1. Introduction
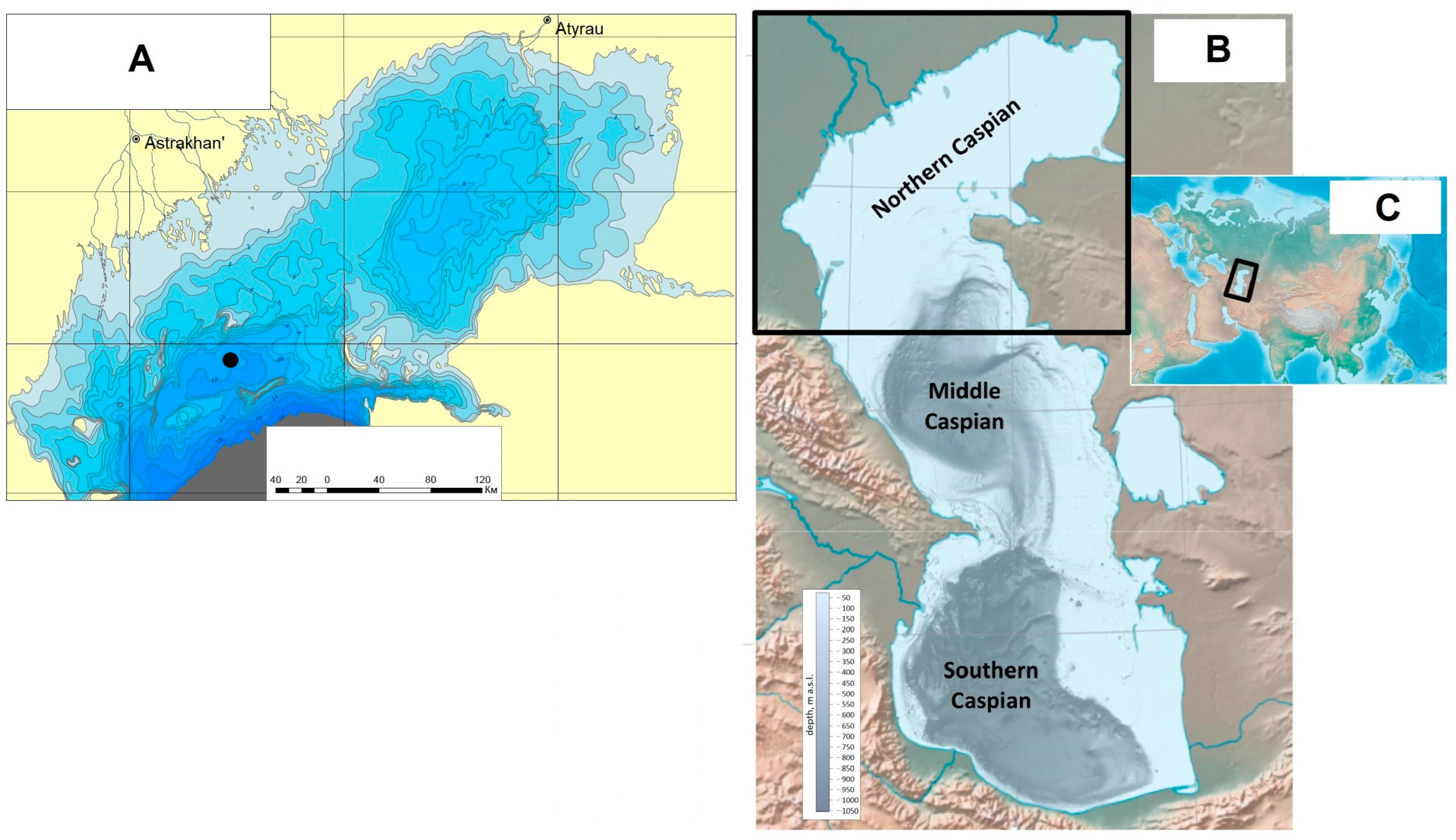
The Caspian Sea is the largest lake in the world, distinguished by the uniqueness of its paleogeographic evolution. The three parts (northern, middle, and southern), separated by the Mangyshlak and Apsheron Threshholds, have clearly different physical-geographic features, relief, and hydrological regimes (
Figure 1). The research presented herein considers the evolution of the Northern Caspian Sea environment. The area of the Northern Caspian basin is about 95,000 km
2, with the most common depth of 5 m. This is a gently undulating accumulative plain with sandbanks and islands, as well as depressions and linear hollows, which were the channels of the Volga and Ural Rivers at the time of a low Caspian Sea level. The dominant type of the bottom sediment is coarse silt. Medium- and fine-grained sands occur on shallow banks. The surface of the largest banks is often composed of broken shells (coquina).
The Northern Caspian basin is located in the zone of continental moderate climate. This part of the sea receives 88% of the total flow of river freshwater (Volga, Ural, Terek). The average annual salinity of the Northern Caspian Sea ranges from 6.4 to 11.7‰. In the mouth of the Volga River, salinity is 1–3‰. The water temperature varies from 27° in summer to 0 in winter. The ice cover develops annually.
Long-term water level fluctuations play a large role in the hydrological regime of the Caspian Sea. Since the beginning of instrumental observations in 1830, the amplitude of oscillations is 4 m (from −25.3 m in the 1880s to −29 m in 1977). In 1978, the Caspian Sea level began to rise, going up by more than 2 m in 16 years. The year 1995 marks the level stabilization and fall. The current Caspian Sea level is −28 m below the global sea level.
Paleogeographical data suggest significant changes in the Caspian level in the Pleistocene. Caspian Sea level depends upon many factors, including climate, state of the hydrographic network, volume of surface and underground flow, precipitation, tectonic and seismic activity of the basin, and other factors [
1,
2,
3,
4,
5,
6,
7]. The intensity of the influence of these factors varied through the Caspian geological history. During the Khazarian transgression, corresponding to marine isotopic stages (MIS) 7–5, climate change played the defining role [
4,
5,
7].
MIS 7 (243–191 ka) covers the last interglacial epoch of the Middle Pleistocene (Cherepet’, Romny, Schöningen epochs), with two peaks separated by endothermic cooling [
8]. Two broadleaf pollen maxima corresponding to the optimal phases of the thermoxerotic and thermohygrotic stages of this interglacial are separated by a palynozone with a significant reduction in the pollen content of thermophilic dendroflora taxa. It is followed by a complex glacial phase (Dnieper, Saale), corresponding to MIS 6 (191–130 ka) at the very end of the Middle Pleistocene. Three interstadial stages were identified there, with the middle one broken into two periods (i.e., Dnieper and Moscow) [
8,
9]. The beginning of the Late Pleistocene (MIS 5) is marked by an interglacial phase (Mikulino, Eemian). The geochronological position of the interglacial phase is accepted by the International Stratigraphic Commission in MIS 5e [
10,
11] (estimated 128–115 ka). Considering the fact that a number of researchers argue with such a position in the view of their own data and suggest that the interglacial phase covers the entire MIS 5 or its greater part [
12,
13,
14,
15], in the work presented in [
12] it was proposed to isolate the Eemian interglacial sensu stricto (s.s.) (a warm period, manifested in sections of western Europe) and the Eemian interglacial sensu lato (s.l.) (a period of thermophilic forest vegetation in southwestern and southern Europe). Researchers who accept that the Eemian (Mikulino) interglacial epoch corresponds to MIS 5e refer to a complex climatic interval corresponding to MIS 5d–a to the Weichselian (Valdai) glacial epoch.
The history of the Caspian Sea development and the evolution of its natural environment are covered in numerous studies [
3,
4,
5,
6,
16,
17,
18,
19,
20,
21]. They mainly focus on the reconstruction of paleogeographic events (transgression or regression, the position of the basin level) and environment conditions derived from faunal analysis. Most of the publications result from studies of Pleistocene sediments in exposed sections in the coastal zone of the Caspian Sea. The works based on marine drilling materials are scarce. Primarily, our engineering-geological survey studies of the borehole cores from the Upper Pleistocene strata in the Northern Caspian Sea [
22,
23].
Most Caspian researchers accept the fact that in the second half of the Middle Pleistocene, the Early Khazarian transgression developed and reached a level of 20–30 m [
3,
4,
5,
6]. Some experts [
5,
6] reconstruct three stages there. The beginning of the Late Pleistocene is marked by the Late Khazarian transgression—a basin with a low (−10 m) level [
3,
4,
5,
6]. The Caspian endemic
Didacna surachanica is the prevailing species in its malacofauna [
4,
5,
6,
23,
24,
25]. Popov [
26,
27], using data on the boreholes drilled in the northwestern Caspian Sea, has established that at the beginning of the Late Pleistocene in the Caspian Sea, there developed an independent large transgression—Hyrcanian (Girkanian). Specific features of its fauna are the prevalence of
Didacna cristata,
D. subcatillus, and
D. hyrcana, and the presence of warm- and fresh-water
Corbicula fluminalis. At the same time, Popov [
27] suggested that the Late Khazarian transgression associated with
Didacna surachanica as the prevailing species took place in the Middle Pleistocene. Most researchers deny the existence of the Hyrcanian basin in the Caspian history [
4,
5,
28,
29,
30]. However, analysis of the marine borehole cores allowed us to conclude that the Hyrcanian basin existed. Two transgressive basins, the Late Khazarian and the Hyrcanian, evolved in MIS 5 [
22,
23]. The discussion has been reopened [
31,
32,
33,
34,
35] and the question of the Hyrcanian stage in the history of the Caspian Region and its relationship with the Late Khazarian remains urgent.
Equally important is the understanding of the paleogeographic conditions during the existence and evolution of the Caspian basins, and their feedback to climate change. Palynological analysis of the sediments corresponding to the Khazarian basins may yield valuable information. Only scarce data on the coastal areas are available and there is practically no data on the sediments of marine boreholes. Spore-pollen analysis of unstratified Khazarian sequences began in the early 1950s along the sections of the Northern Caspian Sea Region and continued through the 1960s [
36,
37,
38,
39,
40]. Palynological data on the sediments located on the right bank of the Ural River in the Mergenevo section, defined as the Late Khazarian but lacking the biostratigraphic justification, were obtained by Nemkova [
41]. There, pebbles and clayey silt, 2 m thick (layers 9–10), lying in the lower part of the section under the Late Khalynian sediments (layers 9–10) and covered with a 15 cm interlayer of reddish-brown clay (layer 8) were defined as the Late Khazarian deposits. For layers 9–10, a series of steppe spectra was obtained (AP [i.e., trees and shrubs]—13–18%, NAP [i.e., grasses and subshrubs]—82–95%, and SP [i.e., spores]—1–5%), where pollen of
Chenopodiaceae (35–40%),
Artemisia (25–40%), and of various herbaceous species prevailed. There was also pollen of
Ephedra distachya and of aquatic and coastal-water plants. Tree species are represented by pollen of fir (3–8%), pine (8–12%), and, to a smaller extent, of birch, alder, hornbeam, linden, elm, hazel, and willow. In layer 8, the amount of tree-species pollen, primarily fir, increased, while the content of
Chenopodiaceae and
Artemisia decreased. It is possible that the spore-pollen spectra of the Mergenevo section indeed characterize a fragment in the Late Khazarian evolution of the Pleistocene vegetation and climate of the Northern Caspian Sea Region. Two spore-pollen spectra from the confirmed Upper Khazarian sediments, indicating steppe natural conditions, were obtained by Rybakova [
42].
The uncertainty of many issues in the paleogeography of the Caspian Sea or the absence of data related to its certain aspects raise the matter of the response of the basin’s natural environment to the global and regional climate change and the correlation with paleogeographic events in the adjacent territories.
The objective of our work was to reconstruct transgressive-regressive events in the Northern Caspian Sea during the Khazarian epoch of its Pleistocene history, to determine the natural conditions of the development of the basins, and to assess their response to climate change.
2. Materials and Methods
Our work is based on the studies of the sedimentary formations of the upper part of the Quaternary sequence in the Northern Caspian Sea. Conducted in this area engineering-geological surveys included a large-scale two-mode frequency seismoacoustic profiling performed concurrently using boomer and sparker sources, static sounding, and borehole drilling (up to 80 m deep). Thus, we were able to thoroughly characterize the sequence structure and its stratification. The work was conducted by OOO Morinzhgeologia (Riga, Latvia) under the direction of Yu. Bezrodnykh and B. Romanyuk. The borehole was drilled in the eastern part of the extensive shallow flat-bottomed depression Shirotnaya, confined in south to Kulalinskaya and Bezymyannaya banks (
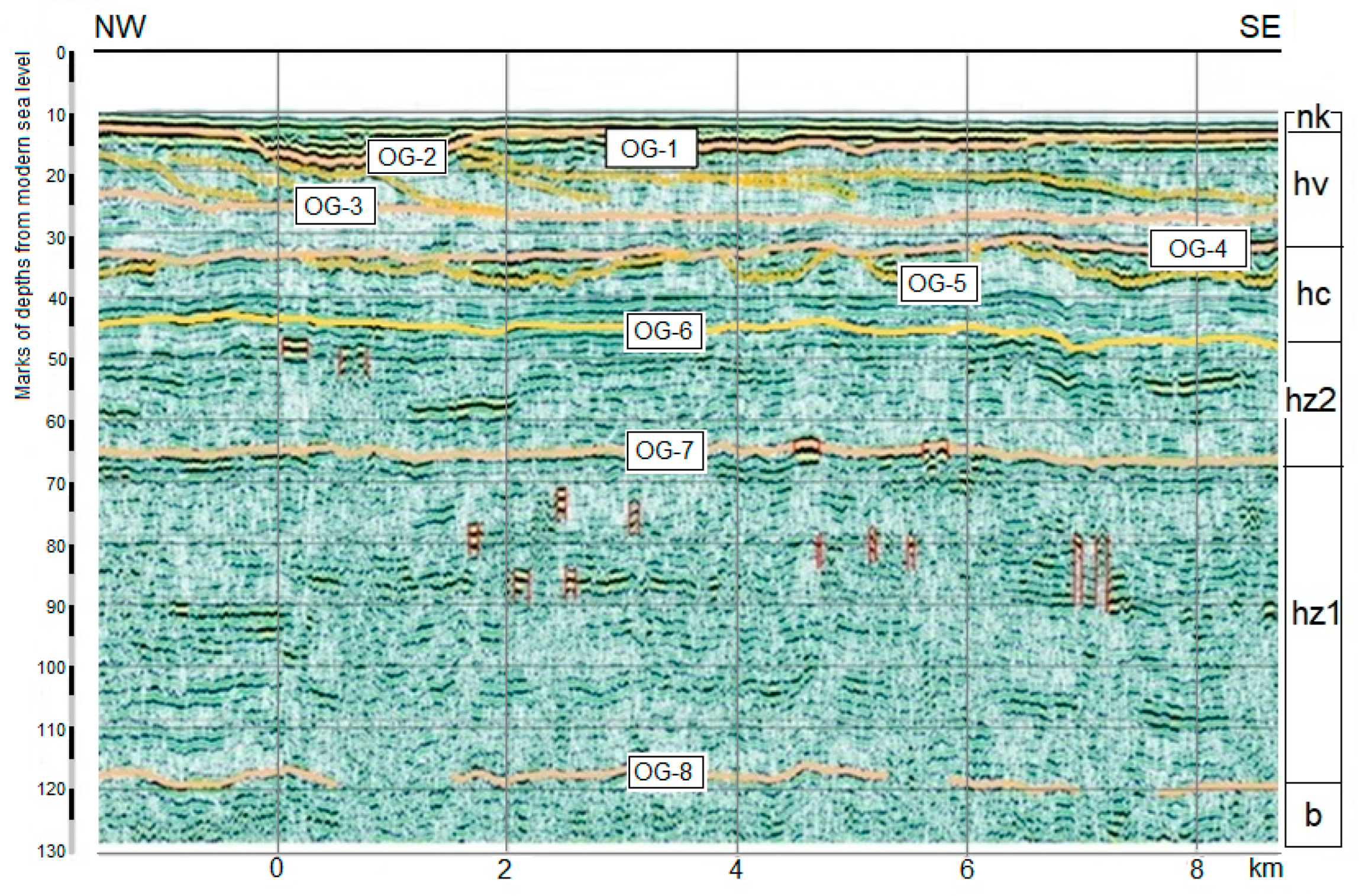
Figure 1). The Caspian Sea depth in the area is 11.9 m. The depth of drilling is 56.4 m. The core (ID NC/KOR4) is stored at the Laboratory of Recent Sediments and Paleogeography of Pleistocene, Faculty of Geography, M.V. Lomonosov Moscow State University.
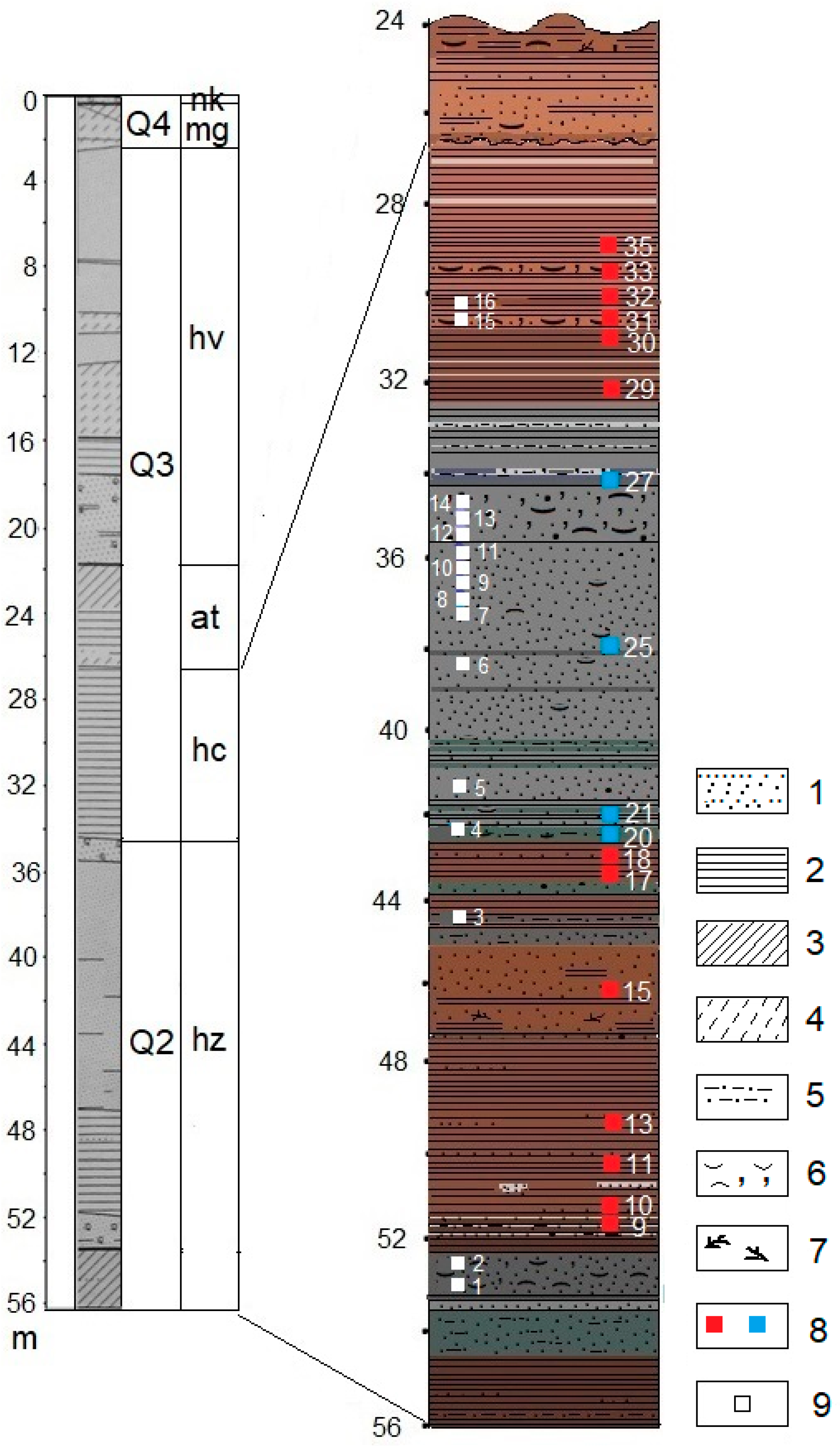
The core was subjected to lithological, malacological, and palynological analysis. This paper discusses the results related to the interval 56.0–26.6 m. The results of the core integrated analysis in the interval 26.6–21.8 m, reflecting the Atelian regressive stage in the Caspian history (MIS 4—first half of MIS 3), were published earlier [
43,
44].
Analysis of lithological structure of the sediments and the core description were conducted by V. Sorokin at the Department of Oil and Gas Sedimentology and Marine Geology, Faculty of Geology, M.V. Lomonosov Moscow State University. The lithological studies included a macro description of the core with visual determination of the structural, textural, and morphological characteristics using data on granulometric composition and density.
Malacological analysis was conducted by T. Yanina at the Laboratory of Recent Sediments and Paleogeography of Pleistocene, Faculty of Geography, M.V. Lomonosov Moscow State University. The analysis included molluscs’ taxonomic identification and preservation. For each sample, the sediment was washed through a 1 mm-sieve. Molluscs were identified to the species level and the total number of members of each species was counted. The specimens of bivalves were taxonomically identified according to paleontological descriptions with plates [
25,
27] and reference collections from the Paleontological Institute of Russian Academy of Science. The gastropod specimens were taxonomically identified according to paleontological plates and figures [
45,
46]. Fossils were studied for signs of dissolution, abrasion, and fragmentation. The findings of articulated bivalves pointed to in situ samples. We analyzed more than 3000 mollusc remains. We followed the gastropod taxonomy of [
45,
46,
47]; for bivalves, we used [
25,
47]. The data provided the basis for the subdivision of the sedimentary series and paleobasin reconstruction. Paleoreconstruction is based on the interpretation of the alternating freshwater, brackish water, and slightly brackish water mollusc assemblages. We focused on the bivalve genus
Didacna Eichwald, an index fossil for the Caspian Sea [
6,
25], for biostratigraphic subdivision. To reconstruct the Northern Caspian Sea environment, we used the published information on species’ preferred environmental conditions (salinity ranges, habitat and depth ranges) and personal observations for modern Caspian molluscs.
Palynological analysis was conducted by N. Bolikhovskaya at the Laboratory of Recent Sediments and Paleogeography of Pleistocene, Faculty of Geography, M.V. Lomonosov Moscow State University. Since the palynological studies of the extracted sediments were conducted to such extent for the first time ever, we consider it necessary to describe the method in more detail. Based on the sedimentation-lithological features of the sequence, 64 samples were selected for palynological analysis to perform climatostratigraphic subdivision of deposits, and to reconstruct the climatic and phytocenotic features of the landscapes of the Northern Caspian Sea Region. The separation of pollen and spores from sediments, their identification, and the climatostratigraphic and palaeophytocoenotic interpretation of the obtained spore-pollen data were carried out according to standard methods [
8].
Extraction of pollen, spores, and other palynomorphs was performed from 30–50 g clay samples and 70 g sand samples of the studied sediments through the separation technique [
48] and its modified version [
8] developed to derive representative pollen and spore concentrates from clay, loamy and sandy deposits, relatively poor in plant microfossils. The main procedures of this method are the decarbonization and dispergation of the samples with the use of 10% HCl and 10% KOH and also double and triple separation of their suspension in a heavy liquid with a specific gravity of 2.1–2.2 over 10 min. Contamination of the distinguished spore-pollen concentrate by large amount of pelitic particles stimulates their treatment by concentrated hydrofluoric acid (HF).
Rigorous palynotaphonomic and palynomorphological analyses of pollen, spores, and other palynomorphs was performed during the spore-pollen studies of Khazarian sediments in order to identify the autochthonous and reworked pollen and spore grains. Taphonomic studies and identifications of microfossils were carried out using an AXIO Imager D1 microscope. The identified palynomorphs were photographed with an AXIO Cam digital camera.
In some studied concentrated spore-pollen samples, as well as in Atelian sediments [
43,
44], Mesozoic miospores and broken and mineralized brown palynomorphs of the Pleistocene were present in noticeable quantities. Data on the number of redeposited forms in the studied palynological macerates are shown in the
Section 3.3. The percentage of allochthonous components from the total amount of microfossils studied in each palynological concentrate ranges from 0.5 to 15%. The composition of the repositioned forms is dominated by pre-Cenozoic miospores.
A collection of digital images of pollen, spores, and other palynomorphs of an undoubted autochthonous assemblage and redeposited microfossils was compiled for each sample studied. Examples of pollen and spores in situ and reworked grains are shown in
Figure 2.
Thus, thorough palynotaphonomic and palynomorphological studies were carried out to differentiate between autochthonous and allochthonous microfossils. As a result, 17 representative spore-pollen spectra have been obtained to date, which allowed for climatostratigraphic subdivision of the studied sequences of the Khazarian sediments and compilation of a provisional version of the history of climatic and phytocenotic events during its accumulation.
The deposits of the interglacial stages are characterized by paleofloras and spore-pollen spectra, and also reconstructed paleovegetation communities formed under climatic conditions similar to, or warmer and more humid than the current climatic conditions of the Northern Caspian Region.
Climatic and phytocenotic reconstructions derived from the results of palynological research of Quaternary sediments and definitions of the rank (interglacial or interstadial) of the paleoclimatic periods are largely based on the examination of subrecent spectra of sediments of different genesis, selected at the sampling sites both in the studied region and other areas, and on the determination of the degree of correspondence between the obtained subrecent palynospectra and pollen- and spore-producing modern plant communities.
The modern Lower Volga Region and the entire Caspian Lowland are the areas with the most arid and continental climate not only on the Russian Plain, but also on the European subcontinent as a whole. The average annual precipitation runs from 160 to 350 mm. The average annual air temperature varies from +7.6 to +9 °C. The average January temperature may reach −10 °C with absolute minima falling to −40 °C. Summer is hot; the average July temperature in the desert zone ranges from +24 to +27 °C. In the territory immediately adjacent to the Northern Caspian, zonal desert phytocenoses are common. The vegetation of the zonal desert landscapes is dominated by communities of sagebrush (Artemisia subgenus [hereinafter, subgen.], Seriphidium, Dracunculus). The vegetation cover of the coastal strip of the Caspian Sea is dominated by herbs and shrubs from the Chenopodiaceae family and Artemisia genus, which form saltwort and saltwort-sagebrush communities (composed of Halocnemum strobilaceum, Artemisia halophila, Suaeda altissima, S. confusa).
Dense thickets of common reed grass (Phragmites communis) stretch along the sea. To the north, the desert transitions into the semi-desert zone where sagebrush-gramineous cenoses dominate. The forest vegetation of the studied area is most clearly identified in the valley of the lower reaches of the Volga River, which stretches from north to south for more than 350 km; the number of taxa in the composition of woody-shrubby and herbaceous vegetation decreases in the same direction.
Thick floodplain forests of willow and meadow gramineous-forb, gramineous, gramineous-sedge, sagebrush-forb, and other communities occupy the Volga–Akhtuba floodplain. In forests, along with various species of willow (Salix spp.), there are poplar (Populus nigra, P. alba), maple (Acer tataricum), and aspen (Populus tremula); less frequent are oak (Quercus robur), ash (Fraxinus excelsior), elm (Ulmus laevis, U. carpinifolia), birch (Betula pendula, B. pubescens), and black alder (Alnus glutinosa). The dendroflora of the Volga delta is extremely poor—forest there is formed mainly by white willow (Salix alba)—and only in the northern part of the delta do ash and elm grow in willow forests. On the deluvial plumes of Baer knolls, oleaster (Elaeagnus angustifolia) is widespread.
Comparison of the obtained fossil palynospectra with the spore-pollen spectra of subrecent samples of modern subaqueous and subaerial deposits, which we collected in the phytocenoses of the Northern Caspian desert zone, allows for palynoindication of the Quaternary phytocenotic successions and identification of climatic characteristics of the reconstructed paleogeographic stages. The results of spore-pollen analysis clearly demonstrate that the subrecent palynospectra of these samples appropriately reflect the zonal and intrazonal identity and composition of pollen- and spore-producing plant communities (a,b in
Table 1).
The desert spectra are associated with a low tree-shrub pollen and spores content, (AP—0–5%) and (SP—0–4%), respectively. In the dominant group of grasses and subshrubs (NAP—95–100%), pollen of goosefoot (Chenopodiaceae) (25–97%) and of sagebrush (up to 36%, mainly
Artemisia subgen.,
Seriphidium,
A. subgen.
Dracunculus) prevail. An exception is the sample of estuary sediments, where the AP content reaches 48% due to the
Salix alba pollen. All spore-pollen spectra of the Khazarian sediments in the studied core of the IGS borehole have significantly higher contents of tree and shrub pollen (11–65%) and spores (8–28%). Therefore, for the identification of zonal paleophytocenoses we used an extensive amount of our own and published data on subrecent (subfossil) spectra of modern subaqueous and subaerial deposits from the steppe, forest-steppe, and forest zones of the southern part of the Russian Plain and the Caucasus [
49,
50,
51,
52,
53,
54].
Our paleogeographic reconstructions also used data on the spore-pollen composition in the surface layer of sediments of the entire Caspian Sea [
55,
56], including the Northern Caspian, and of the “pollen rain”, collected over the Northern Caspian [
57]. These palynological data point to a clear comparability of both the subrecent and recent spectra to the coastal zone vegetation. Thus, in the recent palynospectra of the Northern Caspian, from 92 to 100% falls on the pollen of herbaceous-subshrubby plants where pollen of goosefoot Chenopodiaceae and sagebrush
Artemisia (primarily subgen.
Seriphidium) composes 47–81% and 5–26%, respectively. In the air samples collected over the Caspian Sea, tree pollen consists mainly of pine (
Pinus sylvestris) and isolated pollen grains of birch, willow, and oak. In the palynospectra of the bottom sediments of the Northern Caspian, NAP pollen clearly prevails (90–97%). The AP group is dominated by
Pinus sylvestris. Pollen grains of birch and alder are present in smaller quantities. There are isolated occurrences of pollen of willow, hazel, oak, hornbeam, linden, and elm.
In our climatic and phytocenotic interpretation of the Khazarian spore-pollen spectra we considered all methodological aspects of the palynological analysis of the deposits of the Northern Caspian Sea, Northern Caspian Region, and other areas.
4. Discussion
4.1. Structure and Faunal Composition of the Sedimentary Sequence
The section structure and the facies-lithological composition of its sediments reflect the evolution of the Northern Caspian basin over the period of its formation. A pronounced layer of regressive sediments at the base of the core (56.00–53.60 m), indicates a significant decrease in the Caspian level and its retreat from the area of the borehole location. The transgressive eroded sedimentary sequence (53.60–47.40 m) lying above it, composed of coarse-grained coastal sands with shell material at the base and overlain by a clayey layer with silty-sand interlayers, indicates the developing transgression of the Caspian Sea. The sandy sediments contain numerous detritus and fragments of shells, often small. The faunistic group is dominated by the shells of
Dreissena rostriformis distincta. The presence of
Didacna catillus volgensis, which is often found in the Lower Khazarian deposits of the Lower Volga Region [
6], and the fragments of small and thin-valved
Didacna that are not identifiable to the species level (which is typical of the Early Khazarian transgression), rather testifies in favor of the Early Khazarian basin. The clayey, deeper part of the core thickness lacks shell material.
The water level decrease and the development of a very desalinated basin at low depths is reflected in the fine-grained sandy sediments, with clay interlayers, overlying clays (up to 42.70 m). In the range of 44.18–44.22 m, it contains shells of freshwater molluscs of Viviparus, Valvata, and Dreissena polymorpha genera. Plant remnants were noted at the base of this sedimentary layer.
A structurally similar sandy layer, which begins at 42.7 m and exhibits gradual disappearance of clay interlayers, indicates the development of the basin at low water level but under different sedimentation conditions. In contrast to the predominant brown color of the lower part of the sedimentary sequence, the sands are gray. They contain shell detritus and occasional whole shells. The faunal community is dominated by Dreissena caspia; the following also occur Dreissena polymorpha, Dr. rostriformis distincta, Monodacna caspia, Theodoxus pallasi, and Micromelania caspia. All shells are small and thin-valved; a poor living environment was apparently associated with low salinity and lack of nutrients. There are very small shells of juvenile Didacna, closest to Didacna cf. subcatillus.
In the upper part of the sandy serie, along with the species listed above, appear shell fragments of
Didacna nalivkini, a characteristic species of the Late Khazarian transgressive basin [
6,
25]. The sandy sequence ends with the layer of dark gray medium-grained sand (35.70–34.65 m), which differs in the large size of sand grains, the abundance of detritus, and shell fragments; the shell material contains a relatively large quantity of fragments of large
Didacna shells, which we identified as
D. nalivkini (also possibly
D. surachanica). This, as well as the presence of rare
Didacna subcrassa, points to the Late Khazarian age of this stratum.
Above, there are yet again deeply eroded clays of different shades of gray (34.65–32.50 m) with thin layers of chemogenic carbonate of silty grain size, with decomposed remnants of mollusc shells at the base of the clays. The shell material consists mainly of fragments, often not identifiable to the species. The whole specimens include
Theodoxus pallasi and
Micromelania caspia; the fragments include
Dreissena rostriformis distincta,
Didacna cf.
subcatillus. In the interval of 32.50–26.60 m, the gray clayey thickness transitions into dark brown clay, also with thin interlayers of chemogenic carbonate, with detritus, and occasionally whole shells of
Dreissena rostriformis distincta (prevail),
Dr. caspia,
Monodacna caspia,
Micromelania caspia,
Theodoxus pallasi,
Didacna subcatillus,
D. umbonata, and
D. cristata. The shells occur in thin silty interlayers, which indicates that the molluscs depended on the oxygen content in the basin. The accumulation of moderately deep-water clays of this layer is associated with the environment of a transgressive basin whose level exceeded the Late Khazarian basin. Its faunistic characteristics are determined by the concurrent presence of
Didacna subcatillus,
D. cristata, and rare forms of the Late Khazarian fauna, which corresponds to the faunistic characteristics of the Hyrcanian transgression suggested by Popov [
26,
27]. Weakly brackish Caspian species of
Monodacna and
Dreissena genera are abundant, which points to desalinated conditions. We regard this basin as Hyrcanian.
Above, there is a deeply eroded sandy-clayey sequence with plant remnants and interlayers of black peat-like material, corresponding to the Atelian regression of the Caspian Sea. We have described this earlier [
43,
44]. This period is associated with the formation of terrestrial landscapes on the territory of the present Northern Caspian basin.
Thus, the structure and faunistic composition of the sedimentary sequence of the studied core at 56.00–26.60 m point to the existence of three transgressive stages, namely, Early Khazarian, Late Khazarian, and Hyrcanian, separated by regressive phases. The deepest of the transgressive basins is the Early Khazarian, while the shallowest is the Late Khazarian.
4.2. Palynological Record: Climatic and Phytocenotic Changes
The palynological record of the studied sedimentary strata indicates that the Khazarian deposits were formed during three paleoclimatic stages: two warming periods, each was marked with changes in vegetation in the landscapes of adjacent areas under interglacial climate conditions, and the period of significant cooling separating these warm periods, which led to the development of periglacial vegetation in the Northern Caspian Region.
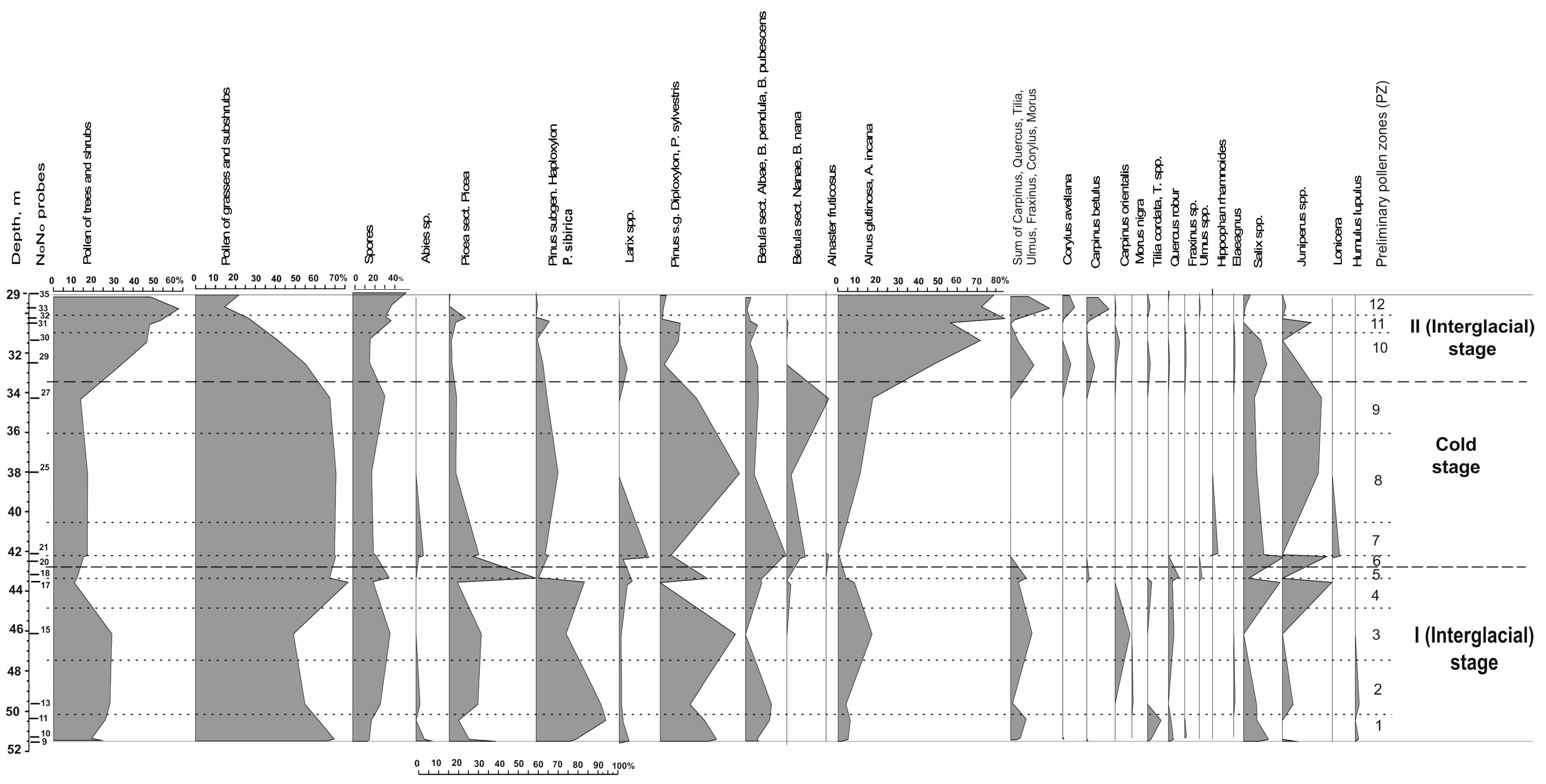
Comparison of fossil spore-pollen spectra obtained for the entire period of accumulation of the Khazarian deposits with subrecent palynospectra of the Northern Caspian and other regions suggests the existence of markedly more humid, than modern, climate of two warm paleoclimatic stages in the sediment accumulation and a significantly colder climatic stage between the two warm ones. The climate of the cold period was much more severe than the modern climate within forest-to-desert extent on the Russian Plain. Climatic and phytocenotic changes and changes in natural environments are clearly illustrated by a diagram with the graphs of the AP/NAP/SP shares, calculated relative to their sum in each spectrum, as well as the share of pollen of all taxa in the group of trees and shrubs, calculated relative to the sum of AP of each spectrum (
Figure 5).
The diagram shows the provisional palynozones identified according to the data obtained at the present stage of the research. A spore-pollen diagram with the composition and content of palynomorphs in all groups will be compiled and published after obtaining the maximum possible number of representative spectra for the entire 56 m sequence of deposits penetrated by the borehole. A complete palynological record of such a representative thickness of the marine sediments of the Caspian will make it possible to rectify the boundaries of all Late Pleistocene paleoclimatic stages.
4.2.1. The First Warm Period
The first warm (interglacial rank) period of accumulation of deposits of the Khazarian transgression in the Northern Caspian is marked with seven representative spore-pollen spectra of samples Nos. 9–18 in the thickness at 43.25–51.45 m composed of dark brown clays with interlayers and lenses of fine sand or silt where grass and the shrub pollen (NAP) prevails (51–79%), followed by the tree and shrub pollen (AP) (11–30%) and the spores of higher spore plants (SP) (8–19%) (
Table 3,
Figure 5). The palynological record obtained so far for the sediments of this thickness (PZ 1–PZ 5) reflects the dominance of open steppe landscapes.
According to the composition and percentage of pollen in the prevailing group (NAP), in the territory adjacent to the Caspian paleobasin in the grass-subshrub cover of the steppes and limitedly developed forests, depending on the ecologo-cenotic conditions, sagebrush-goosefoot, gramineous-forb, forb-gramineous and, less often, goosefoot-sagebrush associations dominated. The spore-pollen spectra are dominated by pollen Chenopodiaceae/Amaranthaceae (31–48%) and Artemisia (9–40%; primarily Artemisia subgen. Seriphidium, however, two samples at 50.4 and 51.4 m had a greater share of Artemisia subgen. Euartemisia). The share of ephedra, gramineous, and sagebrush pollen is 1–2%, 7–20%, and 3–10%, respectively. At a relatively low content of forb pollen (4–15%; only in the sample at 46.1 m—21%), the composition of its taxa (including Fabaceae) is relatively diverse—Asteraceae, Arctium sp., Cichoriaceae, Polygonaceae, Rosaceae (Agrimonia), Lamiaceae, Apiaceae, Ranunculaceae, Brassicaceae, Valerianaceae, Liliaceae, Alliaceae, Iridaceae, Plantaginaceae, Euphorbiaceae, and others. Pollen of aquatic and coastal aquatic plants (3–16% of the total NAP pollen) is present in all spectra—burweed (Sparganium), pondweed (Potamogeton), foxtail (Myriophyllum), and cattail (Typha). In six spectra, Sparganium pollen prevails; in the spectrum of the sample at 51.45 m, the share of water plantain (Alismataceae) is greater.
Among higher spore plants, the greater share belongs to the spores of ferns (Polypodiaceae—from 6 to 47%) and sphagnum and green mosses (14–48% and 2–57%, respectively), as well as to water fern Salvinia natans (0–37%).
The group of trees and shrubs is dominated by coniferous pollen, most of which belongs to the pollen of dark coniferous trees—spruce (Picea sect. Picea) and stone pine (Pinus subgen. Haploxylon, Pinus sibirica) (the total share is 29–49%), with a very noticeable role of Scots pine (Pinus sylvestris, P. subgen. Diploxylon—0–39%). The share of birch (Betula sect. Albae, Betula pendula, B. pubescens—up to 13%), alder (Alnus glutinosa, Alnus incana—up to 17%), and willow (Salix—up to 19%) is significant. The interglacial natural environment is manifested by the constant presence of pollen from broad-leaved species—hornbeam, oak, linden, ash, elm, and others (Carpinus betulus, C. orientalis, Quercus robur, Tilia cordata, Fraxinus sp., Ulmus sp., Corylus avellana, Morus nigra), whose share in the spectra of the climatic-optimum phases is quite high—4–11%.
The predominance of dark coniferous species pollen (
Picea sect.
Picea,
Pinus subgen.
Haploxylon) in the steppe palynospectra in the AP group with a fairly high content of pollen from broad-leaved trees, birch, alder, and willow is a phytocenotic nonsense, which may be explained by high pollen productivity and transportability of coniferous taxa pollen. The pollen of spruce and stone pine probably entered the Khazarian marine sediments by air and water from the coniferous forests of the Volga Upland and the Obshshy Syrt, which were relics of the boreal taiga forests of one of the preceding glacial epochs of the Middle Pleistocene. It was worth noting that according to the palynological record of the loess-soil sequence of the Otkaznoye sequence located in the Northwestern Caspian Region, the Middle Neopleistocene Kaluga cryochron was the only ice epoch in the Neopleistocene when the belt of dark coniferous forests of the Caucasus descended down to the Middle Kuma valley during one of the phases of its cryohygrotic stage [
10]. This phase was marked by a cold and relatively humid climate; spruce forests dominated in the highest parts of the watershed plateau, while cedar-spruce forests with larch and birch occupied the lower watershed levels and slopes.
On the spore and pollen diagram of the Lower Khazarian deposits (
Figure 5), the curve of the total pollen of broad-leaved species shows two maxima (11%) at 46.1 m and 51.35 m. The maxima are separated by a fall to 1% at 49.6 m, accompanied by a rise in the amount of juniper pollen (
Juniperus) and spores, among which sphagnum moss microfossils dominated (48%). Freshwater algae
Botryococcus and
Pediastrum were also abundant. This minimum of pollen from broad-leaved species most likely reflects the intraglacial (endothermal) cooling since the lithological composition of the sediments did not change.
In the warm phase preceding the endothermal, as evidenced by the spectra of samples Nos. 9 through 11 (PZ 1), the dominant vegetation consisted of ash-oak-linden communities with an admixture of birch and possibly pine, occupying the highest hypsometric levels in the Lower Volga paleovalley. Willow forests, with broad-leaved trees and alder, dominated in the floodplain forests’ composition.
During the endothermal period (PZ 2), goosefoot-sagebrush, with ephedra and gramineous-sedge-forb cenoses dominated, while willow forests with juniper in the undergrowth and shrub layer prevailed in the floodplain forest habitats. The existence of waterlogged ecotopes is evidenced by the high content of Sphagnum spores.
A relatively large amount of pollen of oriental hornbeam (Carpinus orientalis—8%) in the sample at 46.1 m (PZ 3), the presence of Morus nigra pollen, changes in the content of spruce, cedar pine, Scots pine, oak, linden, and alder pollen in the spectra, a markedly higher share of pollen from alder, willow and juniper (PZ 3–4), and the appearance in the spectrum of the final phase (PZ 5) of common hornbeam and elm (Carpinus betulus, Ulmus) reflect significant transformations that took place in the forest communities in the second half of this warm stage. Our future plans include supplementing the palynological record of the entire considered warm stage with new data that will allow us to compile a detailed chronicle of climatic and phytocoenotic successions. Therefore, this paper does not cover detailed reconstructions of the phases in the development of vegetation recorded by PZ 1 through PZ 5 palynozones.
In general, it is important to note that the composition of characteristic taxa of the dendropalynoflora of the first warm stage (I Interglacial Stage) of the Khazarian transgression is rather rich—
Picea sect.
Picea,
Pinus subgen.
Haploxylon,
Pinus sylvestris,
P. subgen.
Diploxylon,
Juniperus,
Carpinus orientalis,
C. betulus,
Quercus robur,
Tilia cordata,
Fraxinus sp.,
Ulmus sp.,
Corylus avellana,
Morus nigra,
Alnus glutinosa,
Alnus incana,
Betula pendula,
B. pubescens,
Salix, and others. We should emphasize the significant role of oriental hornbeam (
Carpinus orientalis) pollen. Detailed palynological studies of the Pleistocene sediments of the Northwestern Caspian Region (the above-mentioned section Otkaznoye in the Middle Kuma valley), as well as of the Upper Don (Strelitsa) and other areas of the southern part of the Russian Plain, showed that
Carpinus orientalis and oriental hornbeam groves are the most important climatic and stratigraphic components of the flora and phytocenoses of the interglacial stage comparable to MIS 7 [
8,
58,
59]. In this regard, considering all palynological evidence, it is reasonable to assign the marine Khazarian deposits at 43.25–51.45 m to the last interglacial epoch of the Middle Pleistocene (MIS 7).
4.2.2. The Cold Period
The sediments with typical periglacial patterns and floras of the cold stage in the Khazarian epoch have not been previously identified in the Khazarian marine sequences. The results of the spore-pollen analysis of the deposits of the Khazarian cold stage are shown in
Figure 5 (PZ 6-PZ 9) and
Table 4. The natural environment of this cryochron reflects four representative spectra characterizing the time of accumulation of sediment sequence at 32.65–43.25 m. Pollen of broad-leaved species is absent in all the spectra. The share of NAP pollen is the highest (69–72%), with dominant Chenopodiaceae/Amaranthaceae and sagebrush
Artemisia subgen.
Seriphidium,
A. subgen.
Euartemisia,
A. subgen.
Dracunculus (up to 73% in total). The content of spores (10–17%) is relatively high. The spores of green and sphagnum mosses dominate. The share of spores of Polypodiaceae ferns and club mosses is quite noticeable and characterized by the presence of arcto-boreal tundra and north-taiga taxa (
Lycopodium appressum,
Diphasium alpinum,
Selaginella selaginoides). Boreal flora (
Picea sect.
Picea,
Pinus sibirica,
P. sylvestris,
P. subgen.
Diploxylon,
Juniperus,
Betula sect.
Albae,
B. pendula,
B. pubescens) dominates the AP group; pollen of
Betula sect.
Nanae and
B. nana plays a significant role. Only one pollen grain of
Alnaster fruticosus/Duschekia fruticosa was found.
These palynological data indicate that in the territory adjacent to the Northern Paleo-Caspian, open landscapes of periglacial steppes of sagebrush-goosefoot communities with ephedra (Ephedra distachya) were widespread and blended into or adjoined grass-gramineous, gramineous-grass, and gramineous-sedge groups, wetlands, coniferous and coniferous-birch woodlands and shrub formations of dwarf birch in valley floodplains, gullies, and other biotopes. At the present stage of research, we can suggest the following climatic and phytocenotic successions in the evolution of the vegetation cover. In the initial phase (PZ 6), forested areas were composed of spruce-cedar and larch-birch communities with juniper and Betula nana in the shrub layer with prevalence of green mosses, and of willow stands in floodplain ecotopes. In the next phase (PZ 7), the share of larch and Pinus sibirica increased in these limited forest formations. The phase identified by the spectrum at 38 m (PZ 8), where the content of Betula sect. Nanae and B. nana pollen decreases to 2% and Scots pine (Pinus sylvestris) pollen dominates, reflects a long-term interstadial warming with prevailing pine open forests, alder forests, and juniper thickets in forested areas. High content of spores of Sphagnum, Huperzia, and Lycopodium sp. in the spectrum apparently indicates the expansion of waterlogged biotopes. The presence of Athyrium filix-femina spores, which now grow on wet peat-humic soils in moist coniferous and mixed forests, indicates rather mild interstadial climatic conditions. The last established phase (PZ 9) of the Khazarian cooling was marked by a greater participation of alder in floodplain forests and a new expansion of dwarf birches into shrub communities.
Comparison of the floristic and phytocenotic reconstructions obtained for the Khazarian time with the climatic and phytocenotic successions of the Cherepet’ interglacial and Dnieper cold stage described in detail according to the loess-soil sequence of the section Otkaznoye [
8], where a significant role of broad-leaved species in the pine-birch and alder forests of the Dnieper-Moscow interstadial stage has been established in the Middle Kuma valley, implies that the layer of Khazarian sediments at 32.65–43.25 m is an important lithological fragment of the paleoclimatic events of the middle of the Dnieper-Moscow Dnieper (=Moscow) cryochron.
Analysis of the entire palynological record of the sediment sequence at 29.0–51.45 m suggests that the core NC/KOR4 contains no deposits of an extremely long period corresponding to the second half of the Dnieper cryochron.
4.2.3. The Second Warm Stage
The second warm stage of the Khazarian transgression associated with the landscape and climatic conditions of the accumulation of the lithological varieties of the overlying brown clays found in the core at 29.0–32.65 m is marked by six representative spore-pollen assemblages (
Table 5,
Figure 5).
The presence of Carpinus betulus, C. orientalis, Quercus robur, Tilia cordata, Fraxinus sp., Ulmus sp., Corylus avellana, Alnus glutinosa, A. incana, and others in the composition of the dendropalynoflora pollen (these species are indicative of the Mikulino flora of the Middle and the South Russian Plain), the high content (with the prevalence of alder) of broad-leaved species pollen, and the predominance of Carpinus betulus in the nemoral complex, indicate that the formation of clays at 29.0–32.65 m was occurring during one of the intervals of the thermohygrotic stage of the Mikulino interglacial. The obtained palynological record (PZ 10-PZ 12) reflects three phases of the transformations that took place in the plant communities of the prevalent forest-steppe and forest landscapes, whose forest stands were dominated by alder (Alnus glutinosa, A. incana) during most of this interval.
The first phase (PZ 10) was marked by the dominant forest-steppes communities (AP—34–48%, NAP—43–57%, SP—9%), where the most favorable habitats for broad-leaved species were occupied by ash-oak-linden-hornbeam formations with an oriental hornbeam grove (Carpinus orientalis) and hazel (Corylus avellana) in the undergrowth; alder forests inhabited valley, swampy, and lowland ecotopes with high groundwater levels. The total share of broad-leaved species pollen in the spectra reaches 12%. Grass cover of forests consisted mainly of forbs (ranunculaceae, brassicaceae, caryophyllaceae, lamiaceae, apiaceae, asteraceae, and others), sedges, and ferns (including Polypodium vulgare и Athyrium filix-femina). Open steppe landscapes were occupied by sagebrush-goosefoot with ephedra, gramineous-sedge, and gramineous-forb communities.
The second phase (PZ 11) reflects the phytocenoses of forest-steppe landscapes (AP—50–55%, NAP—28–30%, SP—17–20%) of the endothermal cooling, during which broad-leaved trees almost completely disappeared from the forests. The share of juniper and, possibly, other conifers increased noticeably in the dominant alder forest stands. In the grass-shrub cover, the diversity of forbs has significantly decreased. The role of gramineous species has increased, as well as the role of Polypodiaceae and grape-fern (Botrychium) among higher spore plants.
The third phase (PZ 12) is associated with the prevalence of forest landscapes in the territory adjacent to the Northern Caspian Sea (AP—50–65%, NAP—15–22%, SP—20–28%). Dominated alder forests and hornbeam forests with an admixture of linden (Tilia cordata) and abundant undergrowth of hazel. The total pollen of broad-leaved species in the spectra approached 18%. Gramineous-forb communities and Polypodiaceae ferns prevailed in the grass-shrub layer.
The climatic conditions of the second warm stage of the Khazarian transgression were undoubtedly more humid than during the first warm stage, which was marked by the dominant steppe landscapes. Analysis of the entire palynological record of the sediment sequence suggests that the core NC/KOR4 contains no deposits of the thermoxerotic stage of the Mikulino interglacial.
4.3. The Climatic Conditions during the Khazarian Epoch
The results of climatic and phytocenotic reconstructions allow reaching a conclusion about the climatic conditions of the transgressive-regressive stages of the Northern Caspian in the Khazarian epoch.
The final stage of the early Khazarian transgression evolved in the warm interglacial epoch (MIS 7). During the first warm phase of this interglacial, including endothermal cooling, the sea level rose, as evidenced by the relatively deep-water composition of the sediments and the abundance of one of the deepest Caspian bivalve species, Dreissena rostriformis distincta. In the second warm phase that followed the endothermal, the level decreased. At the location of the borehole, there was a heavily desalinated basin with freshwater and weakly brackish water malacofauna.
With the onset of cooling, the water level began to gradually rise, and Caspian species of molluscs, mainly Dreissena caspia, emerged and propagated, indicating desalinated conditions. The water level rise was not significant, the basin evolved at low levels. The environmental conditions for the habitat of molluscs were not favorable. The long-term interstadial warming has changed the environment in the basin; under more favorable conditions for the malacofauna, the composition of molluscs became richer and characteristic representatives of the Late Khazarian fauna, Didacna nalivkini, appeared. The basin remained fairly shallow. It is unclear when the development of the Late Khazarian basin ended, because, judging by the palynological evidence, a significant sediment thickness was eroded by the subsequent events. The final transgression of the Khazarian stage, the Hyrcanian, represents a relatively deep-water basin, developed in the thermohygrotic stage of the interglacial (Mikulino, Eemian) in MIS 5.
We believe that the main achievement of the research work presented herein is the palynological reconstructions based on the core sediments of an offshore borehole. Such studies, in contrast to malacological studies, are rare. The results of our palynological research have introduced new discussion issues into the paleogeography of the Northern Caspian Sea, and indeed of the entire Caspian Region.
The ideas about the evolution of the Early Khazarian transgression in the second half of the Middle Pleistocene are relatively well-established. However, the performed palynological reconstructions demonstrated for the first time ever that its transgressive phase evolved during the first warm phase of the MIS 7 interglacial and in the endothermal cooling.
The Caspian regression took place in the second phase and the detailed reconstruction of its evolution has yet to be completed. The onset of the transition of the Late Khazarian transgression into the interstadial warming of the glacial epoch MIS 6 turned out to be unexpected and to contradict previous constructions.The imperfection of the geological record prevents us from reaching a conclusion about the full cycle of the development and culmination of the Late Khazarian transgression. According to the established opinion, the Late Khazarian transgressive epoch “commences” the Late Pleistocene in the Caspian Region and corresponds to the warm interglacial epoch. Although, as noted in the introductory section of this paper, there is also a point of view about the development of the Late Khazarian transgression at the end of the Middle Pleistocene [
27].
According to climatic and stratigraphic constructions based on the palynological data, this section contains only deposits corresponding to the thermohygrotic phase of the warm epoch; the Hyrcanian transgression also evolved at that time. Recent studies have shown that the age of the Hyrcanian deposits in the Manych depression, obtained by optically stimulated luminescence (OSL), is 107 ± 7 ka [
60]. Under cold and dry conditions of glaciation (MIS 4), the Caspian regressed [
44].
5. Conclusions
A comprehensive study of the core of the borehole drilled to a depth of 56.4 m in the eastern part of the flat-bottomed Shirotnaya basin allowed us to identify the evolutional stages of the Northern Caspian Sea reflected in its structure and to reconstruct the paleogeographic sedimentation conditions.
The sedimentary sequence at 29.0–56.4 m characterizes the Khazarian transgressive-regressive epoch in the history of the Caspian Sea. The structure, facies-lithological, and malacological composition of the core capture three transgressive stages of the Caspian, namely Early Khazarian, Late Khazarian, and Hyrcanian, separated by regressive phases. The deepest of them was the Early Khazarian, marked by the abundance of Dreissena rostriformis distincta in the composition of the malacofauna. This stage was followed by the regressive basin with desalinated conditions and the presence of freshwater and slightly brackish water malacofauna. The Late Khazarian transgression developed at low levels; its sandy sediments were marked by the presence of Didacna nalivkini and D. surachanica. The final transgression of the Khazarian epoch is the Hyrcanian. It was a relatively deep-water basin with Didacna subcatillus. Sediments of the Hyrcanian transgression overlie the Upper Khazarian with deep erosion, indicating the incompleteness of the geological record in the core interval.
The representative spore-pollen assemblages made it possible to carry out the climatic and stratigraphic subdivision of the studied Khazarian deposits and to suggest a provisional version of the history of climatic and phytocenotic events during their accumulation. The palynological record shows that the Khazarian deposits formed during three paleoclimatic stages, specifically two warmings of the interglacial rank, each characterized by changes in vegetation in the landscapes of the adjacent land areas, and a significant cooling separating them, which led to the development of periglacial vegetation in the Northern Caspian Region.
The composition of the characteristic taxa of the dendropalynoflora of the first warm stage, including the significant role of oriental hornbeam (Carpinus orientalis) pollen, allowed us to conclude that it was comparable to the last interglacial epoch of the Middle Pleistocene MIS 7. Two warm phases separated by endothermal cooling were established there.
The palynological spectra of the cold stage of the glacial rank (MIS 6) indicate that open landscapes of periglacial steppes were widespread in the territory adjacent to the Northern Paleo-Caspian, where sagebrush-goosefoot communities with ephedra (Ephedra distachya) dominated. They shared the territory of valleys, ravines, and other biotopes with forb-gramineous, gramineouse-forb, and gramineous-sedge communities, waterlogged areas, coniferous and coniferous-birch woodlands, and shrub formations of dwarf birch (Betula nana). Climatic and phytocenotic successions (four palynozones) were identified in the evolution of the vegetation cover, indicating climatic changes during the cold stage. Thus, the third palynozone reflects a long-term interstadial warming with pine open forests, alder stands, and juniper thickets that dominated forested areas. The sediments with typical periglacial patterns and floras have never been found in the Khazarian marine strata before.
The second warm stage, judging by the participation in the dendropalynoflora of representative species of the Mikulino flora of the Middle and the South Russian Plain, is comparable to the thermohygrotic stage of the Mikulino (Eemian) interglacial (MIS 5). The palynological record reflects three transformation phases that took place in the plant communities of the dominant forest-steppe and forest landscapes, where alder (Alnus glutinosa, A. incana) prevailed in the forest stands during most of this interval.
Analysis of the entire palynological record of the Khazarian sedimentary thickness leads to the conclusion that the borehole core contains no deposits of a long period corresponding to the second half of the Dnieper cryochron (MIS 6) and the thermoxerotic stage of the Mikulino interglacial (MIS 5).
The results of climatic and phytocenotic reconstructions allowed drawing a conclusion about the climatic conditions of the transgressive-regressive stages of the Northern Caspian Sea developing in the Khazarian epoch. The Early Khazarian transgressive stage took place during the warm interglacial period (MIS 7). Over the first warm phase of this interglacial, including endothermal cooling, the sea level rose; during the second warm phase, it dropped, which resulted in a significant impact of freshwater on the basin. With the onset of cooling, the level of this basin began to gradually rise, but it continued to evolve at low levels and the composition of the malacofauna was poor. The onset of the interstadial warming promoted conditions favorable for the habitation of the malacofauna whose composition became richer; thus, characteristic fauna species of the Late Khazarian appeared. The basin remained fairly shallow. The further development of the events cannot be reconstructed, because, judging by the palynological evidence, a significant thickness of the sediments was eroded. The final transgression of the Khazarian epoch, the Hyrcanian, evolved in the thermohygrotic stage of the interglacial in MIS 5.
The results of our palynological studies have introduced new discussion points into the paleogeography of the Northern Caspian Sea that will be addressed at the next stages of our research.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}