
Ostracod Assemblages in the East Siberian Sea: A Comparative Study of River-Influenced and River-Isolated Shelf Ecosystems
Abstract
:1. Introduction
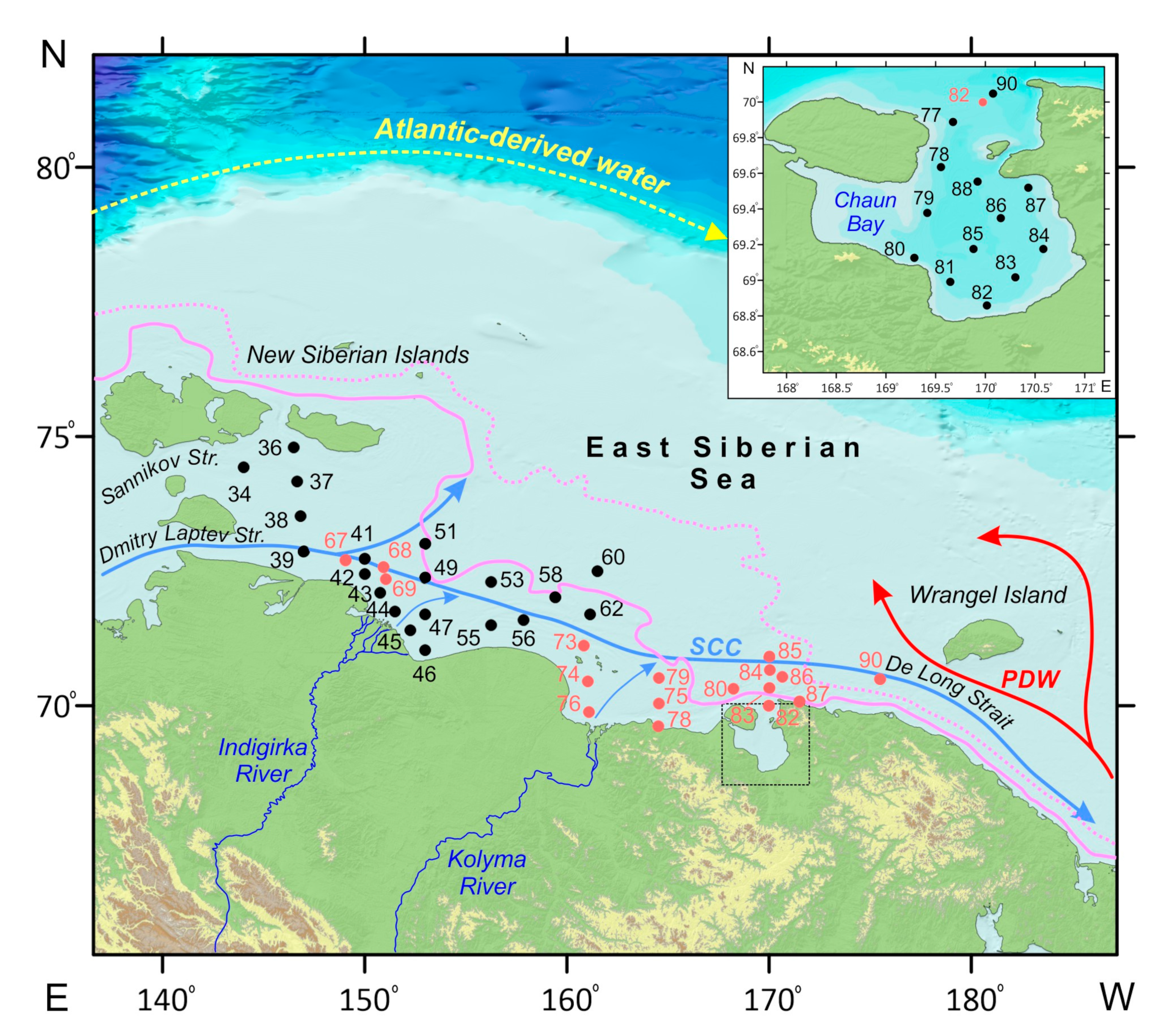
2. Regional Setting

3. Materials and Methods
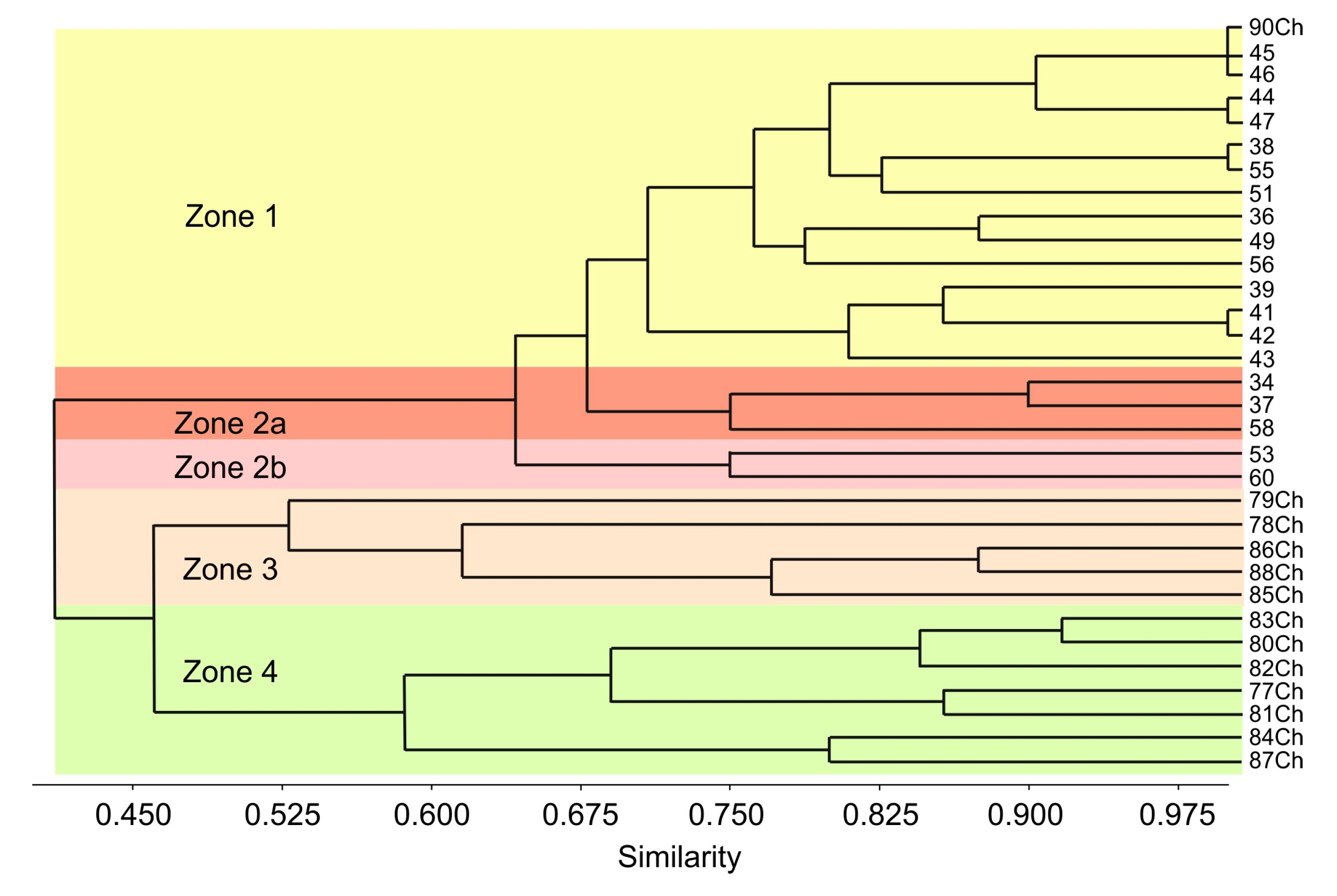
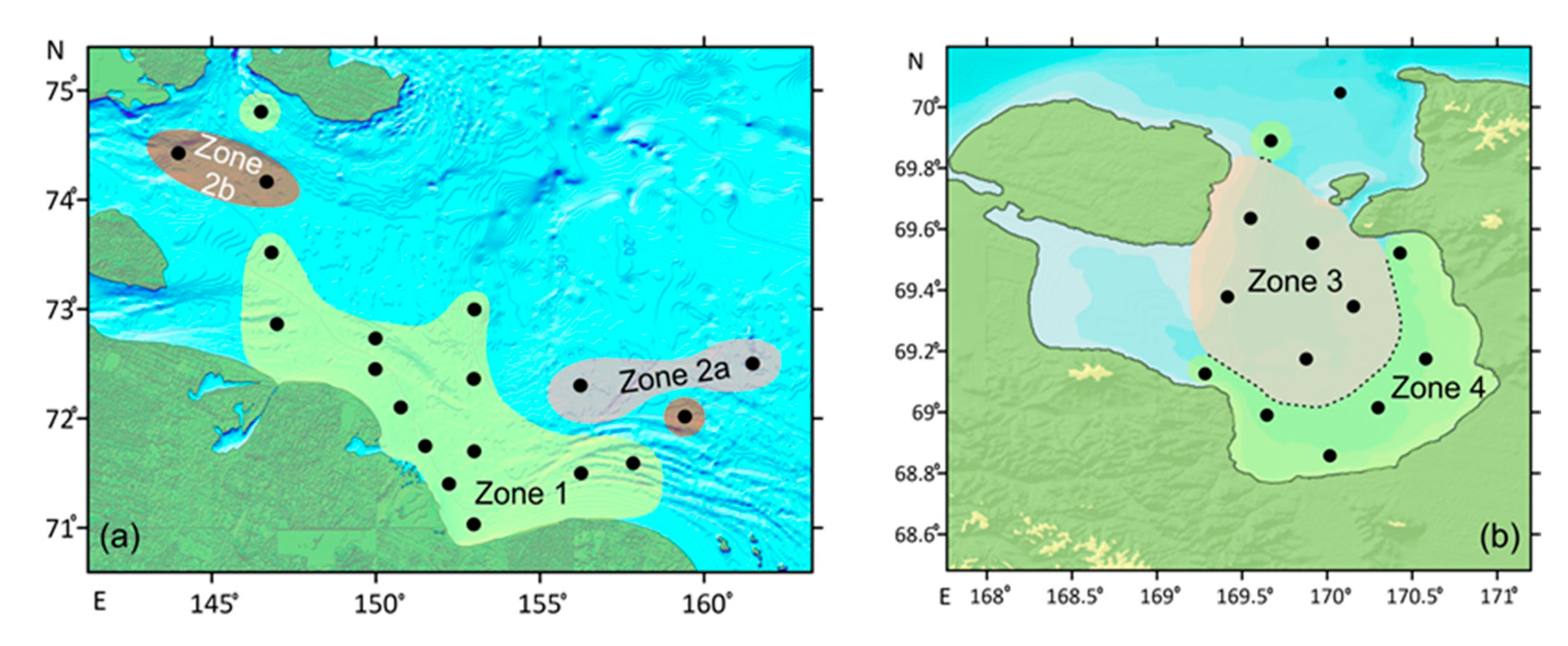
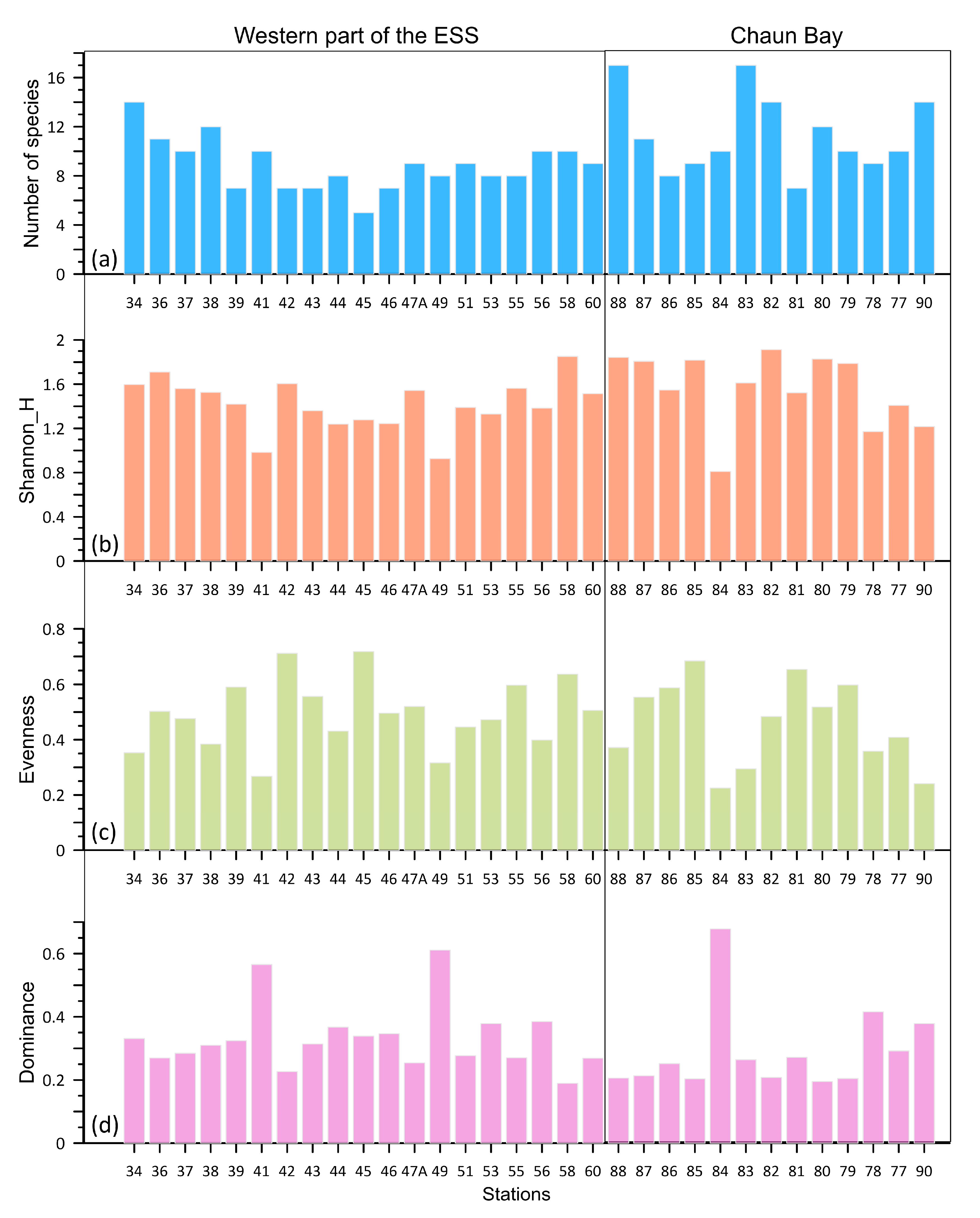
4. Results
4.1. The Area of the Indigirka River Mouth
4.2. The Chaun Bay
5. Discussion
5.1. A Comparison of the Ostracod Assemblages from the Western Part of the ESS and the Chaun Bay
5.2. Implications of Carbonate-Corrosive Environment
5.3. Brown Crusts and Spots on the Ostracod Valves from the Chaun Bay
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
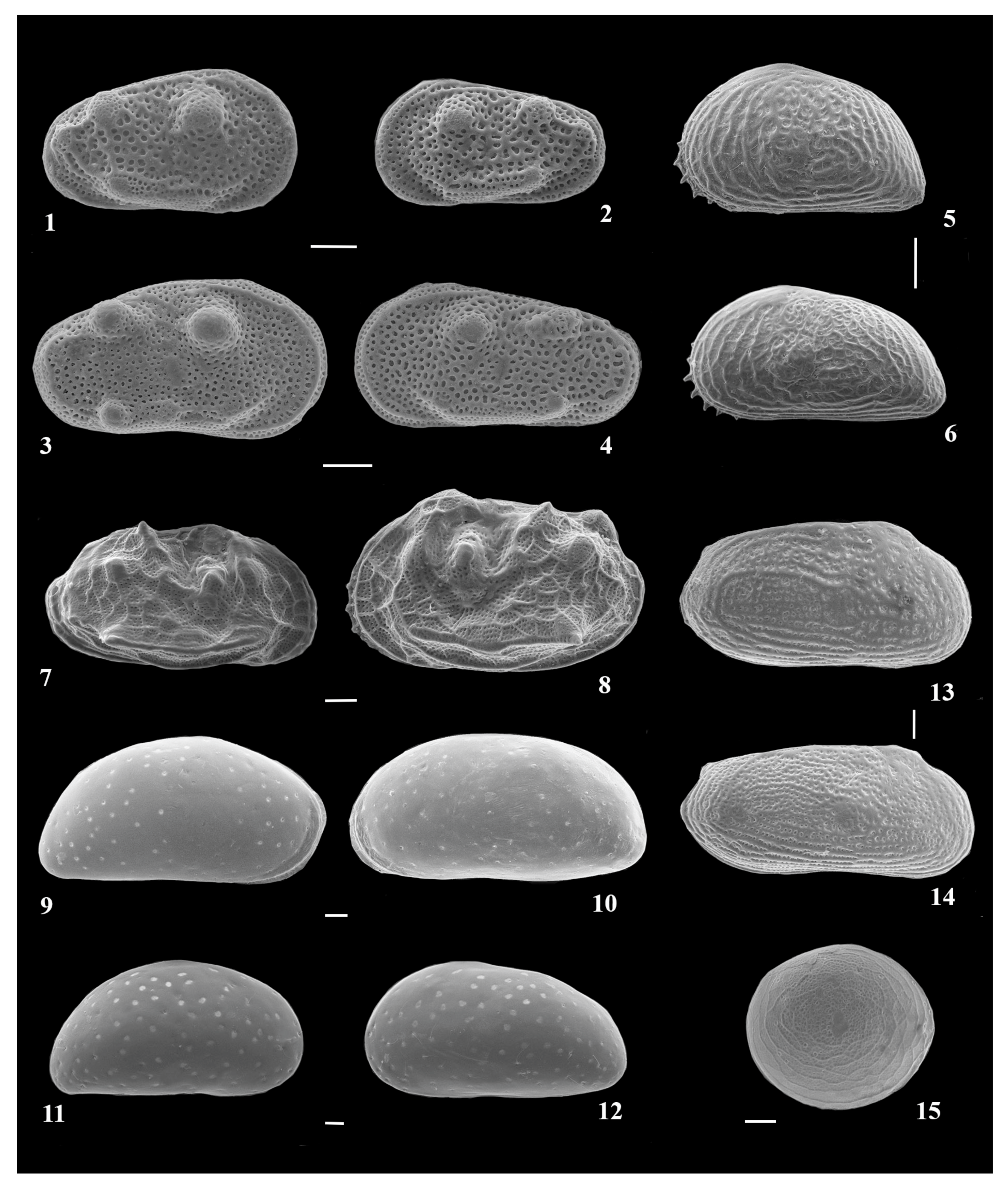{kind=link}
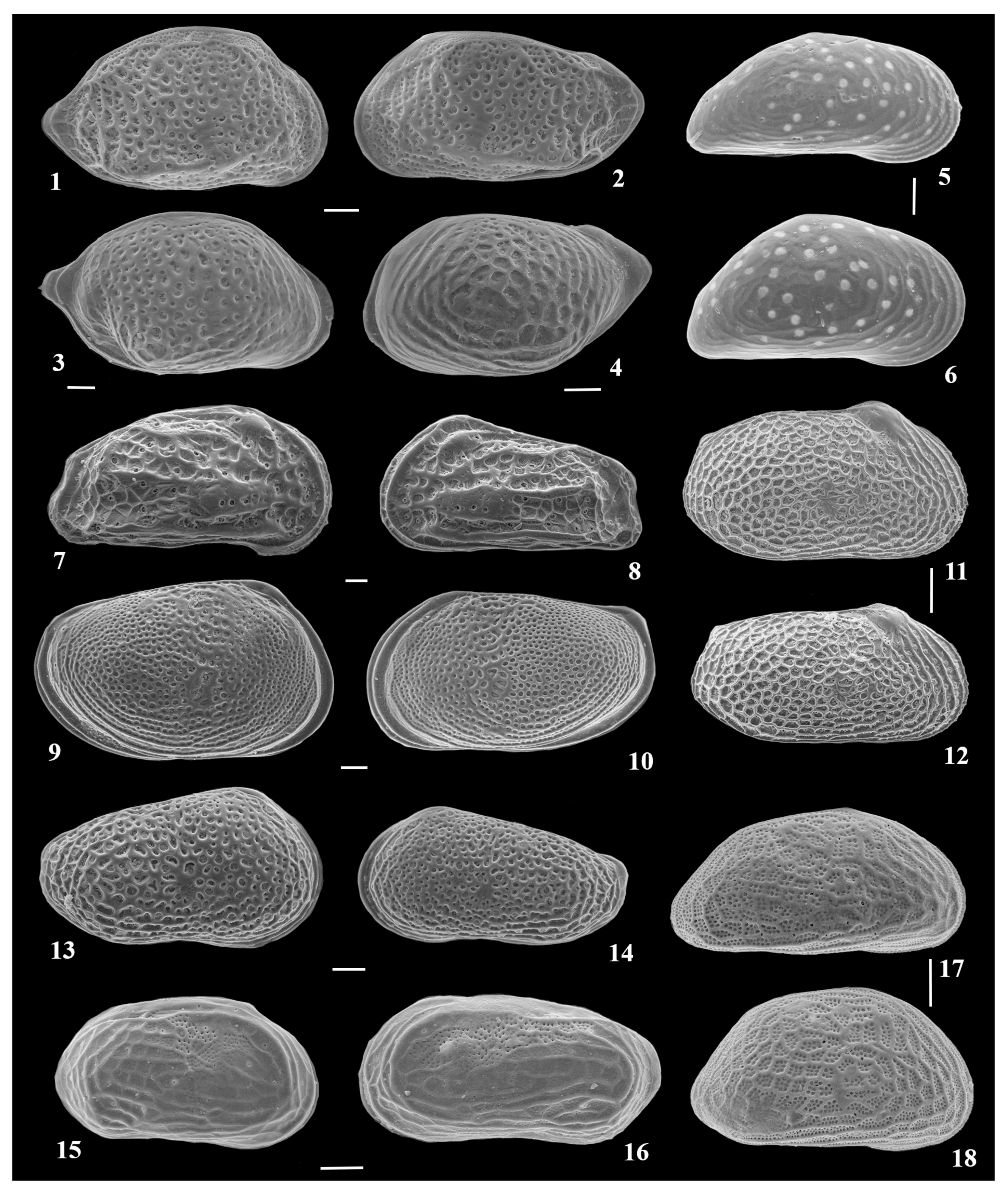{kind=link}
| № | Coordinates: N; E | Data | Depth (m) | Sediments | t °C | S‰ | O2, mL/L | O2% | pH |
|---|---|---|---|---|---|---|---|---|---|
| R/V «Nikolay Kolomeytsev» (NK), East Siberian Sea, 2000 | |||||||||
| 67 | 72°42.165′; 149°02.917′ | 06. 09. | 10 | sm, d | 2.9 | 21.98 | 7.95 | 97.6 | 7.78 |
| 68 | 72°34.1′; 150°56.8′ | 06. 09. | 15 | sm | 2.36 | 22.78 | 7.98 | 97.2 | 7.73 |
| 69 | 72°20.8′; 151°03.41′ | 06. 09. | 13 | sm | 2.68 | 22.58 | 7.96 | 97.6 | 7.74 |
| 73 | 71°07.218′; 160°49.803′ | 07. 09. | 9 | sm | 2.53 | 24.01 | 7.91 | 97.5 | 7.8 |
| 74 | 70°26.837′; 161°01.477′ | 08. 09. | 7 | sm | 2.87 | 25.25 | 7.76 | 97.4 | 7.81 |
| 75 | 70°02.397′; 164°33.753′ | 08. 09. | 20 | sm | 2.65 | 29.72 | 7.51 | 95.4 | 7.82 |
| 76 | 69°52.623′; 161°05.953′ | 08. 09. | 8 | sm, d | 2.46 | 26.69 | 7.83 | 98 | 7.76 |
| 78 | 69°37.105′; 164°31.745′ | 08. 09. | 10 | sm | 0.42° | 30.97 | 6.73 | 82.3 | 7.55 |
| 79 | 70°30.827′; 164°32.441′ | 09. 09. | 16 | sm | 1.89 | 28.97 | 7.73 | 96.9 | 7.9 |
| 80 | 70°18.99′; 168°14.125′ | 09. 09. | 34 | sm | 0.21° | 31.47 | 6.82 | 82.6 | 7.64 |
| 82 | 69°59.894′; 169°58.481′ | 09. 09. | 15 | ms | 0.37 | 31.69 | 6.98 | 85.6 | 7.66 |
| 83 | 70°19.583′; 170°00.134′ | 09. 09. | 27 | m, s | 0.47 | 31.23 | 7.42 | 90.9 | 7.85 |
| 84 | 70°39.842′; 170°01.530′ | 09. 09. | 25 | sm | 0.7 | 31.46 | 7.48 | 92.5 | 7.82 |
| 85 | 70°53.9′; 170°00.684′ | 09. 09. | 27 | sm | 0.33 | 31.89 | 6.71 | 80.9 | 7.73 |
| 86 | 70°32.310′; 170°39.587′ | 10. 09. | 27 | m, sm | −0.77 | 32.08 | 6.85 | 81.8 | 7.72 |
| 87 | 70°04.133′; 171°30.533′ | 10. 09. | 24 | st, s, m | 0.3 | 31.64 | 7.08 | 86.7 | 7.72 |
| 90 | 70°29.647′; 175°29.117′ | 10. 09. | 41 | m, sm | −1.76 | 33.36 | 5.96 | 69.9 | 7.58 |
| R/V “Ivan Kireev” (IK), the western part of the ESS, 2004 | |||||||||
| 34 | 74°25′50″; 144°00′08″ | 02.09. | 18 | m | 1.27 | 18.62 | 8.59 | 98.85 | 7.75 |
| 36 | 74°48′12″; 146°29′83″ | 02.09. | 14 | m | 1.46 | 19.23 | 8.62 | 100 | 7.79 |
| 37 | 74°09′83″; 146°40′25″ | 02.09. | 15 | ms | 0.89 | 17.98 | 8.79 | 99.73 | 7.76 |
| 38 | 73°30′98″; 146°50′23″ | 02.09. | 12 | ms | 1.90 | 17.63 | 8.54 | 99.25 | 7.76 |
| 39 | 72°51′78″; 146°59′85″ | 02.09. | 9 | m | 2.95 | 14.74 | 8.41 | 98.51 | 7.71 |
| 41 | 72°44.01′; 150°00.02′ | 02.09. | 15 | m | 1.8 | 16.69 | 8.48 | 97.70 | 7.67 |
| 42 | 72°27′01″; 149°59′94″ | 03.09. | 9 | m | 3.02 | 13.12 | 8.38 | 97.30 | 7.66 |
| 43 | 72°06′02″; 150°45′70″ | 03.09. | 7 | m | 3.17 | 12.84 | 8.29 | 96.41 | 7.64 |
| 44 | 71°45′01″; 151°30′61″ | 03.09. | 11 | m | 3.07 | 14.84 | 8.21 | 96.52 | 7,61 |
| 45 | 71°24′02″; 152°14′71″ | 03.09. | 11 | m | 3.58 | 12.45 | 7.99 | 93.69 | 7.47 |
| 46 | 71°02′01″; 152°59′98″ | 03.09. | 9 | m | 4.54 | 10.11 | 8.15 | 96.36 | 7.58 |
| 47 | 71°41′99″; 153°00′01″ | 03.09. | 15 | m | 0.99 | 17,88 | 8.34 | 94.89 | 7.40 |
| 49 | 72°22′00″; 152°59′84″ | 04.09. | 19 | m | −0.73 | 26.13 | 5.66 | 65.14 | 7.39 |
| 51 | 73°00′01″; 153°00′04″ | 04.09. | 22 | m | −1.07 | 29.41 | 3.13 | 36.56 | 7.23 |
| 53 | 72°18′02″; 156°15′44″ | 04.09. | 20 | m | −0.51 | 23.77 | 8.14 | 92.59 | 7.62 |
| 55 | 71°30′01″; 156°15′77″ | 04.09. | 12.5 | m | 2.89 | 16.80 | 8.20 | 97.24 | 7.68 |
| 56 | 71°35′38″; 157°50′68″ | 05.09. | 15.4 | m | 1.81 | 18.11 | 8.43 | 98.08 | 7.66 |
| 58 | 72°00′97″; 159°25′45″ | 05.09. | 21 | m | −0,41 | 22.82 | 8.42 | 95.45 | 7.62 |
| 60 | 72°30′08″; 161°30′09′ | 05.09. | 23 | m | −0,62 | 25.00 | 8.42 | 96.44 | 7,63 |
| 90 | 70°02.81′; 170°04.62′ | 08. 09. | 18 | m | −0.76 | 27.959 | 7.33 | 85.36 | 7.65 |
| R/V “Ivan Kireev” (IK), East Siberian Sea, Chaun Bay, 2004 | |||||||||
| 77 | 69°53.28′; 169°40.43′ | 07. 09. | 13 | sm | 3.89 | 20.458 | 7.88 | 7.74 | |
| 78 | 69°38.06′; 169°33.13′ | 07. 09. | 15 | sm | 3.89 | 20.019 | 7.69 | 95.64 | 7.73 |
| 79 | 69°22.69′; 169°24.95′ | 07. 09. | 14 | sm | 6.68 | 19.872 | 7.53 | 100.19 | 7.79 |
| 80 | 69°07.57′; 169°17.11′ | 07. 09. | 10 | s | 7.66 | 20.386 | 7.19 | 98.24 | 7.77 |
| 81 | 68°59.43′; 169°38.77′ | 07. 09. | 13 | sm, s, st, imn | 7.99 | 20.812 | 7.23 | 99.89 | 7.79 |
| 82 | 68°51.52′; 170°00.96′ | 07. 09. | 10.4 | s, st, c, imn | 8.56 | 19.56 | 7.26 | 100.77 | 7.79 |
| 83 | 69°01.02′; 170°18.06′ | 07. 09. | 13 | ms | 5.63 | 23.949 | 7.19 | 95.80 | 7.80 |
| 84 | 69°10.52′; 170°35.02′ | 07. 09. | 11 | m | 8.34 | 19.453 | 7.18 | 99.03 | 7.79 |
| 85 | 69°10.50′; 169°52.87′ | 07. 09. | 15 | sm | 1.52 | 25.047 | 7.56 | 91.56 | 7.44 |
| 86 | 69°20.90′; 170°09.37′ | 07. 09. | 15.4 | sm | 1.18 | 25.587 | 7.43 | 89.52 | 7.45 |
| 87 | 69°31.17′; 170°25.97′ | 07. 09. | 11 | m | 8.33 | 19.642 | 7.20 | 99.44 | 7.77 |
| 88 | 69°33.18′; 169°55.27′ | 08. 09. | 16 | m | 2.54 | 25.325 | 6.89 | 85.80 | 7.48 |
| Taxa | Station | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 34 | 36 | 37 | 38 | 39 | 41 | 42 | 43 | 44 | 45 | |
| Sarsicytheridea bradii | 182 17 | - - | 108 18 | 39 - | - - | 3 - | 2 - | 48 - | - - | - - |
| Semicytherura sp. 3 | 15 5 | 25 3 | - - | - - | - - | - - | - - | - - | - - | - - |
| Cluthia cluthae | 22 - | 5 - | 3 - | - - | - - | - - | - - | - - | - - | - - |
| Cluthia horni | 5 - | - - | - - | - - | - - | - - | - - | - - | - - | - - |
| Pteroloxa chaunensis | 13 4 | 31 8 | 1 - | 1 - | 2 - | 31 - | 37 - | 5 - | 85 12 | - 2 |
| Palmenella dentatomarginata | 1 2 | 21 4 | 29 2 | 9 2 | 1 - | - - | - - | - - | - - | - - |
| Cytheropteron occulatum | 2 2 | - 9 | 10 6 | - - | - - | 1 - | - - | - - | 2 - | 7 10 |
| Sclerochilus (S.) laptevensis | 2 2 | - - | - - | - - | - - | 1 1 | - - | - - | 2 - | - - |
| Pseudopolycope sp. | 11 - | - - | - - | 164 - | 12 - | 14 1 | 76 2 | 23 - | 12 - | - - |
| Roundstonia globulifera | 1 - | 39 2 | 5 - | 14 - | - - | 4 - | - - | - - | - - | 3 - |
| Eucythere sp. | 14 1 | 2 - | 21 2 | 1 - | - - | - - | - - | 2 - | - - | - - |
| Pontocythere sp. | 1 - | - - | 2 - | 1 - | - - | - - | - - | - - | - - | - - |
| Heterocyprideis sorbyana | 66 15 | 11 16 | 60 20 | 84 - | 8 - | 73 - | 88 2 | 153 - | 8 - | 7 38 |
| Paracyprideis sp. | 6 - | 225 31 | - - | 14 - | 2 - | 499 - | 66 - | 155 - | 231 1 | 25 24 |
| Cytheropteron sulense | - - | 82 56 | 2 - | 9 3 | 27 2 | 41 8 | 8 1 | - - | 91 6 | 40 67 |
| Cytheropteron montrosiense | - - | 38 12 | - - | 3 6 | - - | - - | - - | - - | 2 - | - 2 |
| Arctostoma dudarevi | - - | - - | - - | 1 - | - - | - - | - - | - - | - - | - - |
| Rabilimis septentrionalis | - - | 1 - | - - | - - | 1 - | 7 - | 15 - | 14 - | - - | - - |
| Total number: Living specimens Valves | 341 48 | 480 141 | 241 48 | 340 11 | 53 2 | 674 10 | 292 5 | 400 - | 433 - | 82 143 |
| Taxa | Station | |||||||||
| 46 | 47A | 47B | 49 | 51 | 53 | 55 | 56 | 58 | 60 | |
| Sarsicytheridea bradii | - - | - - | - - | - - | 314 - | - 2 | 7 - | 2 - | 34 2 | 54 2 |
| Semicytherura sp. 3 | - - | - - | - - | - - | - - | - - | - - | 1 - | - - | 1 - |
| Cluthia cluthae | - - | - - | - - | 18 - | 120 - | - - | - - | 21 - | 21 - | - - |
| Cluthia horni | - - | - - | - - | - - | - - | - - | - - | 9 - | 2 - | - - |
| Pteroloxa chaunensis | 10 - | 105 7 | 59 8 | 11 - | 2 - | 41 - | 18 1 | 18 1 | - - | 91 5 |
| Palmenella dentatomarginata | - - | - - | - - | - - | - - | - - | - - | - - | 1 - | - - |
| Cytheropteron occulatum | 1 - | 3 - | - 2 | - - | 1 - | 7 - | - - | - - | - - | 10 - |
| Sclerochilus (S.) laptevensis | - - | 1 - | - - | - - | - - | 4 - | - - | - - | - - | 6 2 |
| Pseudopolycope sp. | - - | 8 - | - - | 18 6 | 4 - | 4 3 | 25 - | 55 1 | 8 - | 18 - |
| Roundstonia globulifera | 4 2 | 20 - | - 1 | 6 - | 5 - | 2 - | 1 - | - - | 33 6 | 1 - |
| Eucythere sp. | - - | - - | - - | - - | - - | - - | - - | - - | 3 - | - - |
| Heterocyprideis sorbyana | 113 3 | 46 3 | 46 12 | 17 3 | 177 8 | - - | 5 - | 13 2 | 18 5 | - - |
| Paracyprideis sp. | 101 - | 174 - | 181 36 | 260 4 | 326 33 | 96 12 | 67 3 | 178 3 | 1 - | - - |
| Cytheropteron sulense | 32 3 | 109 16 | 10 6 | 1 2 | - - | 13 1 | 26 - | 4 2 | - - | 105 36 |
| Cytheropteron montrosiense | 2 - | 1 - | - - | 4 - | 1 - | - 2 | 1 - | 3 - | 5 6 | 3 - |
| Arctostoma dudarevi | - - | - - | - - | - - | - - | 4 - | - - | - - | - - | - - |
| Total number: Living specimens Valves | 263 8 | 467 26 | 296 65 | 335 15 | 950 41 | 171 20 | 150 4 | 304 9 | 126 19 | 289 45 |
| Taxa | Station | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 88 | 87 | 86 | 85 | 84 | 83 | 82 | 81 | 80 | 79 | |
| Sarsicytheridea bradii | 3264 176 | - - | 2516 335 | 24 6 | 4 - | 216 6 | 32 4 | 84 10 | - - | 107 4 |
| Semicytherura sp. 3 | - - | - - | - - | - - | - - | - - | - - | - - | - - | 1 - |
| Cluthia cluthae | 678 - | 5 - | - - | - - | - - | 13 6 | - - | - - | - - | - - |
| Cluthia horni | 1424 - | 224 - | 1 - | 36 - | - - | 236 33 | 4 - | - - | 18 - | 5 - |
| Pteroloxa chaunensis | 400 48 | 200 - | 13 - | 3 - | 15 1 | - - | - - | - - | - - | - - |
| Palmenella dentatomarginata | 2 - | 89 - | - - | - - | 4 - | 9 1 | 1 1 | - - | 1 1 | - - |
| Roundstonia globulifera | - - | 2 - | - - | - - | - - | 5 - | 67 - | 10 - | 138 - | - - |
| Eucythere sp. | - - | - - | - - | - - | 5 - | 18 - | 6 - | - - | 12 5 | 14 - |
| Pontocythere sp. | - - | - - | - - | - - | - - | 1 - | 10 - | - - | - - | - - |
| Heterocyprideis sorbyana | 112 2 | - - | 636 216 | 83 33 | - - | 4 3 | 6 - | 7 - | 4 1 | - 6 |
| Paracyprideis sp. | 1216 - | 1054 - | 826 418 | - - | 16 - | 13 - | 1 1 | 40 48 | - - | 146 3 |
| Arctostoma dudarevi | - - | - - | - - | - - | - - | - - | - - | - 2 | - - | - - |
| Rabilimis septentrionalis | 1 - | - - | 91 1106 | 26 13 | - - | - - | - - | - - | - - | - - |
| Acanthocythereis horrida | 94 50 | - - | - 38 | - - | - - | 3 - | - - | - - | - - | - - |
| Semicytherura complanata | 120 - | 513 22 | - - | - - | 528 - | 9 - | - - | - - | 1 - | 2 - |
| Munseyella kiklukhensis | 240 - | - - | 1087 48 | 26 - | - - | - - | - - | - - | - - | 68 - |
| Cytheropteron sp. cf. eleani | 41 5 | 88 6 | - - | - - | 42 5 | 283 - | - - | - - | - - | - - |
| Sarsicytheridea parapapillosa | - - | - - | 1008 133 | 7 2 | - - | 6 - | - - | 36 6 | 116 20 | 29 3 |
| Robertsonites tuberculatus | 1 - | - - | - - | - - | - - | - - | - - | - - | - - | 54 - |
| Kuiperiana venepidermoidea | - - | 154 - | - - | - - | 6 - | 3 - | 54 4 | - - | 21 - | - - |
| Semicytherura sp. 2. | - - | 220 - | - - | - - | 20 - | 10 4 | 11 - | 16 - | 77 16 | - - |
| Kotoracythere arctoborealis | 1584 32 | 1273 - | - - | - - | 4 - | - - | - - | - - | - - | - - |
| Jonesia arctica | - - | - - | - - | - - | - - | - - | - - | - - | - - | 25 - |
| Microcytherura nealei | - - | - - | - - | - - | - - | 1 - | 136 88 | - - | 13 - | - - |
| Cytheropteron sibiricum | - - | - - | - - | 1 - | - - | - - | 29 8 | - - | 1 - | - - |
| Cytheretta teshekpukensis | - - | - - | - - | - - | - - | 2 - | 8 - | - - | 94 - | - - |
| Sclerochilus semiletovi | - - | - - | - - | - - | - - | - - | - - | 1 - | - - | - - |
| Cytheropteron sp. | - - | - - | - - | - - | - - | - - | 1 - | - - | - - | - - |
| Finmarchinella (B.) logani | - - | - - | - - | 20 2 | - - | - - | - - | - - | - - | - - |
| Sclerochilus (S.) jurasovi | 1 - | - - | - - | - - | - - | - - | - - | - - | - - | - - |
| Heterocyprideis macrotuberculata | 3 - | - - | - - | - - | - - | - - | - - | - - | - - | - - |
| Cytheropteron pyramidale | 1 - | - - | - - | - - | - - | - - | - - | - - | - - | - - |
| Total number: Living specimens Valves | 9182 313 | 3862 28 | 6178 2294 | 226 56 | 644 6 | 832 53 | 366 106 | 194 67 | 496 43 | 451 16 |
| Taxa | Station | |||||||||
| 78 | 77 | 90 | ||||||||
| Sarsicytheridea bradii | 253 220 | 196 - | 1536 - | |||||||
| Cluthia cluthae | 4 8 | - - | 16 - | |||||||
| Cluthia horni | 64 224 | - 6 | - - | |||||||
| Pteroloxa chaunensis | 24 32 | 1676 - | 232 - | |||||||
| Cytheropteron occulatum | - - | - - | 3 1 | |||||||
| Sclerochilus (S.) laptevensis | - - | - - | 5 1 | |||||||
| Pseudopolycope sp. | 8 - | - - | - - | |||||||
| Roundstonia globulifera | - - | 32 - | 18 - | |||||||
| Heterocyprideis sorbyana | - 16 | 2930 - | 6 - | |||||||
| Paracyprideis sp. | 848 176 | 2084 - | 1552 - | |||||||
| Cytheropteron sulense | - - | - - | 51 5 | |||||||
| Cytheropteron montrosiense | - - | - - | 1 - | |||||||
| Argilloecia sp. | - - | - - | - 2 | |||||||
| Acanthocythereis horrida | - - | - - | 64 16 | |||||||
| Semicytherura complanata | - 352 | 304 - | 86 8 | |||||||
| Munseyella kiklukhensis | - - | - - | 2 0 | |||||||
| Cytheropteron sp. cf. eleani | 32 120 | - - | 3 2 | |||||||
| Sarsicytheridea parapapillosa | 127 68 | 110 - | - - | |||||||
| Robertsonites tuberculatus | - - | 1 - | - - | |||||||
| Kuiperiana venepidermoidea | - - | 8 - | - - | |||||||
| Semicytherura sp. 2. | - - | 32 - | - - | |||||||
| Kotoracythere arctoborealis | 1908 264 | - - | - - | |||||||
| Total number: Living specimens Valves | 3268 1480 | 7373 6 | 3575 34 | |||||||
| Taxa | Station | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 90 | 87 | 86 | 85 | 84 | 83 | 82 | 80 | 79 | 78 | |
| Sarsicytheridea bradii | - - | 1 4 | - - | - - | - - | 7 - | - - | 4 16 | - - | - - |
| Semicytherura sp. 3 | - - | - - | - - | - - | - - | - - | - - | 1 - | - - | - - |
| Cluthia cluthae | - - | 1 1 | - - | - - | - - | - - | - - | 1 - | - - | - - |
| Cluthia horni | - - | 2 4 | - - | - - | - - | - - | - - | 3 1 | - - | - - |
| Pteroloxa chaunensis | - - | 2 - | - - | 18 15 | - - | - - | - 1 | - - | - - | - - |
| Pontocythere sp. | - - | 3 4 | - - | 2 1 | - - | - - | - - | - - | - 1 | - - |
| Heterocyprideis sorbyana | - - | - - | 3 - | - - | - - | - 4 | - - | 3 3 | - - | 1 - |
| Paracyprideis sp. | - - | 5 6 | 2 1 | 2 1 | 1 - | - - | 5 - | 3 2 | - - | 12 22 |
| Cytheropteron sulense | - - | - - | - - | - - | - - | - - | 1 - | - - | - - | - - |
| Acanthocythereis horrida | - - | 3 3 | - 1 | - - | - - | - - | - - | - - | - - | - - |
| Cytheropteron sp. cf. eleani | - - | - - | - - | - 1 | - - | - - | - - | 2 - | - - | - - |
| Sarsicytheridea parapapillosa | - - | - 1 | 7 4 | - 3 | - - | - - | - - | - - | - 1 | - - |
| Robertsonites tuberculatus | 1 1 | - - | - - | - - | - - | - - | - - | - - | - - | - - |
| Heterocyprideis macrotuberculata | - - | - 4 | - - | - - | - - | 1 - | - - | 3 3 | - - | - - |
| Elofsonella neoconcinna | - - | - - | - - | - - | - - | - - | - - | 5 4 | - - | - - |
| Normanicythere leioderma | - - | - - | - - | - - | - - | - - | - - | 7 1 | - - | - - |
| Cytheropteron champlainum | - - | - 1 | - - | - - | - - | - - | - - | - - | - - | - - |
| Cytheropteron arcuatum | - - | - - | - - | - - | - - | - - | - - | 1 - | - - | - - |
| Jonesia sp. 1 | 2 - | - - | - - | - - | - - | - - | - - | - - | - - | - - |
| Cytheropteron suzdalskyi | 2 2 | - - | - - | - - | - - | - - | - - | - - | - - | - - |
| Total number: Living specimens Valve | 5 3 | 17 28 | - 12 6 | 22 21 | 1 0 | 8 4 | 6 1 | 33 30 | - 2 | 13 22 |
| Taxa | Station | |||||||||
| 76 | 75 | 74 | 73 | 69 | 68 | 67 | ||||
| Sarsicytheridea bradii | - - | - - | 14 10 | - - | - - | 1 1 | - - | |||
| Cluthia horni | - - | 1 - | - - | - - | - - | - - | - - | |||
| Pteroloxa chaunensis | - - | 2 10 | - - | - - | 1 - | 1 - | 3 - | |||
| Palmenella dentatomarginata | - 1 | 1 - | - - | - - | - - | - - | - - | |||
| Heterocyprideis sorbyana | - - | - - | 24 19 | - - | 2 - | 4 5 | - - | |||
| Paracyprideis sp. | 7 3 | 1 - | - - | - - | 24 14 | 13 15 | 3 - | |||
| Sarsicytheridea parapapillosa | - - | - 2 | - - | - - | 3 - | - - | - - | |||
| Sarsicytheridea macrolaminata | - - | - - | - - | - 1 | - - | - - | - - | |||
| Cytheropteron sulense | - - | - - | 1 1 | - - | 3 - | - - | 1 - | |||
| Total number: Living specimens Valve | 7 4 | 5 12 | 39 30 | - 1 | 33 14 | 19 21 | 7 - | |||
Appendix B


References
- Schornikov, E.I.; Zenina, M.A. Fauna of the benthic ostracods in the Kara, Laptev and East-Siberian seas (based on materials collected in the POI FEB RAS expeditions). In FEB RAS Marine Investigations in the Arctic; Sergienko, V.I., Semiletov, I.P., Eds.; Dalnauka: Vladivostok, Russia, 2006; pp. 156–211. (In Russian) [Google Scholar]
- Cronin, T.M.; Dowsett, H.J. A quantitative micropaleontologic method for shallow marine paleoclimatology: Application to Pliocene deposits of the Western North Atlantic Ocean. Mar. Micropaleontol. 1990, 16, 117–148. [Google Scholar] [CrossRef]
- Schornikov, E.I. Composition and distribution of the boreal-Arctic ostracod genus Paracyprideis Klie, 1929. In Modern Micropaleontology. Proceedings of the XVI All-Russian Micropaleontological Meeting (Kaliningrad); Nauka: Kaliningrad, Russia, 2015; pp. 107–110. [Google Scholar]
- Schornikov, E.I. Composition and distribution of the boreal-Arctic ostracod genus Heterocyprideis Elofson, 1941. In Modern Micropaleontology. Proceedings of the XVI All-Russian Micropaleontological Meeting (Kaliningrad); Nauka: Kaliningrad, Russia, 2015; pp. 111–115. [Google Scholar]
- Elofson, O. Zur Kenntnis der marinen Ostracoden Schwedens, mit besonderer berücksichtigung des Skageraks. Zool. Bidr. Från Upps. 1941, 19, 215–534. [Google Scholar]
- Akatova, N.A. Ostracoda. In Proceedings of the Glavesvmorput’ Drifting Expeditions on the Icebreaker Sedov, 1937–1940; Biologiya: Moscow-Leningrad, Russia, 1946; Volume 3, p. 11. (In Russian) [Google Scholar]
- Akatova, N.A. A Contribution to the Fauna of Ostracodes from the Shallow Waters of the New Siberian Islands. In Proceedings of the Glavesvmorput’ Drifting Expeditions on the Icebreaker Sedov, 1937–1940; Biologiya: Moscow-Leningrad, Russia, 1946; Volume 3, pp. 224–230. (In Russian) [Google Scholar]
- Chavtur, V.G. Class Ostracoda, Orders Myodocopida and Halocyprida. In List of Species of Free-Living Invertebrates of Eurasian Arctic Seas and Adjacent Deep Waters. Explorations of the Fauna of the Seas; Sirenko, B.I., Ed.; ZIN RAS: Saint Petersburg, Russia, 2001; Volume 51, pp. 98–99. [Google Scholar]
- Cronin, T.M.; Briggs, W.M., Jr.; Brouwers, E.M.; Whatley, R.C.; Wood, A.; Cotton, M.A. Modern Arctic Podocopid ostracod database. In US Geological Survey Open File Report; US Geological Survey: Reston, VI, USA, 1991; pp. 91–385. [Google Scholar]
- Cronin, T.M.; Gemery, L.; Briggs, W.M., Jr.; Brouwers, E.M.; Schornikov, E.I.; Stepanova, A.; Wood, A.; Yasuhara, M.; Siu, S.; Arctic Ostracode Database 2020. NOAA’s National Centers for Environmental Information (NCEI). 2021. Available online: https://www.ncdc.noaa.gov/paleo/study/32312 (accessed on 1 January 2024).
- Gemery, L.; Cronin, T.M.; Briggs, W.M.; Brouwers, E.M.; Schornikov, E.; Stepanova, A.; Wood, A.M.; Yasuhara, M. An Arctic and subarctic ostracode database: Biogeographic and paleoceanographic applications. Hydrobiologia 2015, 786, 59–95. [Google Scholar] [CrossRef]
- Stepanova, A.; Taldenkova, E.; Simstich, J.; Bauch, H. Comparison study of the modem ostracod associations in the Kara and Laptev seas: Ecological aspects. Mar. Micropaleontol. 2007, 63, 111–142. [Google Scholar] [CrossRef]
- Stepanova, A.; Taldenkova, E.; Bauch, H.A. Ostracod palaeoecology and environmental change in the Laptev and Kara seas (Siberian Arctic) during the last 18,000 years. Boreas 2012, 41, 557–577. [Google Scholar] [CrossRef]
- Stepanova, A.; Taldenkova, E.; Bauch, H.A. Recent Ostracoda from the Laptev Sea (Arctic Siberia): Species assemblages and some environmental relationships. Mar. Micropaleontol. 2003, 48, 23–48. [Google Scholar] [CrossRef]
- Stepanova, A.Y. Late Pleistocene-Holocene and Recent Ostracoda of the Laptev Sea and their importance for paleoenvironmental reconstructions. Paleontol. J. 2006, 40, S91–S204. [Google Scholar] [CrossRef]
- Gemery, L.; Cronin, T.M.; Cooper, L.W.; Dowsett, H.J.; Grebmeier, J.M. Biogeography and ecology of Ostracoda in the U.S. northern Bering, Chukchi, and Beaufort Seas. PLoS ONE 2021, 16, e0251164. [Google Scholar] [CrossRef]
- Gemery, L.; Cooper, L.W.; Magen, C.; Cronin, T.M.; Grebmeier, J.M. Stable oxygen isotopes in shallow marine ostracodes from the northern Bering and Chukchi Seas. Mar. Micropaleontol. 2022, 174, 102001. [Google Scholar] [CrossRef]
- Jakobsson, M. Hypsometry and Volume of the Arctic Ocean and Its Constituent Seas. Geochem. Geophys. Geosyst. 2002, 3, 1–18. [Google Scholar] [CrossRef]
- Gordeev, V.V.; Martin, J.M.; Sidorov, I.S.; Sidorova, M.V. A reassessment of the Eurasian river input of water, sediment, major elements, and nutrients to the Arctic Ocean. Am. J. Sci. 1996, 296, 664–691. [Google Scholar] [CrossRef]
- Semiletov, I.P. The failure of coastal frozen rock as an important factor in the biogeochemistry of the arctic shelf water. Dokl. Earth Sci. 1999, 369, 1140–1143. [Google Scholar]
- Dmitrenko, I.; Kirillov, S.; Eicken, H.; Markova, N. Wind-Driven Summer Surface Hydrography of the Eastern Siberian Shelf. Geophys. Res. Lett. 2005, 32, L14613. [Google Scholar] [CrossRef]
- Semiletov, I. The East Siberian Sea as a Transition Zone between Pacific-Derived Waters and Arctic Shelf Waters. Geophys. Res. Lett. 2005, 32, L10614. [Google Scholar] [CrossRef]
- Yulin, A.V.; Sharatunova, M.V.; Pavlova, E.A.; Ivanov, V.V. Seasonal and interannual changes of ice massifs in East Siberian sea. Arct. Antarct. Res. 2018, 64, 229–240. [Google Scholar] [CrossRef]
- Nürnberg, D.; Wollenburg, I.; Dethleff, D.; Eicken, H.; Kassens, H.; Letzig, T.; Reimnitz, E.; Thiede, J. Sediments in Arctic Sea Ice: Implications for Entrainment, Transport and Release. Mar. Geol. 1994, 119, 185–214. [Google Scholar] [CrossRef]
- Solovyev, B.; Spiridonov, V.; Onufrenya, I.; Belikov, S.; Chernova, N.; Dobrynin, D.; Gavrilo, M.; Glazov, D.; Krasnov, Y.; Mukharamova, S.; et al. Identifying a Network of Priority Areas for Conservation in the Arctic Seas: Practical Lessons from Russia. Aquat. Conserv. Mar. Freshw. Ecosyst. 2017, 27, 30–51. [Google Scholar] [CrossRef]
- Osadchiev, A.; Frey, D.; Spivak, E.; Shchuka, S.; Tilinina, N.; Semiletov, I. Structure and Inter-Annual Variability of the Freshened Surface Layer in the Laptev and East-Siberian Seas During Ice-Free Periods. Front. Mar. Sci. 2021, 8, 735011. [Google Scholar] [CrossRef]
- Weingartner, T.J.; Danielson, S.; Sasaki, Y.; Pavlov, V.; Kulakov, M. The Siberian Coastal current: A wind and buoyancy forced coastal current. J. Geophys. Res. 1999, 104, 29697–29713. [Google Scholar] [CrossRef]
- Romanovskii, N.N.; Hubberten, H.-W.; Gavrilov, A.V.; Eliseeva, A.A.; Tipenko, G.S. Offshore Permafrost and Gas Hydrate Stability Zone on the Shelf of East Siberian Seas. Geo-Mar. Lett. 2005, 25, 167–182. [Google Scholar] [CrossRef]
- Schirrmeister, L.; Kunitsky, V.; Grosse, G.; Wetterich, S.; Meyer, H.; Schwamborn, G.; Babiy, O.; Derevyagin, A.; Siegert, C. Sedimentary Characteristics and Origin of the Late Pleistocene Ice Complex on North-East Siberian Arctic Coastal Lowlands and Islands—A Review. Quat. Int. 2011, 241, 3–25. [Google Scholar] [CrossRef]
- Anderson, L.G.; Björk, G.; Jutterström, S.; Pipko, I.; Shakhova, N.; Semiletov, I.P.; Wåhlström, I. East Siberian Sea, an Arctic Region of Very High Biogeochemical Activity. Biogeochemistry 2011, 8, 1745–1754. [Google Scholar] [CrossRef]
- Semiletov, I.; Pipko, I.; Gustafsson, Ö.; Anderson, L.G.; Sergienko, V.; Pugach, S.; Dudarev, O.; Charkin, A.; Gukov, A.; Bröder, L.; et al. Acidification of East Siberian Arctic Shelf Waters through Addition of Freshwater and Terrestrial Carbon. Nat. Geosci. 2016, 9, 361–365. [Google Scholar] [CrossRef]
- Andersson, A.J.; Mackenzie, F.T.; Lerman, A. Coastal ocean CO2-carbonic acid-carbonate sediment system of the Anthropocene. Glob. Biogeochem. Cycles 2006, 20, GB1S92. [Google Scholar] [CrossRef]
- Bertin, C.; Carroll, D.; Menemenlis, D.; Dutkiewicz, S.; Zhang, H.; Matsuoka, A.; Tank, S.; Manizza, M.; Miller, C.E.; Babin, M.; et al. Biogeochemical river runoff drives intense coastal Arctic Ocean CO2 outgassing. Geophys. Res. Lett. 2023, 50, e2022GL102377. [Google Scholar] [CrossRef]
- Romankevich, E.A.; Vetrov, A.A. Carbon in the World Ocean; GEOS: Moscow, Russia, 2021; p. 352. (In Russian) [Google Scholar]
- Babkov, A.I. Hydrological characteristics of the East-Siberian Sea. In Ecosystems, Flora and Fauna of the Chaunskaya Bay and Adjacent Waters of the East-Siberian Sea; ZIN RAS: Saint Petersburg, Russia, 1994; Volume 2, pp. 4–16. (In Russian) [Google Scholar]
- Golikov, A.N.; Gagaev, S.Y.; Galtsova, V.V.; Golikov, A.A.; Danton, K.; Menshutkina, T.V.; Novikov, O.K.; Petryashev, V.V.; Potin, V.V.; Sirenko, B.I.; et al. Ecosystems, flora and fauna of the Chaun Bay of the East-Siberian Sea. In Ecosystems, Flora and Fauna of the Chaun Bay of the East-Siberian Sea; ZIN RAN: Saint Petersburg, Russia, 1994; Volume 47, pp. 4–111. (In Russian) [Google Scholar]
- Schönfeld, J.; Alve, E.; Geslin, E.; Jorissen, F.; Korsun, S.; Spezzaferri, S. The FOBIMO (FOraminiferal BIo-MOnitoring) initiative—Towards a standardized protocol for soft-bottom benthic foraminiferal monitoring studies. Mar. Micropal. 2012, 94–95, 1–13. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological Statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Tishchenko, P.Y.; Wong, C.; Pavlova, G.Y.; Johnson, W.K.; Kang, D.J.; Kim, K.R. The Measurement of pH Values in Seawater Using a Cell without a Liquid Junction. Oceanology 2001, 41, 813–822. [Google Scholar]
- Schornikov, E.I. Class Ostracoda, Orders Platycopida and Podocopida. List of species of free-living invertebrates of Eurasian Arctic seas and adjacent deep waters. In Explorations of the Fauna of the Seas; Sirenko, B.I., Ed.; ZIN RAS: Saint Petersburg, Russia, 2001; Volume 51, pp. 99–103. [Google Scholar]
- Schornikov, E.I. Class Ostracoda. Order Podocopida. In Fauna of the East-Siberian Sea, Regularities of Development and Quantitative Distribution of Bottom Ecosystems. Appendix 2. List of Species of Invertebrates of the East-Siberian Sea. Explorations of the Fauna of the Seas; Sirenko, B.I., Ed.; ZIN RAS: Saint Petersburg, Russia, 2010; Volume 66, pp. 265–272. (In Russian) [Google Scholar]
- Kagan, B.A.; Timofeev, A.A. High-Resolving modeling of the surface semidiurnal M2 tide in the East-Siberian Sea. Fundam. Appl. Hydrophys. 2023, 16, 64–72. [Google Scholar] [CrossRef]
- Charkin, A.N.; Yaroshchuk, E.I.; Dudarev, O.V.; Leusov, A.E.; Goriachev, V.A.; Sobolev, I.S.; Gulenko, T.A.; Pipko, I.I.; Startsev, A.M.; Semiletov, I.P. The Influence of Sedimentation Regime on Natural Radionuclide Activity Concentration in Marine Sediments of the East Siberian Arctic Shelf. J. Environ. Radioact. 2022, 253–254, 106988. [Google Scholar] [CrossRef]
- Ovsepyan, E.A.; Ovsepyan, Y.S.; Zenina, M.A.; Mitrofanova, N.O. Recent benthic foraminiferal assemblages in the western part of the East Siberian Sea. Oceanology 2023, 63, 651–663. [Google Scholar] [CrossRef]
- Grigoriev, M.N.; Razumov, S.O.; Kunitzkiy, V.V.; Spektor, V.B. Dynamics of the Russian East Arctic Sea coast: Major factors, regularities and tendencies. Earth’s Cryosphere 2006, 10, 74–94. (In Russian) [Google Scholar]
- Pipko, I.I.; Semiletov, I.P.; Pugach, S.P.; Wåhlström, I.; Anderson, L.G. Interannual Variability of Air-Sea CO2 Fluxes and Carbon System in the East Siberian Sea. Biogeosciences 2011, 8, 1987–2007. [Google Scholar] [CrossRef]
- Shakhova, N.; Semiletov, I.; Salyuk, A.; Yusupov, V.; Kosmach, D.; Gustafsson, Ö. Extensive Methane Venting to the Atmosphere from Sediments of the East Siberian Arctic Shelf. Science 2010, 327, 1246–1250. [Google Scholar] [CrossRef] [PubMed]
- Shur, Y.; Fortier, D.; Jorgenson, M.T.; Kanevskiy, M.; Schirrmeister, L.; Strauss, J.; Vasiliev, A.; Ward Jones, M. Yedoma Permafrost Genesis: Over 150 Years of Mystery and Controversy. Front. Earth Sci. 2022, 9, 757891. [Google Scholar] [CrossRef]
- Boomer, I.; Whatley, R.S.; Keyser, D. Metallic and organic deposits on carapaces of living Cyprideis torosa (Ostracoda) in disaerobic environments. In Proceedings of the 2nd European Ostracodologists Meeting, Glasgow, UK, 23–27 July 1993; pp. 163–170. [Google Scholar]
- Ulyantsev, A.S.; Charkin, A.N.; Syomin, V.L.; Kireenko, L.A.; Botsul, S.A.; Leusov, A.E.; Semkin, P.Y.; Kukla, S.P. Geological Studies of the Upper Sedimentary Strata of Chaun Bay during Cruise 60 of the R/V Akademik Oparin. Oceanology 2021, 61, 584–585. [Google Scholar] [CrossRef]
- Shulga, N.; Abramov, S.; Klyukina, A.; Ryazantsev, K.; Gavrilov, S. Fast-Growing Arctic Fe–Mn Deposits from the Kara Sea as the Refuges for Cosmopolitan Marine Microorganisms. Sci. Rep. 2022, 12, 21967. [Google Scholar] [CrossRef]
- Whatley, R.C.; Moguilevsky, A. Nota preliminar acerca de la presencia de Diatomeas sobre el Caparazon de Callistocythere asperenticulata Whatley y Moguilevsky (Crustacea, Ostracoda). Physis Ser. A 1973, 32, 489–493. [Google Scholar]



| Taxa | Mention for the ESS | Western Part of the ESS | Chaun Bay * |
|---|---|---|---|
| Philomedes brenda (Baird, 1850) | [1,6,8,40] | ||
| Scleroconcha ruffi Kornicker, 1988 | [40] | ||
| Polycope sp. s. Akatova, 1946 | [40] | ||
| Pseudopolycope sp. B Schornikov and Zenina, 2006 | [1] | ■ | |
| Argilloecia sp. s. Schornikov, 2001 | [1,40] | ||
| Jonesia acuminata (Sars, 1866) | [1,40,41] | ||
| Jonesia arctica Schornikov, 1980 | [1,40] | ■ | |
| Jonesia sp.1 s. Schornikov, 2001 | [1,40] | ||
| Sclerochilus (S.) jurasovi Schornikov, 2004 | [1,40] | ■ | |
| S. (S.) laptevensis Schornikov, 2004 | [1,40] | ■ | |
| S. (S.) semiletovi Schornikov and Zenina, 2006 | [1,40] | ■ | |
| Microcytherura nealei Schornikov and Zenina, 2006 | [1,40] | ■ | |
| Palmenella dentatomarginata Akatova, 1946 | [1,40] | ■ | ■ |
| Cluthia cluthae (Brady, Crosskey and Robertson, 1874) | [1,40] | ■ | ■ |
| C. horni Schornikov and Zenina, 2006 | [1,40] | ■ | ■ |
| Eucythere sp. 1. s. Schornikov, 2004 | [1,40] | ■ | |
| Heterocyprideis fascis (Brady and Norman, 1889) | [1,40] | ■ | ■ |
| H. sorbyana (Jones, 1856) | [1,6,9,40,41] | ■ | ■ |
| Paracyprideis sp. | [1,3,6,9,40,41] | ■ | ■ |
| Sarsicytheridea bradii (Norman in Brady, 1865) | [1,6,9,40,41] | ■ | ■ |
| S. macrolaminata (Elofson, 1939) | [1,40] | ||
| S. parapapillosa (Swain, 1963) | [1,40] | ■ | ■ |
| S. punctillata (Brady, 1865) | [6,9,40] | ||
| Pontocythere sp. s. Schornikov, 2004 | [1,40] | ■ | ■ |
| Krithe glacialis Brady, Crosskey and Robertson, 1874 | [1,6,9,40] | ||
| Acanthocythereis horrida (Sars, 1866) | [1,40] | ■ | |
| A. dunelmensis (Norman, 1865) | [1], [3]?, [6]?, [9]?, [40] | ■ | |
| Robertsonites tuberculatus (Sars, 1866) | [1,6,40] | ■ | |
| Cytheretta teshekpukensis Swain, 1963 | [1,40] | ■ | |
| Finmarchinella (Barentsovia) barenzovoensis (Mandelstam, 1957) | [6,36,40] | ||
| F. (B.) logani (Brady and Crosskey, 1871) | [1,40] | ■ | |
| Elofsonella pinegensis (Lev, 1983) | [1,40,41] | ||
| Rabilimis mirabilis (Brady, 1868) | [1,6,40,41] | ||
| R. septentrionalis (Brady, 1866) | [1,40] | ■ | ■ |
| Kuiperiana venepidermoidea (Swain, 1963) | [1,40] | ■ | |
| Pteroloxa chaunensis Schornikov and Zenina, 2006 | [1,40] | ■ | ■ |
| Roundstonia globulifera (Brady, 1868) | [1,40] | ■ | ■ |
| Semicytherura complanata (Brady, Crosskey and Robertson, 1874) | [1,40] | ■ | |
| Semicytherura sp. 2 s. Schornikov, 2001 | [1,40] | ■ | |
| Semicytherura sp. 3 s. Schornikov and Zenina, 2006 | [1,40] | ■ | ■ |
| Kotoracythere arctoborealis Schornikov and Zenina, 2006 | [1,40] | ■ | |
| Munseyella kiklukhensis (Brouwers, 1990) | [40] | ■ | |
| Cytheropteron arcuatum Brady, Crosskey and Robertson, 1874 | [1,9,40] | ||
| C. champlainum Cronin, 1981 | [1,40] | ||
| C. cf. elaeni Cronin, 1989 s. Schornikov, 2004 | [1,40] | ■ | |
| C. montrosiense Brady, Crosskey and Robertson, 1874 | [1,9,40,41] | ■ | |
| C. occulatum Whatley and Masson, 1979 | [1,40] | ■ | |
| ?C. paralatissimum Swain, 1963 | [6,40,41] | ||
| C. pseudomontrosiense Whatley and Masson, 1979 | [1,9,40] | ||
| C. pyramidale Brady, 1868 | [1,40] | ■ | |
| C. sibiricum Schornikov and Zenina, 2006 | [1,40] | ■ | |
| C. sulense Lev, 1972 | [1,40] | ■ | |
| C. suzdalskyi Lev, 1972 | [1,40] | ||
| Cytheropteron sp. s. Schornikov and Zenina, 2006 | [1,40] | ■ | |
| Arctostoma dudarevi Schornikov and Zenina, 2006 | [1,40] | ■ | □ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zenina, M.; Ovsepyan, E.; Ovsepyan, Y. Ostracod Assemblages in the East Siberian Sea: A Comparative Study of River-Influenced and River-Isolated Shelf Ecosystems. Quaternary 2024, 7, 16. https://doi.org/10.3390/quat7010016
Zenina M, Ovsepyan E, Ovsepyan Y. Ostracod Assemblages in the East Siberian Sea: A Comparative Study of River-Influenced and River-Isolated Shelf Ecosystems. Quaternary. 2024; 7(1):16. https://doi.org/10.3390/quat7010016
Chicago/Turabian StyleZenina, Maria, Ekaterina Ovsepyan, and Yaroslav Ovsepyan. 2024. "Ostracod Assemblages in the East Siberian Sea: A Comparative Study of River-Influenced and River-Isolated Shelf Ecosystems" Quaternary 7, no. 1: 16. https://doi.org/10.3390/quat7010016