
Influence of Ripening Process on Pecan Nut (Carya illinoinensis) Kernel Quality: Phenolic Profile, Antioxidant Activity, and Carbohydrate Composition
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagent and Standard
2.2. Pecan Samples
2.3. Phenolic Extraction
2.4. Estimation of Total Phenols (TP)
2.5. Estimation of Total Flavonoids (TFoids)
2.6. Estimation of Total Condensed Tannins (TCT)
2.7. Determination of Total Flavanols (TFols)
2.8. Estimation of Total o-Diphenols (ToDs)
2.9. HPLC-DAD Analysis of Individual Phenolic Compounds
2.10. Estimation of Antioxidant Activity
2.10.1. DPPH (2,2-Diphenly-1-picrylhydrazyl) Radical Scavenging Activity
2.10.2. Ferric Reducing Antioxidant Power (FRAP)
2.11. Carbohydrate Analysis
2.12. Data Analysis
3. Results and Discussion
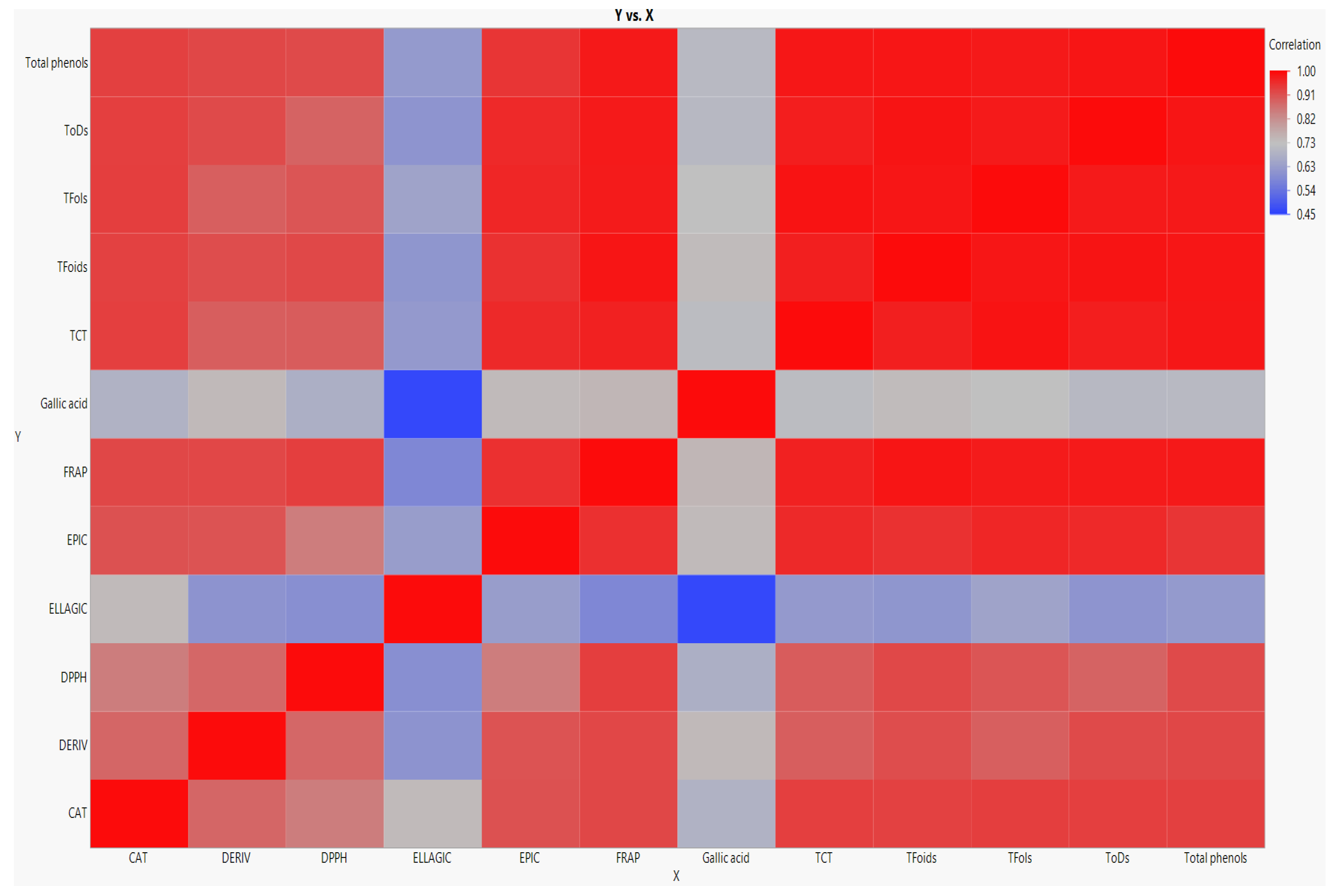
3.1. Variations in Different Classes of Phenolics during Pecan Nut Maturation
3.2. Variations in Individual Phenolic Compounds during Pecan Nut Maturation
3.3. Variations in Antioxidant Activity during Pecan Nut Maturation
3.4. Variations in Carbohydrates during Pecan Nut Development
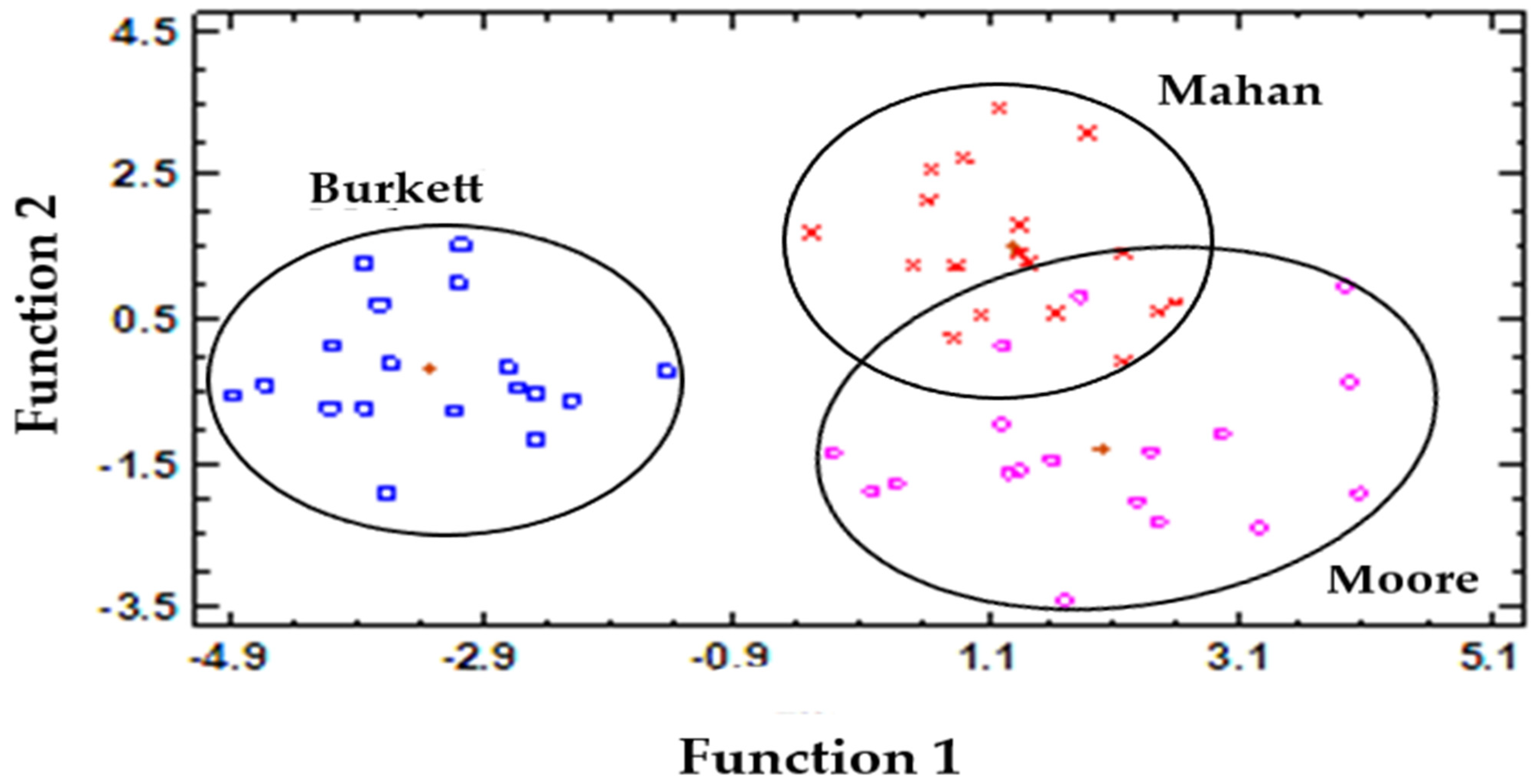
3.5. Canonical Discriminant Analysis (CDA)
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Gan, R.Y.; Li, S.; Zhou, Y.; Li, A.N.; Xu, D.P.; Li, H.B. Antioxidant Phytochemicals for the Prevention and Treatment of Chronic Diseases. Molecules 2015, 20, 21138–21156. [Google Scholar] [CrossRef] [PubMed]
- Ratnam, D.V.; Ankola, D.D.; Bhardwaj, V.; Sahana, D.K.; Kumar, M.N.V.R. Role of antioxidants in prophylaxis and therapy: A pharmaceutical perspective. J. Control. Release 2006, 113, 189–207. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.K.; Paniwnyk, L.; Hassan, S. Polyphenols as natural antioxidants: Sources, extraction and applications in food, cosmetics and drugs. In Plant Based “Green Chemistry 2.0”; Li, Y., Chemat, F., Eds.; Green Chemistry and Sustainable Technology; Springer: Singapore, 2019; pp. 197–235. [Google Scholar] [CrossRef]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic compounds as beneficial phytochemicals in pomegranate (Punica granatum L.) peel: A review. Food Chem. 2018, 261, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Bouali, I.; Tsafouros, A.; Ntanos, E.; Albouchi, A.; Boukhchina, S.; Roussos, P.A. Inter cultivar and temporal variation of phenolic compounds, antioxidant activity and carbohydrate composition of pecan (Carya illlinoinensis) kernels grown in Tunisia. Hortic. Environ. Biotechnol. 2020, 61, 183–196. [Google Scholar] [CrossRef]
- Hayes, J.E.; Allen, P.; Brunton, N.; O’Grady, M.N.; Kerry, J.P. Phenolic composition and in vitro antioxidant capacity of four commercial phytochemical products: Olive leaf extract (Olea europaea L.), lutein, sesamol and ellagic acid. Food Chem. 2011, 126, 948–955. [Google Scholar] [CrossRef]
- Colon, M.; Nerin, C. Role of catechins in the antioxidant capacity of an active film containing green tea, green coffee, and grapefruit extracts. J. Agric. Food Chem. 2012, 60, 9842–9849. [Google Scholar] [CrossRef]
- Rajan, V.K.; Muraleedharan, K. A computational investigation on the structure, global parameters and antioxidant capacity of a polyphenol, Gallic acid. Food Chem. 2017, 220, 93–99. [Google Scholar] [CrossRef]
- Atanosov, A.G.; Sabharanjak, S.M.; Zengin, G.; Mollica, A.; Szostak, A.; Simirgiotis, M.; Huminiecki, Ł.; Horbanczuk, O.K.; Nabavi, S.M.; Mocan, A. Pecan nuts: A review of reported bioactivities and health effects. Trends Food Sci. Technol. 2017, 71, 246–257. [Google Scholar] [CrossRef]
- Rajaram, S.; Burke, K.; Connell, K.; Myint, T.J.; Sabate, J. A monounsaturated fatty acid-rich pecan enriched diet favorably alters the serum lipid profile of healthy men and women. J. Nutr. 2001, 131, 2275–2279. [Google Scholar] [CrossRef]
- Kornsteiner, M.; Wagner, K.H.; Elmadfa, I. Tocopherols and total phenolics in 10 different nut types. Food Chem. 2006, 98, 381–387. [Google Scholar] [CrossRef]
- Tsantili, E.; Konstantinidis, K.; Christopoulos, M.V.; Roussos, P.A. Total phenolics and flavonoids and total antioxidant capacity in pistachio (Pistachia vera L.) nuts in relation to cultivars and storage conditions. Sci. Hortic. 2011, 129, 694–701. [Google Scholar] [CrossRef]
- Bamdad, F.; Kadivar, M.; Keramat, J. Evaluation of phenolic content and antioxidant activity of Iranian caraway in comparison with clove and BHT using model systems and vegetable oil. Int. J. Food. Sci. Technol. 2006, 41, 20–27. [Google Scholar] [CrossRef]
- de la Rosa, L.A.; Alvarez-Parrilla, E.; Shahidi, F. Phenolic compounds and antioxidant activity of kernels and shells of Mexican pecan (Carya illinoinensis). J. Agric. Food Chem. 2011, 59, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Arnous, A.; Makris, D.P.; Kefalas, P. Correlation of pigment and flavanol content with antioxidant properties in selected aged regional wines from Greece. J. Food Compos. Anal. 2002, 15, 655–665. [Google Scholar] [CrossRef]
- Roussos, P.A.; Pontikis, C.A. Phenolic compounds in olive explants and their contribution to browning during the establishment stage in vitro. Gartenbauwissenschaft 2001, 66, 298–303. [Google Scholar]
- Roussos, P.A.; Pontikis, C.A. Changes of free, soluble conjugated and bound polyamine titers of jojoba explants under sodium chloride salinity in vitro. J. Plant Physiol. 2007, 164, 895–903. [Google Scholar] [CrossRef]
- Ferreira, I.C.F.R.; Baptista, P.; Vilas-Boas, M.; Barros, L. Free-radical scavenging capacity and reducing power of wild edible mushrooms from northeast Portugal: Individual cap and stipe activity. Food Chem. 2007, 100, 1511–1516. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Meth. Enzymol. 1999, 299, 15–27. [Google Scholar] [CrossRef]
- Roussos, P.A.; Vemmos, S.N.; Pontikis, C.A. The role of carbohydrates on the salt tolerance of Jojoba [Simmondsia chinensis (Link)] expiants in vitro. Eur. J. Hortic. Sci. 2005, 70, 278–282. [Google Scholar]
- Jia, X.; Luo, H.; Xu, M.; Zhai, M.; Guo, Z.; Qiao, Y.; Wang, L. Dynamic changes in phenolics and antioxidant capacity during pecan (Carya illinoinensis) kernel ripening and its phenolics profiles. Molecules 2018, 23, 435. [Google Scholar] [CrossRef] [PubMed]
- Amin, F.; Masoodi, F.A.; Baba, W.N.; Khan, A.A.; Ganie, B.A. Effect of different ripening stages on walnut kernel quality: Antioxidant activities, lipid characterization and antibacterial properties. J. Food Sci. Technol. 2017, 54, 3791–3801. [Google Scholar] [CrossRef] [PubMed]
- Persic, M.; Mikulic-Petkovsek, M.; Slatnar, A.; Solar, A.; Veberic, R. Changes in phenolic profiles of red-colored pellicle walnut and hazelnut kernel during ripening. Food Chem. 2018, 252, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Pycia, K.; Kapusta, I.; Jaworska, G. Impact of the degree of maturity of walnuts (Juglans regia L.) and their variety on the antioxidant potential and the content of tocopherols and polyphenols. Molecules 2019, 24, 2936. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Li, Y.; Sun, D.; Chen, Q.; Fu, M.; Zhao, H.; Chen, X.; Huang, Y.; Xu, H. Odor, tastes, nutritional compounds and antioxidant activity of fresh-eating walnut during ripening. Sci. Hortic. 2022, 293, 110744. [Google Scholar] [CrossRef]
- Lopes, M.M.A.; Miranda, M.R.A.; Moura, C.F.H.; Filho, J.E. Bioactive compounds and total antioxidant capacity of cashew apples (Anacardium occidentale L.) during the ripening of early dwarf cashew clones. Ciênc. Agrotecnol. 2012, 36, 325–332. [Google Scholar] [CrossRef]
- Dabbou, S.; Lussiana, C.; Maatallah, S.; Gasco, L.; Hajlaoui, H.; Flamini, G. Changes in biochemical compounds in flesh and peel from Prunus persica fruits grown in Tunisia during two maturation stages. Plant Physiol. Biochem. 2016, 100, 1–11. [Google Scholar] [CrossRef]
- Ferrari, V.; Gil, G.; Heinzen, H.; Zoppolo, R.; Ibáñez, F. Influence of Cultivar on Nutritional Composition and Nutraceutical Potential of Pecan Growing in Uruguay. Front. Nutr. 2022, 9, 868054. [Google Scholar] [CrossRef]
- Prabhakar, H.; Sharma, S.; Kong, F. Effects of Postharvest Handling and Storage on Pecan Quality. Food Rev. Int. 2020, 38, 1485–1512. [Google Scholar] [CrossRef]
- Ballistreri, G.; Arena, E.; Fallico, B. Influence of ripeness and drying process on the polyphenols and tocopherols of Pistacia vera L. Molecules 2009, 14, 4358–4369. [Google Scholar] [CrossRef]
- Guizani, M.; Maatallah, S.; Dabbou, S.; Serrano, M.; Hajlaoui, H.; Helal, A.N.; Kilani-Jaziri, S. Physiological behaviors and fruit quality changes in five peach cultivars during three ripening stages in a semi-arid climate. Acta Physiol. Plant. 2019, 41, 154. [Google Scholar] [CrossRef]
- Bizjak, J.; Mikulic-Petkovsek, M.; Stampar, F.; Veberic, R. Changes in primary metabolites and polyphenols in the peel of “Braeburn” apples (Malus domestica Borkh.) during advanced maturation. J. Agric. Food Chem. 2013, 61, 10283–10292. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.; Felizardo, C.; Fernandes-Silva, A.A.; Nunes, F.M.; Barros, A. Polyphenolic compounds, antioxidant activity and l-phenylalanine ammonia-lyase activity during ripening of olive cv. “Cobrançosa” under different irrigation regimes. Food Res. Int. 2013, 51, 412–421. [Google Scholar] [CrossRef]
- Belhadj, F.; Somrani, I.; Aissaoui, N.; Messaoud, C.; Boussaid, M.; Marzouki, M.N. Bioactive compounds contents, antioxidant and antimicrobial activities during ripening of Prunus persica L. varieties from the North West of Tunisia. Food Chem. 2016, 204, 29–36. [Google Scholar] [CrossRef]
- Rauf, A.; Imran, M.; Abu-Izneid, T.; Ul-Haq, I.; Patel, S.; Pan, X.; Naz, S.; Silva, A.S.; Saeed, F.; Suleria, H.A.R. Proanthocyanidins: A comprehensive review. Biomed. Pharmacother. 2019, 116, 108999. [Google Scholar] [CrossRef]
- Assis, J.S.; Maldonado, R.; Muñoz, T.; Escribano, M.I.; Merodio, C. Effect of high carbon dioxide concentration on PAL activity and phenolic contents in ripening cherimoya fruit. Postharvest Biol. Technol. 2001, 23, 33–39. [Google Scholar] [CrossRef]
- Downey, M.O.; Harvey, J.S.; Robinson, S.P. Analysis of tannins in seeds and skins of Shiraz grapes throughout berry development. Aust. J. Grape Wine Res. 2003, 9, 15–27. [Google Scholar] [CrossRef]
- Zeng, X.; Du, Z.; Sheng, Z.; Jiang, W. Characterization of the interactions between banana condensed tannins and biologically important metal ions (Cu2+, Zn2+ and Fe2+). Food Res. Int. 2019, 123, 518–528. [Google Scholar] [CrossRef]
- Zarei, M.; Davarynejad, G.; Abedi, B.; Kafi, M.; Biabani, A. Changes in physical properties, chemical composition and antioxidant activity of four pistachio cultivars at ten maturity stages. Adv. Environ. Biol. 2014, 8, 106–115. [Google Scholar]
- Villarreal-Lozoya, J.E.; Lombardini, L.; Cisneros-Zevallos, L. Phytochemical constituents and antioxidant capacity of different pecan [Carya illinoinensis (Wangenh.) K. Koch] cultivars. Food Chem. 2007, 102, 1241–1249. [Google Scholar] [CrossRef]
- Flores-Córdova, M.A.; Sánchez, E.; Muñoz-Márquez, E.; Ojeda-Barrios, D.L.; Soto-Parra, J.M.; Preciado-Rangel, P. Phytochemical composition and antioxidant capacity in Mexican pecan nut. Emir. J. Food Agric. 2017, 29, 346–350. [Google Scholar] [CrossRef]
- Martín, M.Á.; Fernández-Millán, E.; Ramos, S.; Bravo, L.; Goya, L. Cocoa flavonoid epicatechin protects pancreatic beta cell viability and function against oxidative stress. Mol. Nutr. Food Res. 2013, 58, 447–456. [Google Scholar] [CrossRef]
- Qu, Z.; Liu, A.; Li, P.; Liu, C.; Xiao, W.; Huang, J.; Liu, Z.; Zhang, S. Advances in physiological functions and mechanisms of (−)-epicatechin. Crit. Rev. Food Sci. Nutr. 2020, 61, 211–233. [Google Scholar] [CrossRef] [PubMed]
- Bernatova, I. Biological activities of (−)-epicatechin and (−)-epicatechin-containing foods: Focus on cardiovascular and neuropsychological health. Biotechnol. Adv. 2018, 36, 666–681. [Google Scholar] [CrossRef] [PubMed]
- Seraglio, S.K.T.; Schulz, M.; Nehring, P.; Betta, F.D.; Valese, A.C.; Daguer, H.; Gonzaga, L.V.; Fett, R.; Costa, A.C.O. Nutritional and bioactive potential of Myrtaceae fruits during ripening. Food Chem. 2018, 239, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Gruz, J.; Ayaz, F.A.; Torun, H.; Strnad, M. Phenolic acid content and radical scavenging activity of extracts from medlar (Mespilus germanica L.) fruit at different stages of ripening. Food Chem. 2011, 124, 271–277. [Google Scholar] [CrossRef]
- Bashir, H.A.; Abu-Goukh, A.B.A. Compositional changes during guava fruit ripening. Food Chem. 2003, 80, 557–563. [Google Scholar] [CrossRef]
- Fu, M.; Qu, Q.; Yang, X.; Zhang, X. Effect of intermittent oven drying on lipid oxidation, fatty acids composition and antioxidant activities of walnut. LWT Food Sci. Technol. 2016, 65, 1126–1132. [Google Scholar] [CrossRef]
- Yang, J.; Liu, R.H.; Halim, L. Antioxidant and antiproliferative activities of common edible nut seeds. LWT Food Sci. Technol. 2009, 42, 1–8. [Google Scholar] [CrossRef]
- Singanusong, R.; Mason, R.L.; D’Arcy, B.R.; Nottingham, S.M. Compositional changes of Australia-grown Western Schley pecans [Carya illinoinensis (Wangenh.) K. Koch] during Maturation. J. Agric. Food Chem. 2003, 51, 406–412. [Google Scholar] [CrossRef]
- Wood, B.W.; McMeans, J.L. Carbohydrates and fatty acids in developing pecan fruit. J. Am. Soc. Hortic. Sci. 1982, 107, 47–50. [Google Scholar] [CrossRef]
- Yang, B.M.; Yao, L.X.; Li, G.L.; He, Z.H.; Zhou, C.M. Dynamic changes of nutrition in litchi foliar and effects of potassium–nitrogen fertilization ratio. J. Soil Sci. Plant Nutr. 2015, 15, 98–110. [Google Scholar] [CrossRef]
- Cristofori, V.; Bertazza, G.; Bignami, C. Changes in kernel chemical composition during nut development of three Italian hazelnut cultivars. Fruits 2015, 70, 311–322. [Google Scholar] [CrossRef]
- Carvalho, L.C.; Morais, C.L.M.; Lima, K.M.G.; Leite, G.W.P.; Oliveira, G.S.; Casagrande, I.P.; Neto, J.P.S.; Teixeira, G.H.A. Using Intact Nuts and Near Infrared Spectroscopy to Classify Macadamia Cultivars. Food Anal. Methods 2017, 11, 1857–1866. [Google Scholar] [CrossRef]
- Cristofori, V.; Ferramondo, S.; Bertazza, G.; Bignami, C. Nut and kernel traits and chemical composition of hazelnut (Corylus avellana L.) cultivars. J. Sci. Food Agric. 2008, 88, 1091–1098. [Google Scholar] [CrossRef]
- Amorello, D.; Orecchio, S.; Pace, A.; Barreca, S. Discrimination of almonds (Prunus dulcis) geographical origin by minerals and fatty acids profiling. Nat. Prod. Res. 2015, 30, 2107–2110. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Nut Maturity Stage | Sampling Date | Weeks after Flowering Date (WAFD) | |
|---|---|---|---|
| Stage 1 | Unripe stage | 24.09 | 20 |
| 01.10 | 21 | ||
| 08.10 | 22 | ||
| Stage 2 | Half-ripe stage | 15.10 | 23 |
| 22.10 | 24 | ||
| Stage 3 | Fully ripe stage | 29.10 | 25 |
| Parameters | Total Phenols (mg GAE g−1 FW) | Total Flavonoids (mg CE g−1 FW) | Total Flavanols (mg CE g−1 FW) | Total Condensed Tannins (mg CE g−1 FW) | Total o-Diphenols (mg CAE g−1 FW) | |
|---|---|---|---|---|---|---|
| Cultivar | Mahan | 16.40 c | 11.18 c | 4.44 c | 65.31 c | 2.15 c |
| Moore | 21.42 b | 14.81 b | 5.08 b | 79.42 b | 3.41 b | |
| Burkett | 26.29 a | 17.46 a | 7.09 a | 102.57 a | 5.00 a | |
| WAFD | 20 | 57.92 a | 37.07 a | 14.76 a | 221.13 a | 11.05 a |
| 21 | 13.35 c | 9.33 c | 3.95 bc | 58.07 bc | 1.66 c | |
| 22 | 13.70 c | 8.70 c | 3.60 cd | 59.36 bc | 1.79 c | |
| 23 | 13.36 c | 8.84 c | 2.83 d | 48.19 cd | 2.05 bc | |
| 24 | 17.63 b | 13.46 b | 4.73 b | 65.85 b | 2.46 b | |
| 25 | 12.25 c | 9.56 c | 3.61 d | 41.98 d | 2.12 bc | |
| Cultivar × WAFD | Mahan × 20 | 16.53 ± 2.13 b | 9.71 ± 1.35 c | 4.45 ± 0.76 b | 63.89 ± 4.19 b | 2.01 ± 0.21 bc |
| ×21 | 15.75 ± 1.88 b | 10.63 ± 0.88 c | 4.53 ± 0.58 b | 64.44 ± 6.50 b | 1.95 ± 0.19 bc | |
| ×22 | 22.61 ± 3.93 a | 12.74 ± 1.63 a | 5.91 ± 1.02 a | 95.00 ± 16.53 a | 2.80 ± 0.62 a | |
| ×23 | 15.71 ± 1.46 b | 10.02 ± 1.83 c | 3.39 ± 0.34 c | 52.85 ± 6.56 c | 1.94 ± 0.42 bc | |
| ×24 | 13.83 ± 2.37 c | 11.67 ± 2.24 b | 4.19 ± 0.61 b | 58.65 ± 4.55 bc | 1.72 ± 0.33 c | |
| ×25 | 13.92 ± 0.76 c | 12.34 ± 2.91 a | 4.19 ± 1.34 b | 57.04 ± 12.96 bc | 2.51 ± 0.69 b | |
| Moore × 20 | 54.64 ± 6.60 a | 34.70 ± 4.66 a | 12.36 ± 1.58 a | 186.13 ± 19.28 a | 9.98 ± 1.19 a | |
| ×21 | 15.01 ± 0.76 c | 10.98 ± 2.47 c | 4.32 ± 0.96 bc | 71.29 ± 11.06 c | 1.75 ± 0.29 c | |
| ×22 | 8.37 ± 1.73 d | 5.56 ± 1.22 d | 1.75 ± 0.51 c | 33.72 ± 8.31 d | 1.05 ± 0.15 c | |
| ×23 | 13.09 ± 3.23 c | 9.05 ± 1.98 c | 2.90 ± 0.66 c | 60.68 ± 12.48 c | 1.53 ± 0.46 c | |
| ×24 | 26.82 ± 0.56 b | 20.52 ± 1.33 b | 6.86 ± 0.45 b | 91.89 ± 14.27 b | 4.05 ± 0.18 b | |
| ×25 | 10.59 ± 2.02 d | 8.19 ± 0.45 c | 2.27 ± 0.17 c | 32.79 ± 3.15 d | 2.08 ± 0.42 c | |
| Burkett × 20 | 102.59 ± 10.25 a | 66.81 ± 3.15 a | 27.48 ± 1.55 a | 413.37 ± 39.01 a | 21.15 ± 1.44 a | |
| ×21 | 9.29 ± 2.28 c | 6.37 ± 1.82 c | 3.01 ± 0.92 b | 38.50 ± 6.88 bc | 1.28 ± 0.37 c | |
| ×22 | 10.11 ± 0.99 b | 7.82 ± 0.66 b | 3.14 ± 0.32 b | 49.37 ± 4.00 b | 1.50 ± 0.17 c | |
| ×23 | 11.23 ± 2.21 b | 7.46 ± 1.58 b | 2.19 ± 0.36 c | 31.04 ± 3.50 c | 2.70 ± 0.27 b | |
| ×24 | 12.25 ± 0.65 b | 8.18 ± 0.51 b | 3.16 ± 0.17 b | 47.01 ± 3.50 b | 1.61 ± 0.16 c | |
| ×25 | 12.25 ± 1.17 b | 8.17 ± 1.48 b | 3.60 ± 0.19 b | 36.12 ± 3.14 bc | 1.77 ± 0.09 c |
| Parameters | Gallic Acid (µg g−1 FW) | Catechin (µg g−1 FW) | Epicatechin (µg g−1 FW) | Ellagic Acid Derivative (µg g−1 FW) | Ellagic Acid (µg g−1 FW) | |
|---|---|---|---|---|---|---|
| Cultivar | Mahan | 18.99 b | 430.25 c | 17.12 c | 214.45 c | 448.76 a |
| Moore | 27.94 a | 570.01 b | 49.07 b | 414.48 a | 346.23 b | |
| Burkett | 20.59 b | 798.99 a | 68.67 a | 352.16 b | 396.12 ab | |
| WAFD | 20 | 34.05 a | 1883.20 a | 190.22 a | 1046.26 a | 807.05 a |
| 21 | 22.84 bc | 760.80 b | 17.21 b | 148.41 d | 604.88 b | |
| 22 | 15.35 c | 328.42 c | 13.31 b | 117.13 d | 257.19 cd | |
| 23 | 16.23 c | 216.93 c | 14.46 b | 279.53 b | 167.99 d | |
| 24 | 25.61 b | 240.28 c | 22.83 b | 211.47 c | 311.21 c | |
| 25 | 20.95 bc | 168.89 c | 11.69 b | 159.39 cd | 233.88 cd | |
| Cultivar × WAFD | Mahan × 20 | 12.60 ± 0.94 c | 543.23 ± 82.32 b | 20.86 ± 8.05 b | 166.69 ± 44.41 c | 624.00 ± 82.25 b |
| ×21 | 18.16 ± 4.01 b | 898.61 ± 231.30 a | 18.60 ± 6.73 b | 159.93 ± 36.15 c | 965.37 ± 249.00 a | |
| ×22 | 16.56 ± 2.99 bc | 433.05 ± 86.05 c | 15.19 ± 4.33 c | 120.29 ± 17.89 c | 239.64 ± 58.18 cd | |
| ×23 | 19.11 ± 2.35 b | 244.27 ± 53.79 d | 22.41 ± 8.95 a | 487.38 ± 16.14 a | 160.85 ± 15.78 d | |
| ×24 | 33.96 ± 11.77 a | 236.45 ± 78.17 d | 16.40 ± 6.96 c | 298.69 ± 42.01 b | 469.10 ± 57.81 c | |
| ×25 | 13.55 ± 5.21 c | 225.88 ± 42.60 d | 9.26 ± 2.50 d | 53.74 ± 15.36 d | 233.61 ± 36.96 cd | |
| Moore ×20 | 36.20 ± 10.53 a | 1733.73 ± 224.12 a | 158.96 ± 17.10 a | 1316.00 ± 110.14 a | 784.35 ± 185.93 a | |
| ×21 | 32.57 ± 4.27 b | 896.08 ± 72.46 b | 28.22 ± 6.38 c | 228.96 ± 26.80 c | 310.90 ± 75.58 b | |
| ×22 | 13.42 ± 4.79 d | 104.94 ± 21.87 d | 19.70 ± 8.68 cd | 140.14 ± 37.88 d | 242.66 ± 55.79 bc | |
| ×23 | 19.83 ± 5.35 c | 244.68 ± 18.17 c | 16.20 ± 8.51 d | 247.10 ± 37.95 c | 117.84 ± 33.32 c | |
| ×24 | 29.94 ± 3.84 b | 296.33 ± 17.67 c | 48.55 ± 11.72 b | 248.98 ± 63.36 c | 314.69 ± 19.77 b | |
| ×25 | 35.69 ± 6.48 a | 144.31 ± 48.73 d | 22.78 ± 6.82 cd | 305.68 ± 80.33 b | 306.97 ± 63.48 b | |
| Burkett ×20 | 53.36 ± 5.38 a | 3372.64 ± 459.81 a | 390.83 ± 88.51 a | 1656.09 ± 21.64 a | 1012.81 ± 120.80 a | |
| ×21 | 17.81 ± 0.97 b | 487.71 ± 94.59 b | 4.80 ± 1.18 bc | 56.33 ± 6.07 d | 538.38 ± 85.30 b | |
| ×22 | 16.09 ± 4.78 b | 447.27 ± 83.63 b | 5.05 ± 1.46 b | 90.97 ± 28.64 c | 289.27 ± 28.10 c | |
| ×23 | 9.76 ± 2.61 c | 161.83 ± 13.91 c | 4.77 ± 1.77 bc | 104.11 ± 32.44 bc | 225.30 ± 24.67 cd | |
| ×24 | 12.92 ± 1.50 bc | 188.05 ± 33.36 c | 3.53 ± 0.35 c | 86.73 ± 10.65 c | 149.85 ± 12.02 d | |
| ×25 | 13.62 ± 2.03 bc | 136.47 ± 18.70 c | 3.04 ± 0.36 c | 118.75 ± 39.68 b | 161.08 ± 39.54 d |
| Parameters | Antioxidant Activity (Expressed as μmol TE g−1 FW) | ||
|---|---|---|---|
| FRAP | DPPH | ||
| Cultivar | Mahan | 114.04 b | 195.96 b |
| Moore | 166.26 a | 211.95 a | |
| Burkett | 179.78 a | 176.76 c | |
| WAFD | 20 | 391.96 a | 344.28 a |
| 21 | 100.89 bc | 162.45 c | |
| 22 | 96.62 c | 159.47 c | |
| 23 | 103.05 bc | 162.44 c | |
| 24 | 126.77 b | 189.25 b | |
| 25 | 100.88 bc | 151.45 c | |
| Cultivar × WAFD | Mahan × 20 | 92.75 ± 14.71 c | 198.60 ± 20.47 b |
| ×21 | 101.52 ± 10.96 bc | 195.67 ± 11.38 b | |
| ×22 | 159.05 ± 29.20 a | 229.87 ± 20.10 a | |
| ×23 | 119.48 ± 10.68 b | 191.88 ± 7.80 b | |
| ×24 | 92.45 ± 13.91 c | 168.50 ± 10.39 c | |
| ×25 | 119.01 ± 42.92 b | 191.24 ± 39.20 b | |
| Moore × 20 | 396.17 ± 85.57 a | 382.41 ± 30.80 a | |
| ×21 | 131.81 ± 27.34 c | 181.07 ± 16.09 c | |
| ×22 | 64.04 ± 13.67 d | 137.47 ± 12.98 d | |
| ×23 | 110.65 ± 29.12 c | 176.57 ± 21.79 c | |
| ×24 | 200.85 ± 49.08 b | 263.97 ± 8.93 b | |
| ×25 | 94.05 ± 11.87 cd | 130.20 ± 12.78 d | |
| Burkett × 20 | 686.95 ± 18.06 a | 451.82 ± 12.99 a | |
| ×21 | 69.36 ± 20.07 c | 110.62 ± 18.47 c | |
| ×22 | 66.79 ± 3.27 c | 111.07 ± 17.43 c | |
| ×23 | 79.01 ± 16.11 bc | 118.87 ± 19.58 c | |
| ×24 | 87.00 ± 5.59 b | 135.29 ± 27.66 b | |
| ×25 | 89.58 ± 7.39 b | 132.90 ± 15.52 b | |
| Parameters | Sucrose | Glucose | Fructose | Total | |
|---|---|---|---|---|---|
| Cultivar | Mahan | 18.57 a | 7.48 a | 7.34 4 b | 33.39 ab |
| Moore | 17.90 ab | 6.88 a | 7.36 b | 32.14 b | |
| Burkett | 14.89 b | 8.55 a | 10.58 a | 34.01 a | |
| WAFD | 20 | 30.79 a | 15.16 a | 23.50 a | 69.45 a |
| 21 | 6.09 e | 9.95 b | 8.89 b | 24.93 c | |
| 22 | 6.57 e | 6.35 bc | 4.83 bcd | 17.76 d | |
| 23 | 12.30 d | 9.09 b | 8.16 bc | 29.54 bc | |
| 24 | 20.80 c | 1.45 d | 2.01 d | 24.26 c | |
| 25 | 26.16 b | 3.82 cd | 3.16 cd | 33.14 b | |
| Cultivar × WAFD | Mahan × 20 | 32.56 ± 0.68 a | 5.46 ± 0.68 c | 7.68 ± 1.06 b | 45.70 a |
| ×21 | 6.32 ± 1.74 e | 8.06 ± 0.61 bc | 6.48 ± 0.38 bc | 20.86 d | |
| ×22 | 6.05 ± 2.84 e | 9.48 ± 0.36 b | 7.27 ± 0.61 b | 22.80 d | |
| ×23 | 16.00 ± 6.56 d | 14.21± 6.22 a | 15.10 ± 9.75 a | 45.31 a | |
| ×24 | 20.84 ± 1.58 c | 2.35 ± 2.23 d | 2.85 ± 2.13 d | 26.04 c | |
| ×25 | 29.65 ± 3.44 b | 5.33 ± 0.14 c | 4.64 ± 1.28 cd | 39.62 b | |
| Moore × 20 | 29.42 ± 4.53 a | 14.00 ± 1.25 a | 23.67 ± 0.37 a | 67.09 a | |
| ×21 | 5.99 ± 0.22 d | 10.36 ± 0.23 b | 8.02 ± 0.45 b | 24.37 c | |
| ×22 | 8.21 ± 2.04 c | 4.56 ± 0.30 c | 2.85 ± 0.27 d | 15.62 d | |
| ×23 | 7.56 ± 1.24 c | 7.20 ± 0.43 bc | 4.81 ± 0.14 c | 19.57 cd | |
| ×24 | 28.43 ±0.58 b | 0.94 ± 0.60 d | 1.37 ± 0.06 e | 30.74 bc | |
| ×25 | 27.80 ± 1.75 b | 4.23 ± 1.15 c | 3.43 ± 1.30 cd | 35.46 b | |
| Burkett × 20 | 30.40 ± 1.82 a | 26.03 ± 2.04 a | 39.14 ± 0.83 a | 95.57 a | |
| ×21 | 5.96 ± 0.06 d | 11.43 ± 6.06 b | 12.18 ± 6.51 b | 29.57 b | |
| ×22 | 5.47 ± 0.43 d | 5.02 ± 0.24 c | 4.37 ± 1.10 c | 14.86 d | |
| ×23 | 13.33 ± 1.72 c | 5.85 ± 1.52 c | 4.56 ± 0.88 c | 23.74 bc | |
| ×24 | 13.12 ± 7.78 c | 1.07 ± 0.54 d | 1.81 ± 0.17 d | 16.00 d | |
| ×25 | 21.04 ± 9.05 b | 1.91 ± 0.1 d | 1.40 ± 0.01 d | 24.35 bc |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouali, I.; Tsafouros, A.; Ntanos, E.; Albouchi, A.; Boukhchina, S.; Roussos, P.A. Influence of Ripening Process on Pecan Nut (Carya illinoinensis) Kernel Quality: Phenolic Profile, Antioxidant Activity, and Carbohydrate Composition. Horticulturae 2023, 9, 1093. https://doi.org/10.3390/horticulturae9101093
Bouali I, Tsafouros A, Ntanos E, Albouchi A, Boukhchina S, Roussos PA. Influence of Ripening Process on Pecan Nut (Carya illinoinensis) Kernel Quality: Phenolic Profile, Antioxidant Activity, and Carbohydrate Composition. Horticulturae. 2023; 9(10):1093. https://doi.org/10.3390/horticulturae9101093
Chicago/Turabian StyleBouali, Intidhar, Athanassios Tsafouros, Efstathios Ntanos, Ali Albouchi, Sadok Boukhchina, and Peter A. Roussos. 2023. "Influence of Ripening Process on Pecan Nut (Carya illinoinensis) Kernel Quality: Phenolic Profile, Antioxidant Activity, and Carbohydrate Composition" Horticulturae 9, no. 10: 1093. https://doi.org/10.3390/horticulturae9101093