
Genome-Wide Identification and Expression Profiling of the NCED Gene Family in Cold Stress Response of Prunus mume Siebold & Zucc

Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification and Physicochemical Properties of NCEDs Gene Family
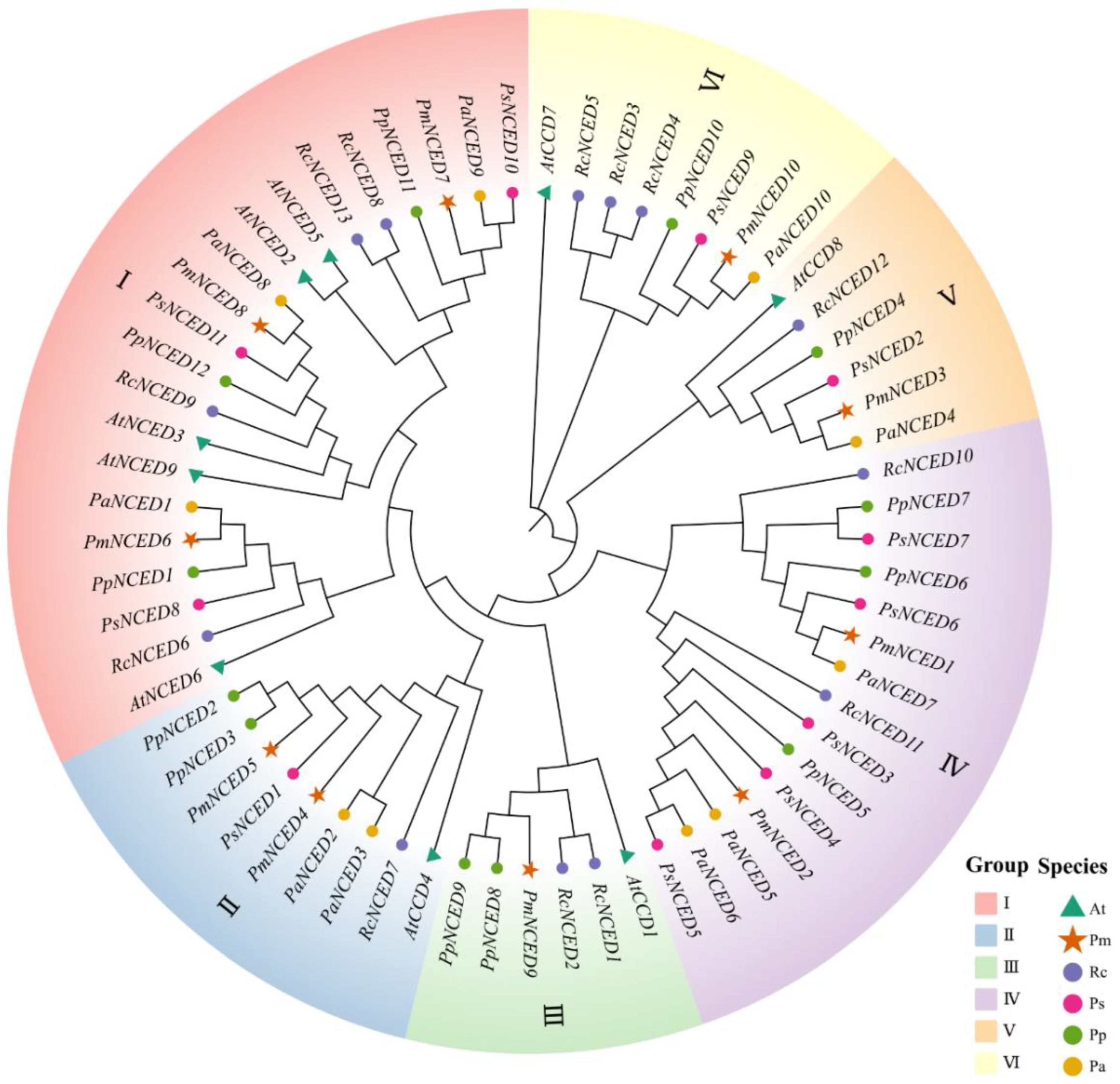
2.2. Phylogenetic Analysis
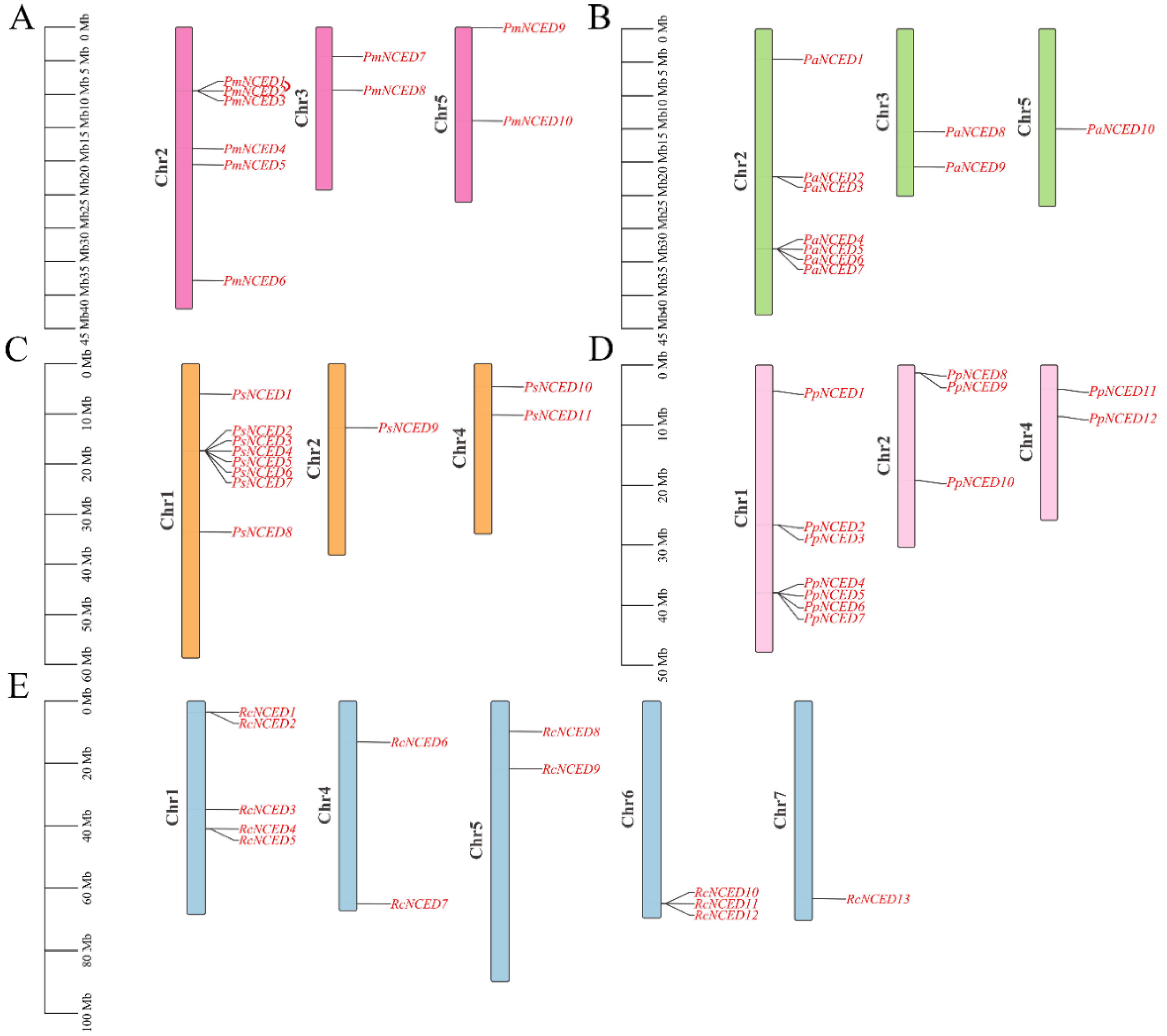
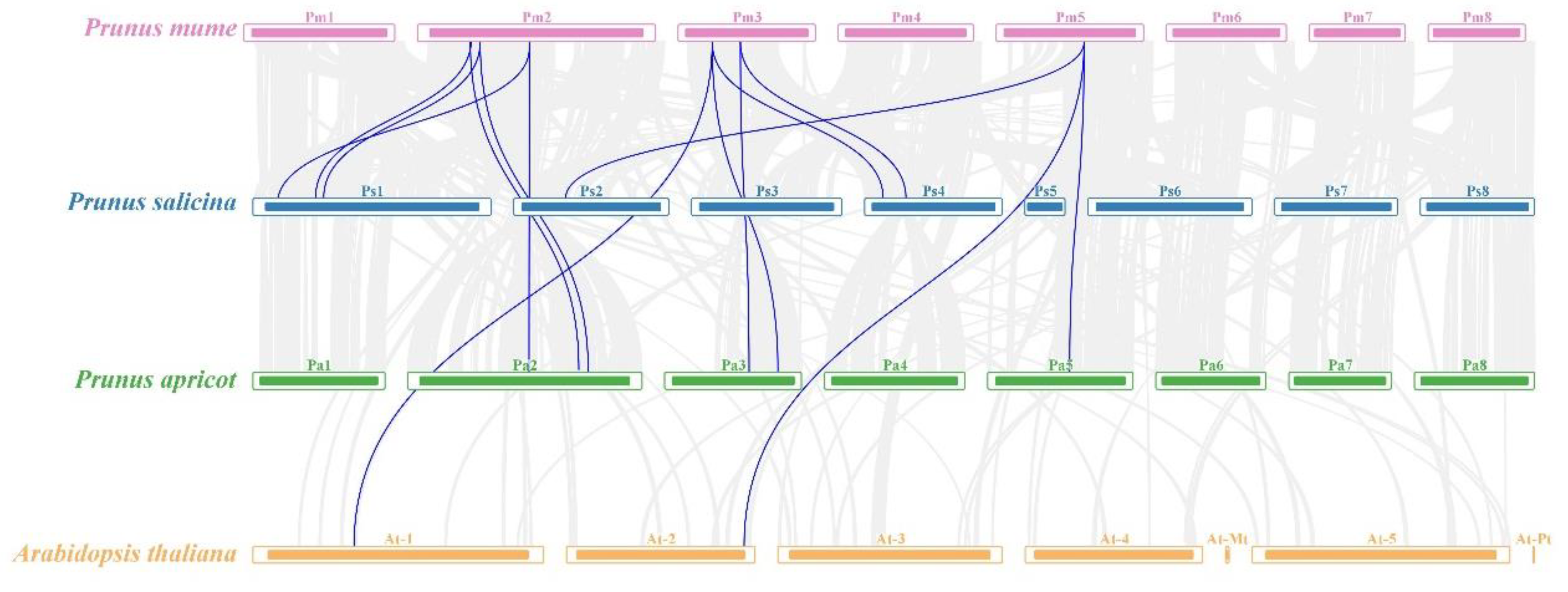
2.3. Chromosomal Localization and Collinear Analysis
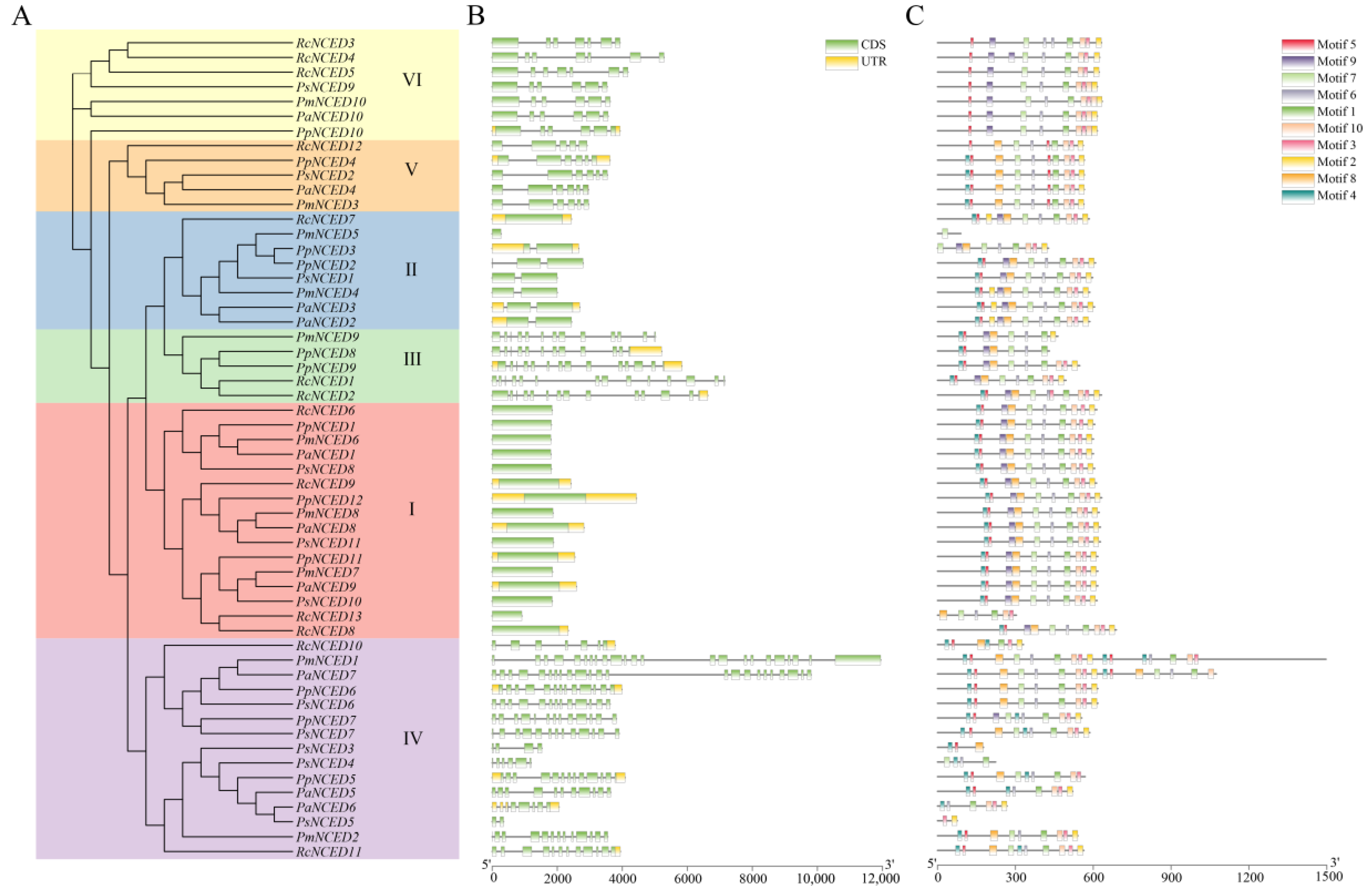
2.4. Gene Structure and Protein Conserved Motif Analysis
2.5. Cis-Acting Regulating Element Analysis
2.6. Expression Pattern Analysis
3. Results
3.1. Genome-Wide Identification of NCEDs Gene Family in Five Rosaceae Species
3.2. Phylogenetic Analysis of NCEDs Gene Family
3.3. Chromosomal Localization of NCEDs Gene Family in Five Rosaceae Species
3.4. Gene Structure and Conserved Motifs Analysis of NCED in Five Rosaceae Species
3.5. Collinear Analysis of NCEDs in Prunus Species and A. thaliana
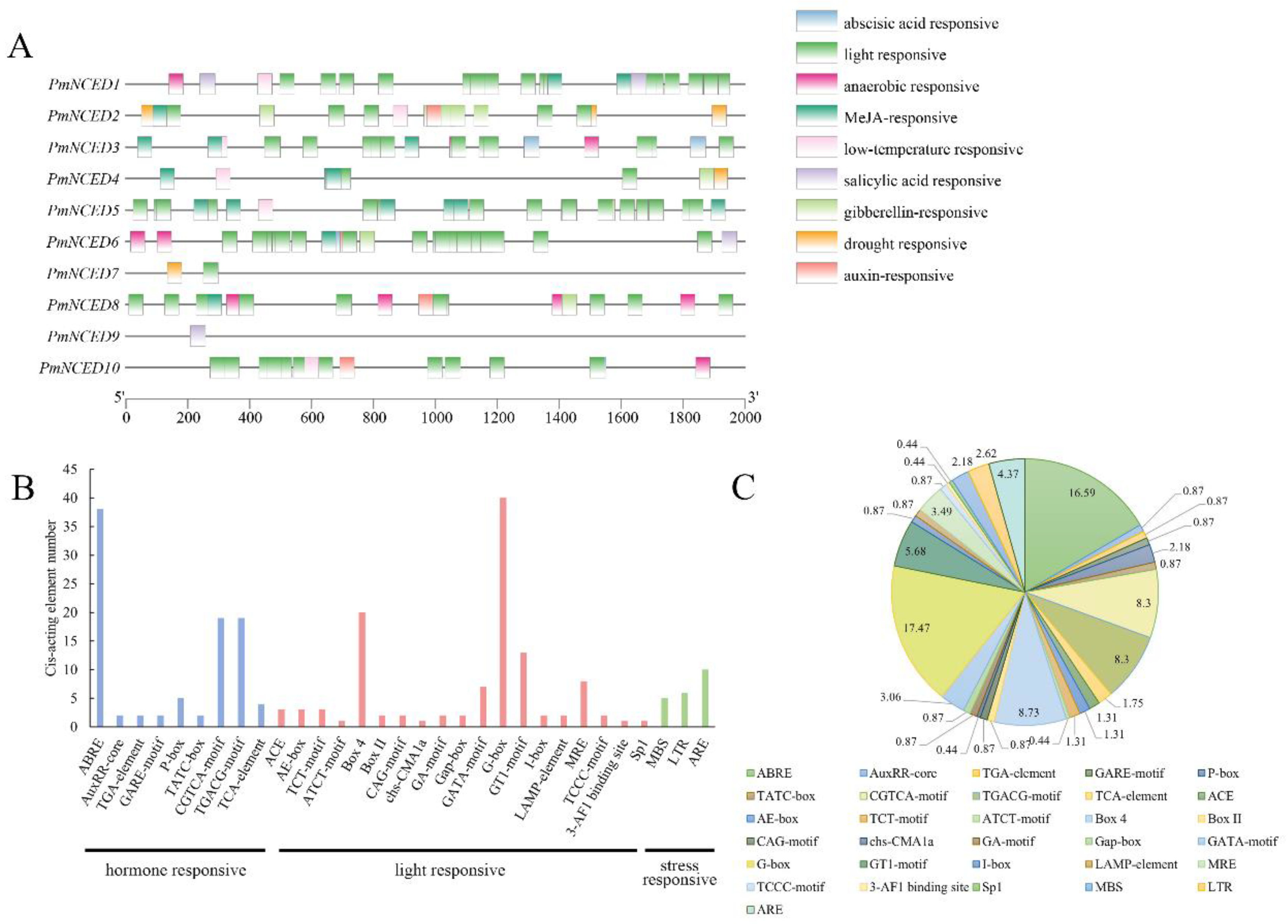
3.6. Analysis of Cis-Elements in the Promoters of PmNCEDs
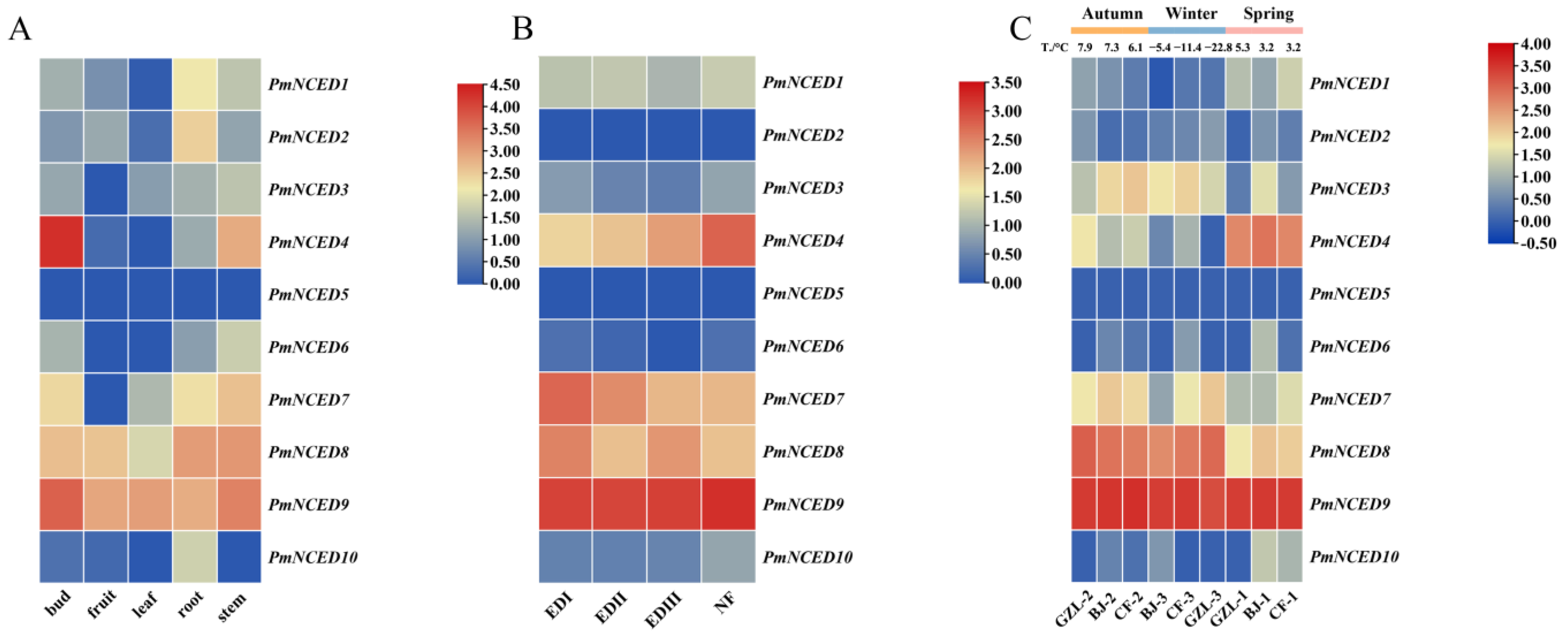
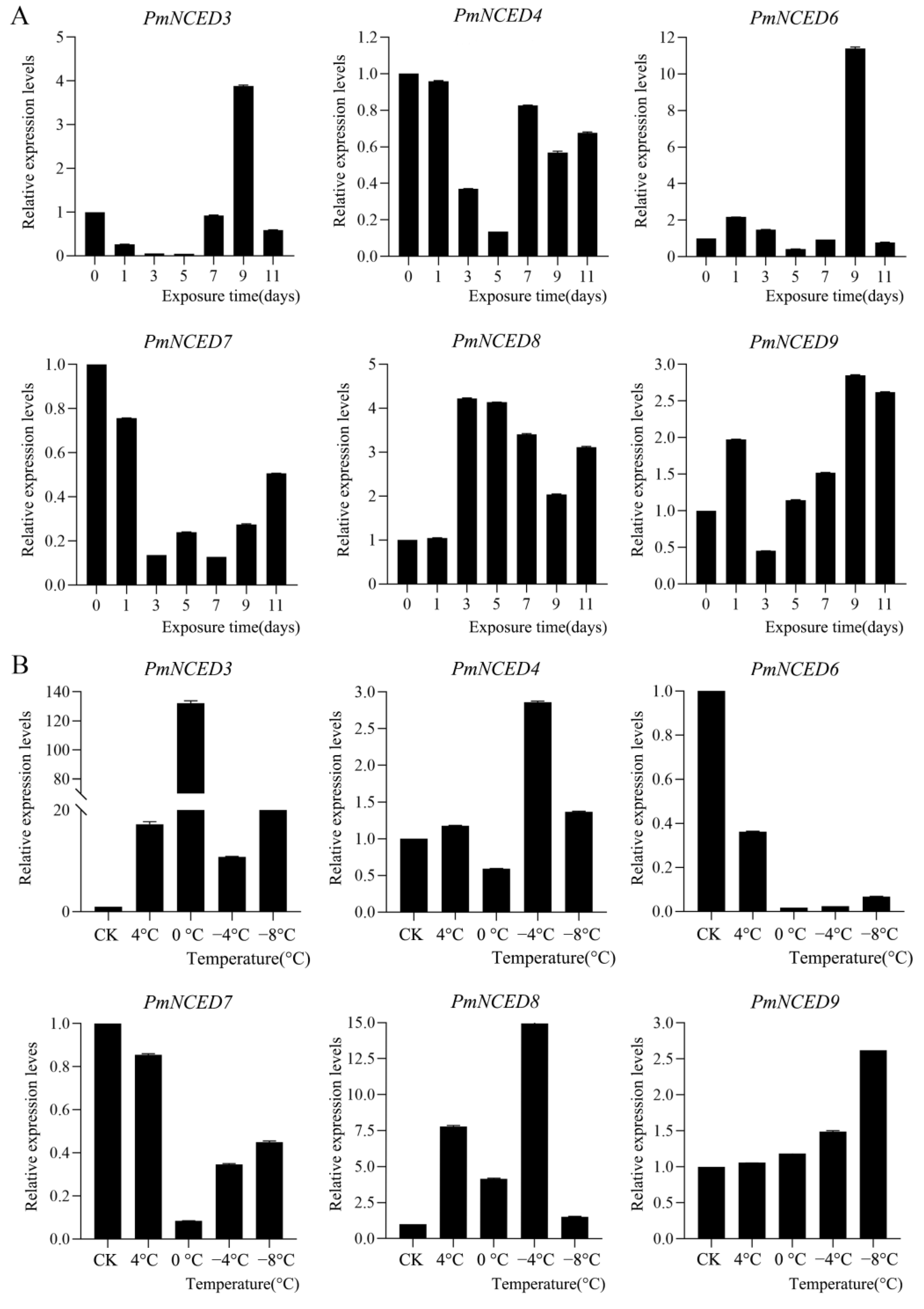
3.7. Expression Pattern of PmNCEDs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Peng, T.; Guo, C.; Yang, J.; Wan, X.; Wang, W.; Zhang, J.; Bao, M.; Zhang, J. Transcriptome analysis revealed molecular basis of cold response in Prunus mume. Mol. Breed. 2023, 43, 34. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, X.; Zheng, T.; Zhang, Z.; Zhang, Y.; Jiang, L.; Ahmad, S.; Sun, L.; Wang, J.; Cheng, T.; Zhang, Q. Genome-wide analysis of the NAC transcription factor gene family reveals differential expression patterns and cold-stress responses in the woody plant Prunus mume. Genes 2018, 9, 494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Zhang, H.; Sun, L.; Fan, G.; Ye, M.; Jiang, L.; Liu, X.; Ma, K.; Shi, C.; Bao, F.; et al. The genetic architecture of floral traits in the woody plant Prunus mume. Nat. Commun. 2018, 9, 1702. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Xiong, L. Genetic engineering and breeding of drought-resistant crops. Annu. Rev. Plant Biol. 2014, 65, 715–741. [Google Scholar] [CrossRef]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene networks involved in drought stress response and tolerance. J. Exp. Bot. 2006, 58, 221–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, B.M.; Ghassemian, M. Genome organization in Arabidopsis thaliana: A survey for genes involved in isoprenoid and chlorophyll metabolism. Plant Mol. Biol. 2003, 51, 925–948. [Google Scholar] [CrossRef]
- Kovács, Z.; Simon-Sarkadi, L.; Szucs, A.; Kocsy, G. Differential effects of cold, osmotic stress and abscisic acid on polyamine accumulation in wheat. Amino Acids 2010, 38, 623–631. [Google Scholar] [CrossRef]
- Roychoudhury, A.; Paul, S.; Basu, S. Cross-talk between abscisic acid-dependent and abscisic acid-independent pathways during abiotic stress. Plant Cell Rep. 2013, 32, 985–1006. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, A.; Robitaille, G.; Castonguay, Y.; Nadeau, P.; Boutin, R. Changes in ABA and gene expression in cold-acclimated sugar maple. Tree Physiol. 1997, 17, 31–37. [Google Scholar] [CrossRef]
- Fenollosa, E.; Gamez, A.; Munne-Bosch, S. Plasticity in the hormonal response to cold stress in the invasive plant Carpobrotus edulis. J. Plant Physiol. 2018, 231, 202–209. [Google Scholar] [CrossRef]
- Wang, H.; Blakeslee, J.J.; Jones, M.L.; Chapin, L.J.; Dami, I.E. Exogenous abscisic acid enhances physiological, metabolic, and transcriptional cold acclimation responses in greenhouse-grown grapevines. Plant Sci. 2020, 293, 110437. [Google Scholar] [CrossRef]
- Priya, R.; Siva, R. Analysis of phylogenetic and functional diverge in plant 9-cis-epoxycarotenoid dioxygenase gene family. J. Plant Res. 2015, 128, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S.H.; Qin, X.; Zeevaart, J.A. Elucidation of the indirect pathway of abscisic acid biosynthesis by mutants, genes, and enzymes. Plant Physiol. 2003, 131, 1591–1601. [Google Scholar] [CrossRef] [Green Version]
- Ren, H.; Gao, Z.; Chen, L.; Wei, K.; Liu, J.; Fan, Y.; Davies, W.J.; Jia, W.; Zhang, J. Dynamic analysis of ABA accumulation in relation to the rate of ABA catabolism in maize tissues under water deficit. J. Exp. Bot. 2006, 58, 211–219. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.C.; Schwartz, S.H.; Zeevaart, J.A.; McCarty, D.R. Genetic control of abscisic acid biosynthesis in maize. Proc. Natl. Acad. Sci. USA 1997, 94, 12235–12240. [Google Scholar] [CrossRef]
- Burbidge, A.; Grieve, T.M.; Jackson, A.; Thompson, A.; McCarty, D.R.; Taylor, I.B. Characterization of the ABA-deficient tomato mutant notabilis and its relationship with maize Vp14. Plant J. 1999, 17, 427–431. [Google Scholar] [CrossRef]
- Qin, X.; Zeevaart, J.A. The 9-cis-epoxycarotenoid cleavage reaction is the key regulatory step of abscisic acid biosynthesis in water-stressed bean. Proc. Natl. Acad. Sci. USA 1999, 96, 15354–15361. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Yang, C.; Wang, C.; Wei, Z.; Xia, D.; Wang, Y.; Liu, G.; Wang, Y. Time-course analyses of abscisic acid level and the expression of genes involved in abscisic acid biosynthesis in the leaves of Betula platyphylla. Mol. Biol. Rep. 2012, 39, 2505–2513. [Google Scholar] [CrossRef]
- Neves, D.M.; Filho, M.A.; Bellete, B.S.; Silva, M.F.; Souza, D.T.; Dos S Soares Filho, W.; Costa, M.G.; Gesteira, A.S. Comparative study of putative 9-cis-epoxycarotenoid dioxygenase and abscisic acid accumulation in the responses of sunki mandarin and rangpur lime to water deficit. Mol. Biol. Rep. 2013, 40, 5339–5349. [Google Scholar] [CrossRef] [PubMed]
- Iuchi, S.; Kobayashi, M.; Taji, T.; Naramoto, M.; Seki, M.; Kato, T.; Tabata, S.; Kakubari, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Regulation of drought tolerance by gene manipulation of 9-cis-epoxycarotenoid dioxygenase, a key enzyme in abscisic acid biosynthesis in arabidopsis. Plant J. 2001, 27, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Jiao, Y.; Xie, N.; Guo, Y.; Zhang, F.; Xiang, Z.; Wang, R.; Wang, F.; Gao, Q.; Tian, L.; et al. OsNCED5, a 9-cis-epoxycarotenoid dioxygenase gene, regulates salt and water stress tolerance and leaf senescence in rice. Plant Sci. 2019, 287, 110188. [Google Scholar] [CrossRef] [PubMed]
- Ahrazem, O.; Rubio-Moraga, A.; Trapero, A.; Gómez-Gómez, L. Developmental and stress regulation of gene expression for a 9-cis-epoxycarotenoid dioxygenase, CstNCED, ssolated from Crocus sativus stigmas. J. Exp. Bot. 2012, 63, 681–694. [Google Scholar] [CrossRef] [Green Version]
- Xia, H.; Wu, S.; Ma, F. Cloning and expression of two 9-cis-epoxycarotenoid dioxygenase genes during fruit development and under stress conditions from malus. Mol. Biol. Rep. 2014, 41, 6795–6802. [Google Scholar] [CrossRef]
- Iuchi, S.; Kobayashi, M.; Yamaguchi-Shinozaki, K.; Shinozaki, K. A stress-inducible gene for 9-cis-epoxycarotenoid dioxygenase involved in abscisic acid biosynthesis under water stress in drought-tolerant cowpea. Plant Physiol. 2000, 123, 553–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wang, Z.; Dong, J.; Wang, M.; Gao, H. Cloning of a 9-cis-epoxycarotenoid dioxygenase gene and the responses of Caragana korshinskii to a variety of abiotic stresses. Genes Genet. Syst. 2009, 84, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Thompson, A.J.; Jackson, A.C.; Symonds, R.C.; Mulholland, B.J.; Dadswell, A.R.; Blake, P.S.; Burbidge, A.; Taylor, I.B. Ectopic expression of a tomato 9-cis-epoxycarotenoid dioxygenase gene causes over-production of abscisic acid. Plant J. 2000, 23, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Zeevaart, J.A. Overexpression of a 9-cis-epoxycarotenoid dioxygenase gene in Nicotiana plumbaginifolia increases abscisic acid and phaseic acid levels and enhances drought tolerance. Plant Physiol. 2002, 128, 544–551. [Google Scholar] [CrossRef]
- Bolser, D.M.; Staines, D.M.; Perry, E.; Kersey, P.J. Ensembl plants: Integrating tools for visualizing, mining, and analyzing plant genomic data. Methods Mol. Biol. 2017, 1533, 1–31. [Google Scholar] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple sequence alignment using clustalW and clustalX. Curr. Protoc. Bioinform. 2002. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Yang, L.; Li, P.; Qiu, L.; Ahmad, S.; Wang, J.; Zheng, T. Identification and comparative analysis of the rosaceae RCI2 gene family and characterization of the cold stress response in Prunus mume. Horticulturae 2022, 8, 997. [Google Scholar] [CrossRef]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chen, W.; Sun, L.; Zhao, F.; Huang, B.; Yang, W.; Tao, Y.; Wang, J.; Yuan, Z.; Fan, G.; et al. The genome of Prunus mume. Nat. Commun. 2012, 3, 1318. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhuo, X.; Zhao, K.; Zheng, T.; Han, Y.; Yuan, C.; Zhang, Q. Transcriptome profiles reveal the crucial roles of hormone and sugar in the bud dormancy of Prunus mume. Sci. Rep. 2018, 8, 5090. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L. Physiological Changes and Gene Expression Pattern in Response to Low Temperature Stress in Prunus mume. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2020. [Google Scholar]
- Wang, T.; Lu, J.; Xu, Z.; Yang, W.; Wang, J.; Cheng, T.; Zhang, Q. Selection of suitable reference genes for miRNA expression normalization by qRT-PCR during flower development and different genotypes of Prunus mume. Sci. Hortic. 2014, 169, 130–137. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data ysing real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Cai, H.; Tian, S.; Liu, C.; Dong, H. Identification of a MYB3R gene involved in drought, salt and cold stress in wheat (Triticum aestivum L.). Gene 2011, 485, 146–152. [Google Scholar] [CrossRef]
- Ping, X.Y.; Zhou, L. Research prospects and progress on chilling injury of economic forest and fruit trees in south China. Chin. J. Eco-Agric. 2018, 26, 467–479. [Google Scholar]
- She, K.; Xu, Z.; Li, X.; Luo, X.; Liu, Y. Sequence characteristics and bioinformatics analysis of NCED gene in maize. Ningxia J. Agric. For. Sci. Technol. 2010, 5, 14–17. [Google Scholar]
- Sato, H.; Takasaki, H.; Takahashi, F.; Suzuki, T.; Iuchi, S.; Mitsuda, N.; Ohme-Takagi, M.; Ikeda, M.; Seo, M.; Yamaguchi-Shinozaki, K.; et al. Arabidopsis thaliana NGATHA1 transcription factor induces ABA biosynthesis by activating NCED3 gene during dehydration stress. Proc. Natl. Acad. Sci. USA 2018, 115, E11178–E11187. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.Z.; Tan, B.C. A distal ABA responsive element in AtNCED3 promoter is required for positive feedback regulation of ABA biosynthesis in arabidopsis. PLoS ONE 2014, 9, e87283. [Google Scholar] [CrossRef]
- Yang, J.; Guo, Z. Cloning of a 9-cis-epoxycarotenoid dioxygenase gene (SgNCED1) from Stylosanthes guianensis and its expression in response to abiotic stresses. Plant Cell Rep. 2007, 26, 1383–1390. [Google Scholar] [CrossRef]
- Nakashima, K.; Yamaguchi-Shinozaki, K. ABA signaling in stress-response and seed development. Plant Cell Rep. 2013, 32, 959–970. [Google Scholar] [CrossRef]
- Gao, Z.; Zhou, Y.; He, Y. Molecular epigenetic mechanisms for the memory of temperature stresses in plants. J. Genet. Genom. 2022, 49, 991–1001. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Gene Name | Gene ID | Chromosome Localization | Length (aa) | MW (kDa) | pI | Prediction of Subcellular Localization |
|---|---|---|---|---|---|---|---|
| Prunus mume | PmNCED1 | Pm005147 | Chr2 | 1496 | 168.43 | 5.83 | Nucleus |
| PmNCED2 | Pm005148 | Chr2 | 540 | 60.33 | 5.32 | Chloroplast | |
| PmNCED3 | Pm005153 | Chr2 | 563 | 62.25 | 6.53 | Mitochondrion | |
| PmNCED4 | Pm006647 | Chr2 | 586 | 64.73 | 6.21 | Peroxisome | |
| PmNCED5 | Pm006977 | Chr2 | 89 | 10.13 | 5.54 | Cell wall | |
| PmNCED6 | Pm008988 | Chr2 | 600 | 66.44 | 7.92 | Chloroplast | |
| PmNCED7 | Pm010425 | Chr3 | 617 | 68.18 | 6.43 | Chloroplast | |
| PmNCED8 | Pm011164 | Chr3 | 622 | 69.01 | 6.39 | Chloroplast | |
| PmNCED9 | Pm016267 | Chr5 | 463 | 52.58 | 6.45 | Peroxisome | |
| PmNCED10 | Pm017769 | Chr5 | 634 | 70.47 | 5.63 | Chloroplast | |
| Prunus apricot | PaNCED1 | PARG03953m01 | Chr2 | 600 | 66.43 | 7.92 | Chloroplast |
| PaNCED2 | PARG06146m02 | Chr2 | 586 | 64.69 | 6.21 | Peroxisome | |
| PaNCED3 | PARG06146m01 | Chr2 | 604 | 66.52 | 6.48 | Peroxisome | |
| PaNCED4 | PARG07910m01 | Chr2 | 565 | 62.47 | 6.20 | Mitochondrion | |
| PaNCED5 | PARG07915m02 | Chr2 | 520 | 58.21 | 5.76 | Cytoplasm | |
| PaNCED6 | PARG07915m01 | Chr2 | 267 | 30.14 | 5.44 | Nucleus | |
| PaNCED7 | PARG07916m01 | Chr2 | 1073 | 120.20 | 5.61 | Chloroplast | |
| PaNCED8 | PARG11107m01 | Chr3 | 627 | 69.57 | 6.63 | Chloroplast | |
| PaNCED9 | PARG11892m01 | Chr3 | 617 | 68.12 | 6.43 | Chloroplast | |
| PaNCED10 | PARG17947m01 | Chr5 | 615 | 68.35 | 5.69 | Chloroplast | |
| Prunus salicina | PsNCED1 | evm.model.LG01.539 | Chr1 | 597 | 65.77 | 6.21 | Peroxisome |
| PsNCED2 | evm.model.LG01.2322 | Chr1 | 565 | 62.55 | 6.36 | Chloroplast | |
| PsNCED3 | evm.model.LG01.2327 | Chr1 | 176 | 19.63 | 5.63 | Nucleus | |
| PsNCED4 | evm.model.LG01.2328 | Chr1 | 223 | 25.15 | 6.22 | Nucleus | |
| PsNCED5 | evm.model.LG01.2329 | Chr1 | 75 | 8.36 | 4.71 | Cytoplasm | |
| PsNCED6 | evm.model.LG01.2330 | Chr1 | 617 | 69.09 | 5.75 | Chloroplast | |
| PsNCED7 | evm.model.LG01.2331 | Chr1 | 586 | 65.92 | 5.55 | Cytoplasm | |
| PsNCED8 | evm.model.LG01.4261 | Chr1 | 605 | 66.82 | 7.93 | Chloroplast | |
| PsNCED9 | evm.model.LG02.1859 | Chr2 | 615 | 68.39 | 5.76 | Chloroplast | |
| PsNCED10 | evm.model.LG04.787 | Chr4 | 614 | 67.72 | 6.36 | Chloroplast | |
| PsNCED11 | evm.model.LG04.1420 | Chr4 | 627 | 69.64 | 7.34 | Chloroplast | |
| Prunus persica | PpNCED1 | transcript:ONI26993 | Chr1 | 605 | 66.95 | 7.24 | Chloroplast |
| PpNCED2 | transcript:ONI30522 | Chr1 | 608 | 66.81 | 6.29 | Peroxisome | |
| PpNCED3 | transcript:ONI30523 | Chr1 | 427 | 47.30 | 6.23 | Peroxisome | |
| PpNCED4 | transcript:ONI33831 | Chr1 | 565 | 62.50 | 6.33 | Chloroplast | |
| PpNCED5 | transcript:ONI33839 | Chr1 | 568 | 63.62 | 5.18 | Nucleus | |
| PpNCED6 | transcript:ONI33840 | Chr1 | 617 | 68.96 | 5.64 | Chloroplast | |
| PpNCED7 | transcript:ONI33841 | Chr1 | 555 | 62.49 | 6.01 | Cytoplasm | |
| PpNCED8 | transcript:ONI20423 | Chr2 | 430 | 49.12 | 6.09 | Cytoplasm | |
| PpNCED9 | transcript:ONI20422 | Chr2 | 547 | 61.83 | 5.97 | Peroxisome | |
| PpNCED10 | transcript:ONI22509 | Chr2 | 615 | 68.70 | 5.77 | Chloroplast | |
| PpNCED11 | transcript:ONI11006 | Chr4 | 617 | 68.16 | 6.43 | Chloroplast | |
| PpNCED12 | transcript:ONI12196 | Chr4 | 632 | 70.35 | 6.63 | Chloroplast | |
| Rosa chinensis | RcNCED1 | transcript:PRQ54664 | Chr1 | 494 | 56.04 | 6.56 | Peroxisome |
| RcNCED2 | transcript:PRQ54676 | Chr1 | 631 | 71.20 | 7.70 | Peroxisome | |
| RcNCED3 | transcript:PRQ56952 | Chr1 | 631 | 71.57 | 6.98 | Chloroplast | |
| RcNCED4 | transcript:PRQ57420 | Chr1 | 624 | 69.89 | 6.27 | Chloroplast | |
| RcNCED5 | transcript:PRQ57426 | Chr1 | 622 | 68.97 | 6.45 | Chloroplast | |
| RcNCED6 | transcript:PRQ36937 | Chr4 | 613 | 67.29 | 6.81 | Chloroplast | |
| RcNCED7 | transcript:PRQ41175 | Chr4 | 584 | 63.72 | 5.70 | Chloroplast | |
| RcNCED8 | transcript:PRQ29478 | Chr5 | 688 | 75.95 | 6.74 | Chloroplast | |
| RcNCED9 | transcript:PRQ30744 | Chr5 | 612 | 67.61 | 7.25 | Chloroplast | |
| RcNCED10 | transcript:PRQ27514 | Chr6 | 330 | 37.34 | 4.71 | Chloroplast | |
| RcNCED11 | transcript:PRQ27515 | Chr6 | 563 | 63.05 | 5.34 | Cytoplasm | |
| RcNCED12 | transcript:PRQ27522 | Chr6 | 561 | 62.10 | 6.04 | Chloroplast | |
| RcNCED13 | transcript:PRQ21333 | Chr7 | 303 | 34.62 | 5.56 | Peroxisome |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, K.; Li, X.; Guo, X.; Yang, L.; Qiu, L.; Liu, W.; Wang, J.; Zheng, T. Genome-Wide Identification and Expression Profiling of the NCED Gene Family in Cold Stress Response of Prunus mume Siebold & Zucc. Horticulturae 2023, 9, 839. https://doi.org/10.3390/horticulturae9070839
Chen K, Li X, Guo X, Yang L, Qiu L, Liu W, Wang J, Zheng T. Genome-Wide Identification and Expression Profiling of the NCED Gene Family in Cold Stress Response of Prunus mume Siebold & Zucc. Horticulturae. 2023; 9(7):839. https://doi.org/10.3390/horticulturae9070839
Chicago/Turabian StyleChen, Ke, Xue Li, Xiaoyu Guo, Lichen Yang, Like Qiu, Weichao Liu, Jia Wang, and Tangchun Zheng. 2023. "Genome-Wide Identification and Expression Profiling of the NCED Gene Family in Cold Stress Response of Prunus mume Siebold & Zucc" Horticulturae 9, no. 7: 839. https://doi.org/10.3390/horticulturae9070839