar] [CrossRef]
- Burke, K.A.; Hensal, K.M.; Umbaugh, C.S.; Chaibva, M.; Legleiter, J. Huntingtin disrupts lipid bilayers in a polyQ-length dependent manner. BBA Biomembr. 2013, 1828, 1953–1961. [Google Scholar] [CrossRef]
- Burke, K.A.; Yates, E.A.; Legleiter, J. Biophysical insights into how surfaces, including lipid membranes, modulate protein aggregation related to neurodegeneration. Front. Neurol. 2013, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- Chaibva, M.; Burke, K.A.; Legleiter, J. Curvature Enhances Binding and Aggregation of Huntingtin at Lipid Membranes. Biochemistry 2014, 53, 2355–2365. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.S.; Khadka, N.K.; She, F.; Cai, J.; Pan, J. Polyglutamine aggregates impair lipid membrane integrity and enhance lipid membrane rigidity. Biochim. Biophys. Acta 2016, 1858, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Manyes, S.; Sanz, F. Nanomechanics of lipid bilayers by force spectroscopy with AFM: A perspective. BBA Biomembr. 2010, 1798, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.J.; Khadka, N.K. Kinetic Defects Induced by Melittin in Model Lipid Membranes: A Solution Atomic Force Microscopy Study. J. Phys. Chem. B 2016, 120, 4625–4634. [Google Scholar] [CrossRef]
- Pan, J.J.; Dalzini, A.; Song, L.K. Cholesterol and phosphatidylethanolamine lipids exert opposite effects on membrane modulations caused by the M2 amphipathic helix. BBA Biomembr. 2019, 1861, 201–209. [Google Scholar] [CrossRef]
- Aryal, C.M.; Bui, N.N.; Song, L.K.; Pan, J.J. The N-terminal helices of amphiphysin and endophilin have different capabilities of membrane remodeling. BBA Biomembr. 2022, 1864, 183907. [Google Scholar] [CrossRef]
- Aryal, C.M.; Bui, N.N.; Khadka, N.K.; Song, L.K.; Pan, J.J. The helix 0 of endophilin modifies membrane material properties and induces local curvature. BBA Biomembr. 2020, 1862, 183397. [Google Scholar] [CrossRef]
- Khadka, N.K.; Aryal, C.M.; Pan, J.J. Lipopolysaccharide-Dependent Membrane Permeation and Lipid Clustering Caused by Cyclic Lipopeptide Colistin. ACS Omega 2018, 3, 17828–17834. [Google Scholar] [CrossRef]
- Adegbuyiro, A.; Sedighi, F.; Jain, P.; Pinti, M.V.; Siriwardhana, C.; Hollander, J.M.; Legleiter, J. Mitochondrial membranes modify mutant huntingtin aggregation. BBA Biomembr. 2021, 1863, 183663. [Google Scholar] [CrossRef]
- Beasley, M.; Groover, S.; Valentine, S.J.; Legleiter, J. Lipid headgroups alter huntingtin aggregation on membranes. BBA Biomembr. 2021, 1863, 183497. [Google Scholar] [CrossRef] [PubMed]
- Stetter, F.W.S.; Hugel, T. The Nanomechanical Properties of Lipid Membranes are Significantly Influenced by the Presence of Ethanol. Biophys. J. 2013, 104, 1049–1055. [Google Scholar] [CrossRef] [PubMed]
- Valenza, M.; Cattaneo, E. Emerging roles for cholesterol in Huntington’s disease. Trends Neurosci. 2011, 34, 474–486. [Google Scholar] [CrossRef]
- Karasinska, J.M.; Hayden, M.R. Cholesterol metabolism in Huntington disease. Nat. Rev. Neurol. 2011, 7, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Leoni, V.; Caccia, C. The impairment of cholesterol metabolism in Huntington disease. BBA Mol. Cell Biol. Lipids 2015, 1851, 1095–1105. [Google Scholar] [CrossRef]
- Gao, X.; Campbell, W.A.; Chaibva, M.; Jain, P.; Leslie, A.E.; Frey, S.L.; Legleiter, J. Cholesterol Modifies Huntingtin Binding to, Disruption of, and Aggregation on Lipid Membranes. Biochemistry 2016, 55, 92–102. [Google Scholar] [CrossRef]
- Veatch, S.L.; Polozov, I.V.; Gawrisch, K.; Keller, S.L. Liquid domains in vesicles investigated by NMR and fluorescence microscopy. Biophys. J. 2004, 86, 2910–2922. [Google Scholar] [CrossRef]
- Róg, T.; Pasenkiewicz-Gierula, M.; Vattulainen, I.; Karttunen, M. Ordering effects of cholesterol and its analogues. BBA Biomembr. 2009, 1788, 97–121. [Google Scholar] [CrossRef]
- Pan, J.J.; Tristram-Nagle, S.; Nagle, J.F. Effect of cholesterol on structural and mechanical properties of membranes depends on lipid chain saturation. Phys. Rev. E 2009, 80, 021931. [Google Scholar] [CrossRef]
- Brown, D.A.; London, E. Functions of lipid rafts in biological membranes. Annu. Rev. Cell Dev. Biol. 1998, 14, 111–136. [Google Scholar] [CrossRef]
- Kollmitzer, B.; Heftberger, P.; Rappolt, M.; Pabst, G. Monolayer spontaneous curvature of raft-forming membrane lipids. Soft Matter 2013, 9, 10877–10884. [Google Scholar] [CrossRef] [PubMed]
- Khadka, N.K.; Teng, P.; Cai, J.F.; Pan, J.J. Modulation of lipid membrane structural and mechanical properties by a peptidomimetic derived from reduced amide scaffold. BBA Biomembr. 2017, 1859, 734–744. [Google Scholar] [CrossRef] [PubMed]
- Han, X.J.; Achalkumar, A.S.; Cheetham, M.R.; Connell, S.D.A.; Johnson, B.R.G.; Bushby, R.J.; Evans, S.D. A Self-assembly Route for Double Bilayer Lipid Membrane Formation. Chemphyschem 2010, 11, 569–574. [Google Scholar] [CrossRef]
- Nagle, J.F.; Zhang, R.T.; TristramNagle, S.; Sun, W.J.; Petrache, H.I.; Suter, R.M. X-ray structure determination of fully hydrated L(alpha) phase dipalmitoylphosphatidylcholine bilayers. Biophys. J. 1996, 70, 1419–1431. [Google Scholar] [CrossRef] [PubMed]
- Quinn, P.J.; Wolf, C. An X-ray diffraction study of model membrane raft structures. FEBS J. 2010, 277, 4685–4698. [Google Scholar] [CrossRef]
- Yang, S.T.; Kreutzberger, A.J.B.; Lee, J.; Kiessling, V.; Tamm, L.K. The role of cholesterol in membrane fusion. Chem. Phys. Lipids 2016, 199, 136–143. [Google Scholar] [CrossRef]
- Parente, R.A.; Nir, S.; Szoka, F.C. Ph-Dependent Fusion of Phosphatidylcholine Small Vesicles—Induction by a Synthetic Amphipathic Peptide. J. Biol. Chem. 1988, 263, 4724–4730. [Google Scholar] [CrossRef]
- Murata, M.; Takahashi, S.; Kagiwada, S.; Suzuki, A.; Ohnishi, S. Ph-Dependent Membrane-Fusion and Vesiculation of Phospholipid Large Unilamellar Vesicles Induced by Amphiphilic Anionic and Cationic Peptides. Biochemistry 1992, 31, 1986–1992. [Google Scholar] [CrossRef]
- Martens, S.; McMahon, H.T. Mechanisms of membrane fusion: Disparate players and common principles. Nat. Rev. Mol. Cell Bio 2008, 9, 543–556. [Google Scholar] [CrossRef]
- Kabelka, I.; Pachler, M.; Prevost, S.; Letofsky-Papst, I.; Lohner, K.; Pabst, G.; Vacha, R. Magainin 2 and PGLa in Bacterial Membrane Mimics II: Membrane Fusion and Sponge Phase Formation. Biophys. J. 2020, 118, 612–623. [Google Scholar] [CrossRef]
- Kabelka, I.; Georgiev, V.; Marx, L.; Pajtinka, P.; Lohner, K.; Pabst, G.; Dimova, R.; Vacha, R. Magainin 2 and PGLa in bacterial membrane mimics III: Membrane fusion and disruption. Biophys. J. 2022, 121, 852–861. [Google Scholar] [CrossRef] [PubMed]
- Suopanki, J.; Götz, C.; Lutsch, G.; Schiller, J.; Harjes, P.; Herrmann, A.; Wanker, E.E. Interaction of huntingtin fragments with brain membranes—Clues to early dysfunction in Huntington’s disease. J Neurochem 2006, 96, 870–884. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.H.; Shirendeb, U.P. Mutant huntingtin, abnormal mitochondrial dynamics, defective axonal transport of mitochondria, and selective synaptic degeneration in Huntington’s disease. BBA Mol. Basis Dis. 2012, 1822, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.H.; Mao, P.Z.; Manczak, M. Mitochondrial structural and functional dynamics in Huntington’s disease. Brain Res. Rev. 2009, 61, 33–48. [Google Scholar] [CrossRef]
- Oliveira, J.M.A. Nature and cause of mitochondrial dysfunction in Huntington’s disease: Focusing on huntingtin and the striatum. J. Neurochem. 2010, 114, 1–12. [Google Scholar] [CrossRef]
- Damiano, M.; Galvan, L.; Deglon, N.; Brouillet, E. Mitochondria in Huntington’s disease. BBA Mol. Basis Dis. 2010, 1802, 52–61. [Google Scholar] [CrossRef]
- Bossy-Wetzel, E.; Petrilli, A.; Knott, A.B. Mutant huntingtin and mitochondrial dysfunction. Trends Neurosci. 2008, 31, 609–616. [Google Scholar] [CrossRef]
- Chen, H.C.; Chan, D.C. Mitochondrial dynamics-fusion, fission, movement, and mitophagy-in neurodegenerative diseases. Hum. Mol. Genet. 2009, 18, R169–R176. [Google Scholar] [CrossRef]
- Shirendeb, U.; Reddy, A.P.; Manczak, M.; Calkins, M.J.; Mao, P.Z.; Tagle, D.A.; Reddy, P.H. Abnormal mitochondrial dynamics, mitochondrial loss and mutant huntingtin oligomers in Huntington’s disease: Implications for selective neuronal damage. Hum. Mol. Genet. 2011, 20, 1438–1455. [Google Scholar] [CrossRef]
- Knott, A.B.; Perkins, G.; Schwarzenbacher, R.; Bossy-Wetzel, E. Mitochondrial fragmentation in neurodegeneration. Nat. Rev. Neurosci. 2008, 9, 505–518. [Google Scholar] [CrossRef]
- Kim, J.; Moody, J.P.; Edgerly, C.K.; Bordiuk, O.L.; Cormier, K.; Smith, K.; Beal, M.F.; Ferrante, R.J. Mitochondrial loss, dysfunction and altered dynamics in Huntington’s disease. Hum. Mol. Genet. 2010, 19, 3919–3935. [Google Scholar] [CrossRef] [PubMed]
- Khadka, N.K.; Ho, C.S.; Pan, J.J. Macroscopic and Nanoscopic Heterogeneous Structures in a Three-Component Lipid Bilayer Mixtures Determined by Atomic Force Microscopy. Langmuir 2015, 31, 12417–12425. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.S.; Khadka, N.K.; Pan, J. Sub-ten-nanometer heterogeneity of solid supported lipid membranes determined by solution atomic force microscopy. BBA Biomembr. 2015, 1858, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.J.; Sahoo, P.K.; Dalzini, A.; Hayati, Z.; Aryal, C.M.; Teng, P.; Cai, J.F.; Gutierrez, H.R.; Song, L.K. Membrane Disruption Mechanism of a Prion Peptide (106–126) Investigated by Atomic Force Microscopy, Raman and Electron Paramagnetic Resonance Spectroscopy. J. Phys. Chem. B 2017, 121, 5058–5071. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gamage, Y.I.; Pan, J. Elucidating the Influence of Lipid Composition on Bilayer Perturbations Induced by the N-Terminal Region of the Huntingtin Protein. Biophysica 2023, 3, 582-597. https://doi.org/10.3390/biophysica3040040
Gamage YI, Pan J. Elucidating the Influence of Lipid Composition on Bilayer Perturbations Induced by the N-Terminal Region of the Huntingtin Protein. Biophysica. 2023; 3(4):582-597. https://doi.org/10.3390/biophysica3040040
Chicago/Turabian StyleGamage, Yasith Indigahawela, and Jianjun Pan. 2023. "Elucidating the Influence of Lipid Composition on Bilayer Perturbations Induced by the N-Terminal Region of the Huntingtin Protein" Biophysica 3, no. 4: 582-597. https://doi.org/10.3390/biophysica3040040