

Insight into the Phylogenetic Relationships and Evolutionary History of Pepper Cultivars (Capsicum annuum L.) through Comparative Analyses of Plastomes

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and DNA Extraction
2.2. DNA Sequencing, Genome Assembly, and Annotation
2.3. Comparative Plastome Analysis
2.4. Phylogenetic Analysis
3. Results
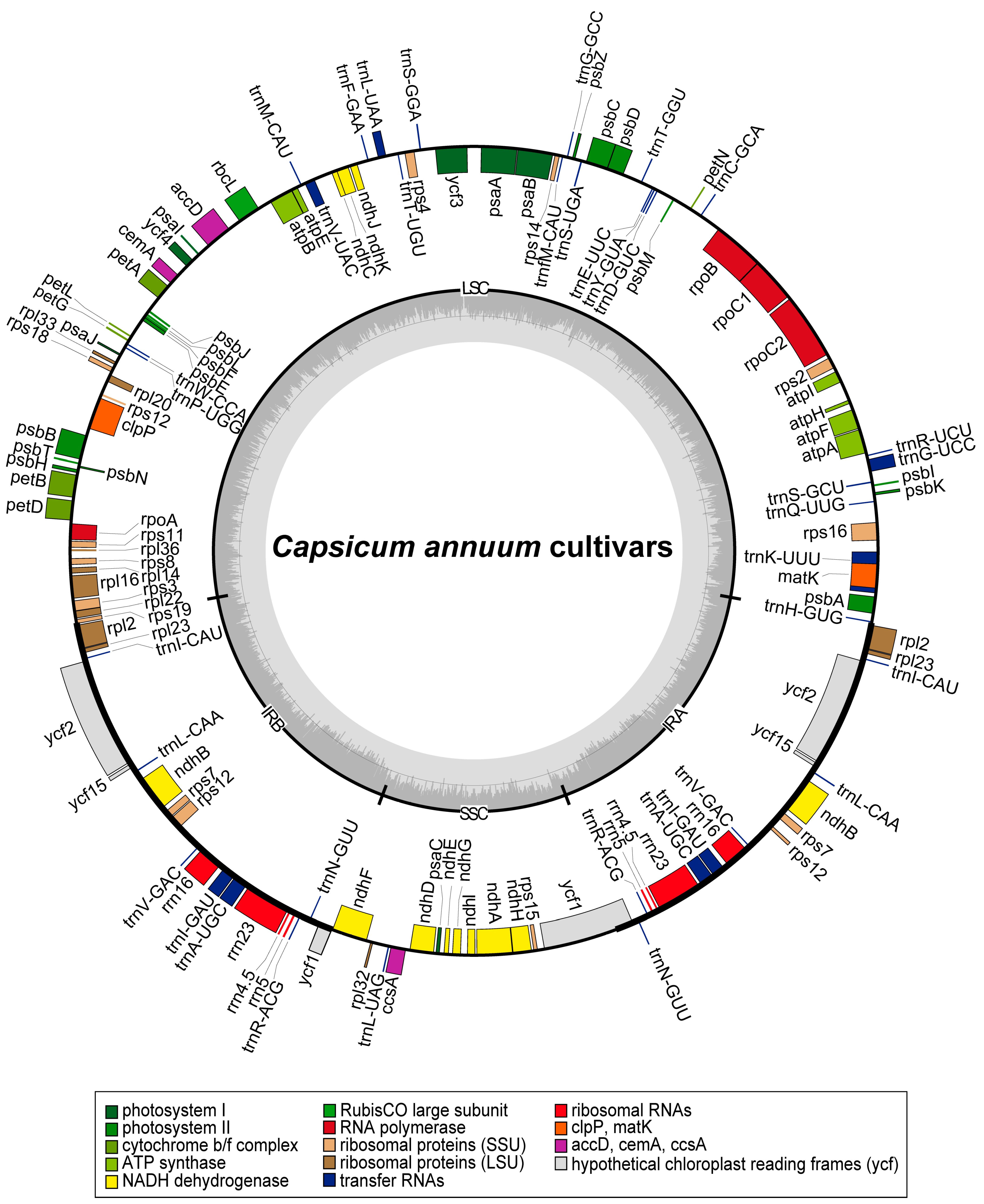
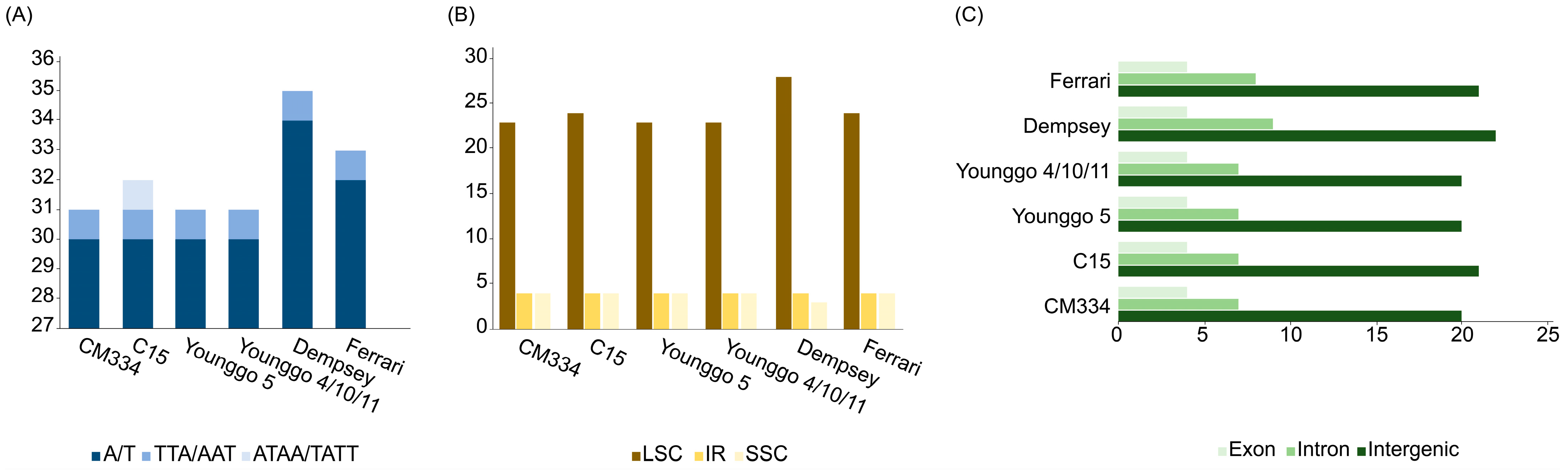
3.1. Plastome Organization and Features
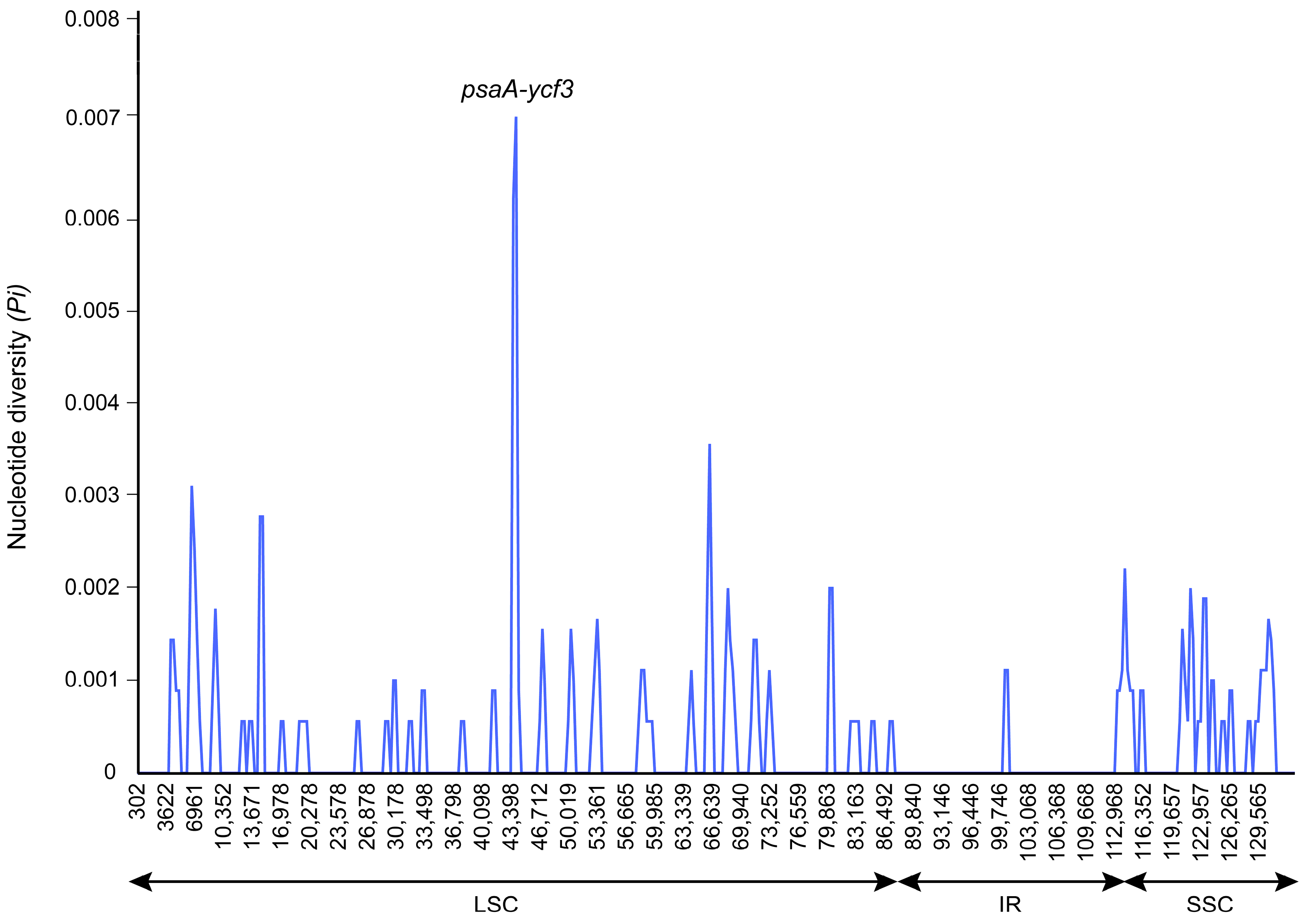
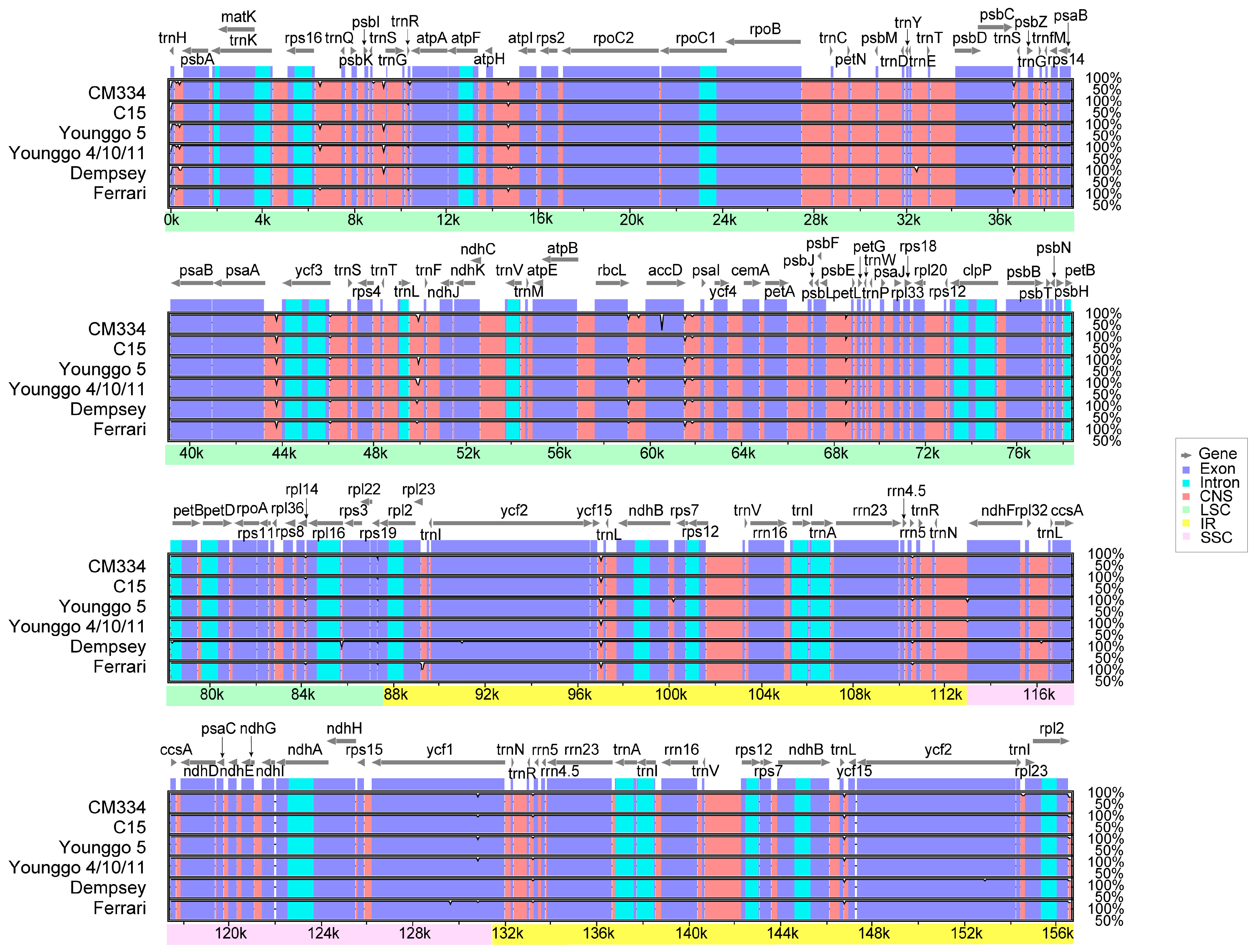
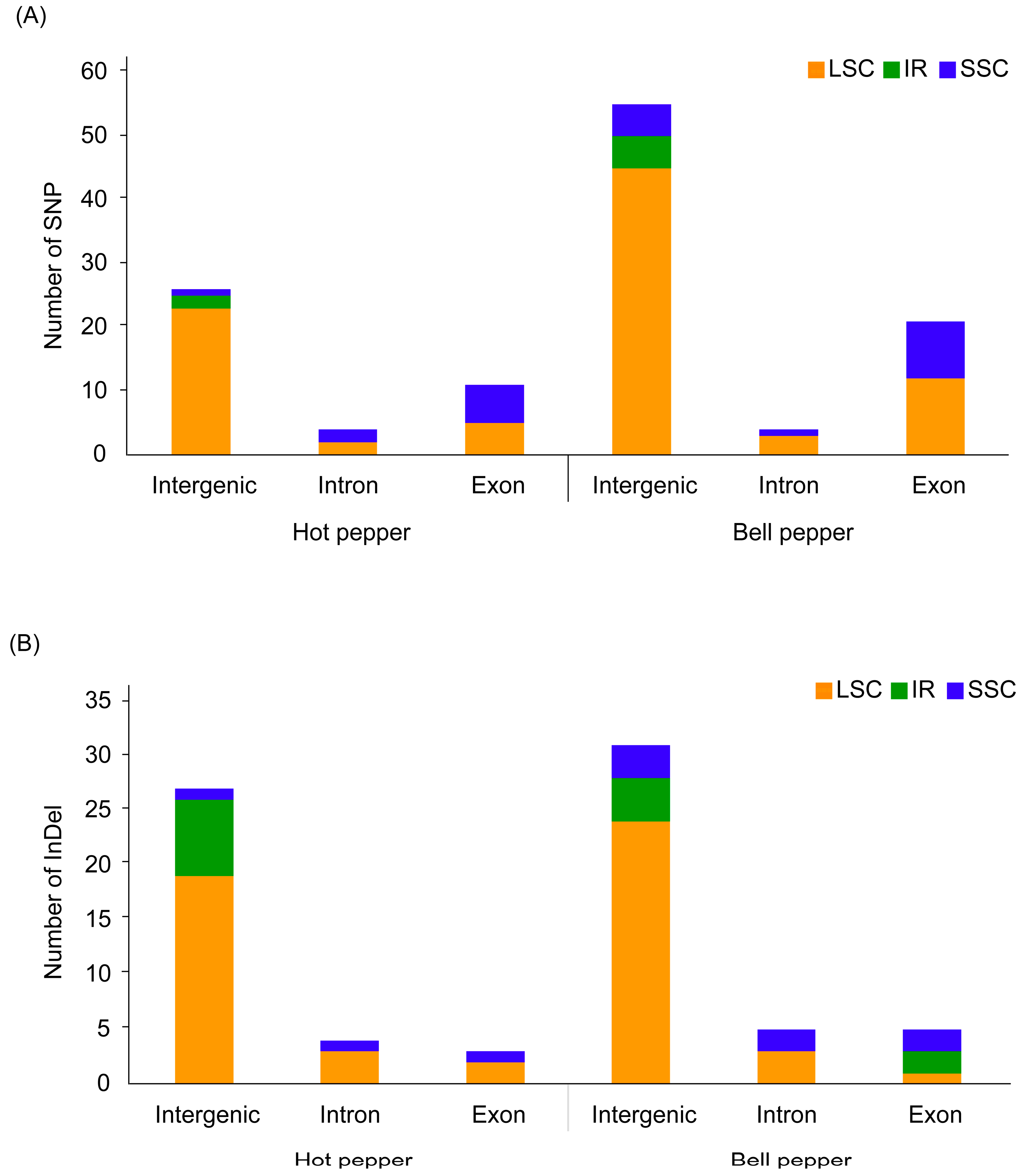
3.2. Plastome Sequence Variability
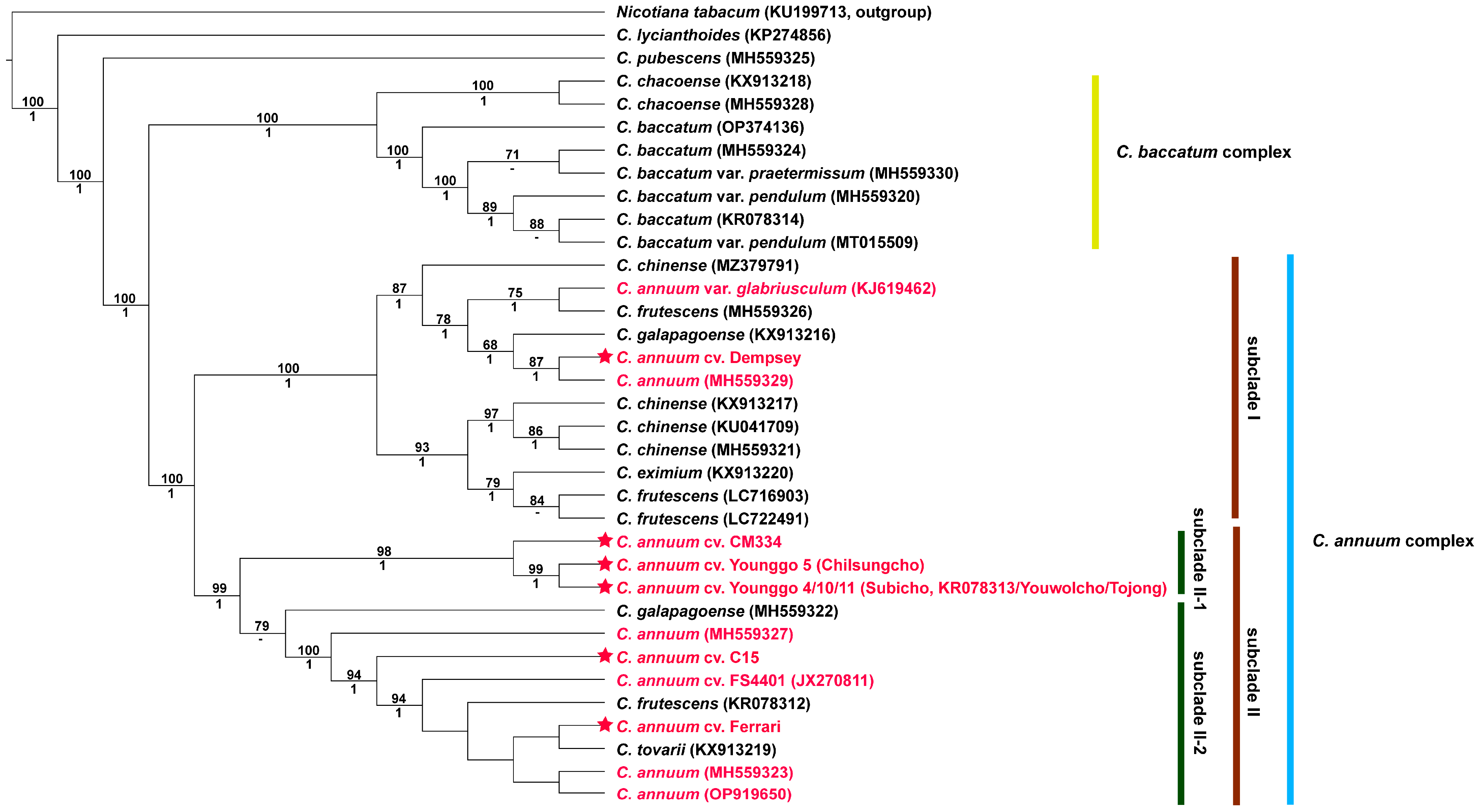
3.3. Phylogenetic Analysis
4. Discussion
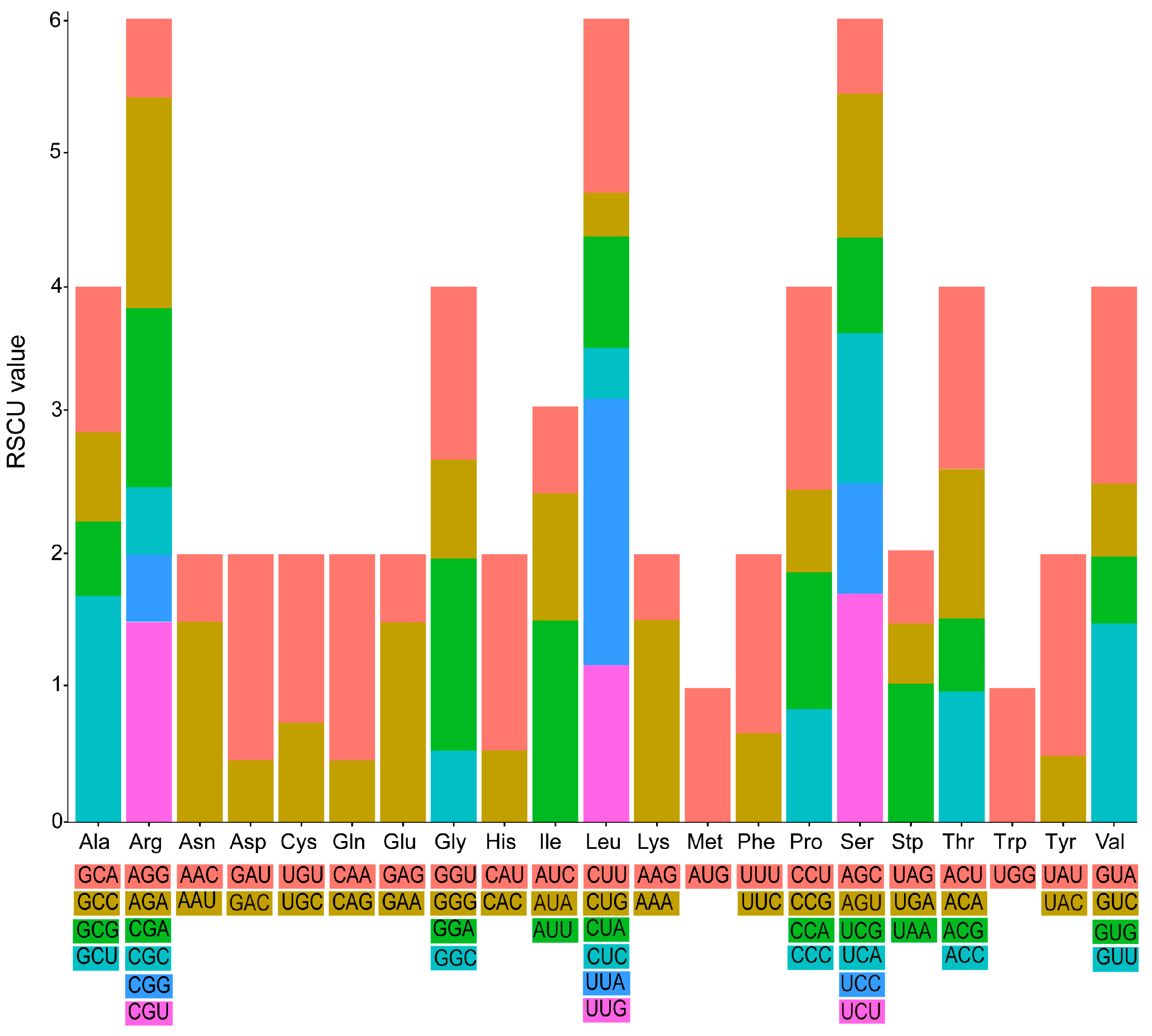
4.1. Plastome Feature
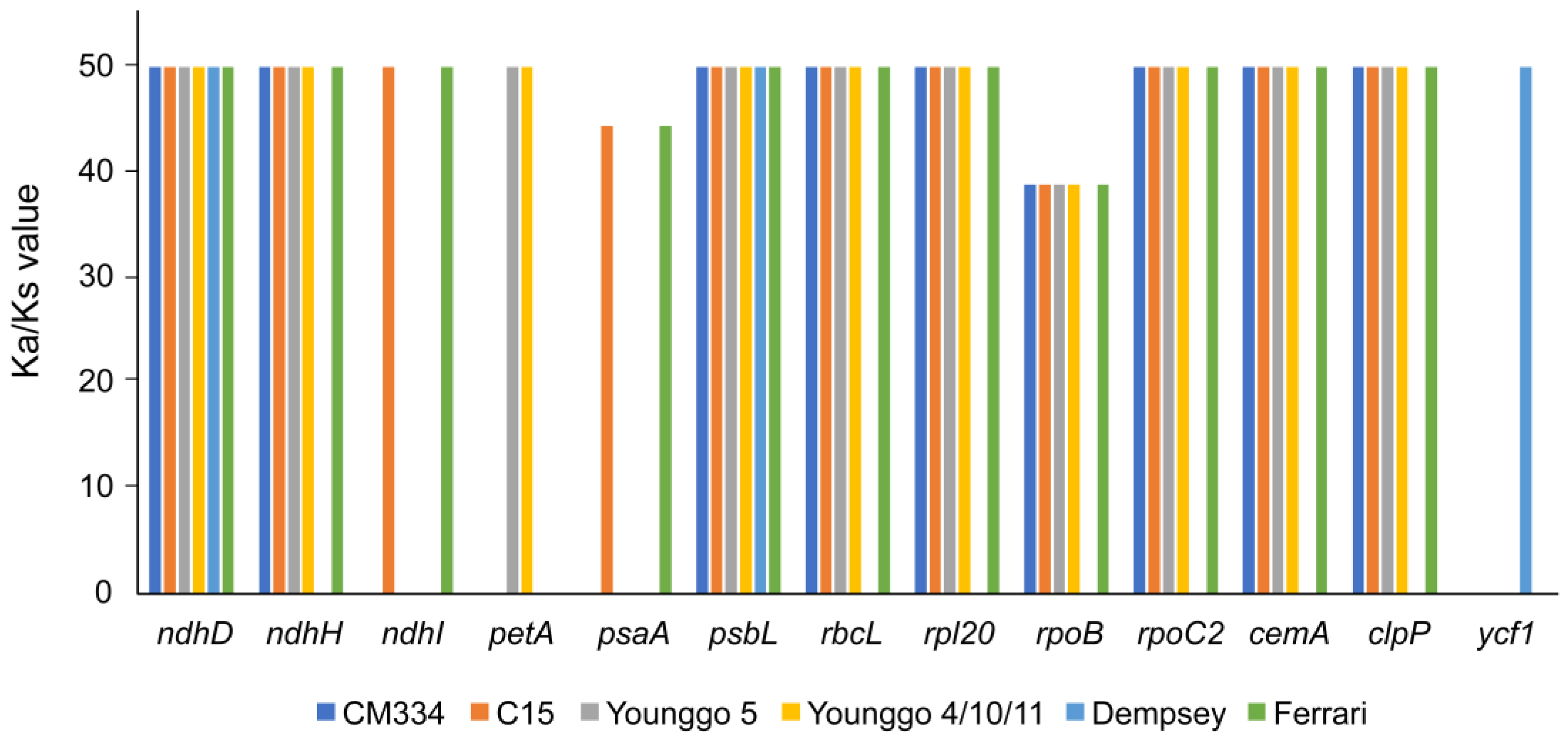
4.2. Sequence Divergence
4.3. Plastome Evolution
4.4. Phylogenetic Relationship
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carrizo García, C.; Barfuss, M.H.; Sehr, E.M.; Barboza, G.E.; Samuel, R.; Moscone, E.A.; Ehrendorfer, F. Phylogenetic relationships, diversification and expansion of chili peppers (Capsicum, Solanaceae). Ann. Bot. 2016, 118, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Barboza, G.E.; García, C.C.; Scaldaferro, M.; Bohs, L. An amazing new capsicum (Solanaceae) species from the andean-amazonian piedmont. PhytoKeys 2020, 167, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Pickersgill, B. Genetic resources and breeding of Capsicum spp. Euphytica 1997, 96, 129–133. [Google Scholar] [CrossRef]
- Shiragaki, K.; Yokoi, S.; Tezuka, T. Phylogenetic analysis and molecular diversity of Capsicum based on rDNA-ITS region. Horticulturae 2020, 6, 87. [Google Scholar] [CrossRef]
- Moscone, E.A.; Scaldaferro, M.A.; Grabiele, M.; Cecchini, N.M.; Sanchez García, Y.; Jarret, R.; Daviña, J.R.; Ducasse, D.A.; Barboza, G.E.; Ehrendorfer, F. The Evolution of Chili Peppers (Capsicum-Solanaceae): A Cytogenetic Perspective. Act. Hortic. 2007, 745, 137–169. [Google Scholar] [CrossRef]
- Walsh, B.M.; Hoot, S.B. Phylogenetic relationships of Capsicum (Solanaceae) using DNA sequences from two noncoding regions: The chloroplast atpB-rbcL spacer region and nuclear waxy introns. Int. J. Plant Sci. 2001, 162, 1409–1418. [Google Scholar] [CrossRef]
- Eshbaugh, W.H. Genetic and biochemical systematic studies of chili peppers (Capsicum-Solanaceae). Bull. Torrey Bot. Club 1975, 102, 396–403. [Google Scholar] [CrossRef]
- Moscone, E.A.; Baranyi, M.; Ebert, I.; Greilhuber, J.; Ehrendorfer, F.; Hunziker, A.T. Analysis of nuclear DNA content in Capsicum (Solanaceae) by flow cytometry and Feulgen densitometry. Ann. Bot. 2003, 92, 21–29. [Google Scholar] [CrossRef]
- Magdy, M.; Ou, L.; Yu, H.; Chen, R.; Zhou, Y.; Hassan, H.; Feng, B.; Taitano, N.; van der Knaap, E.; Zou, X.; et al. Pan-plastome approach empowers the assessment of genetic variation in cultivated Capsicum species. Hortic. Res. 2019, 6, 108. [Google Scholar] [CrossRef]
- Jung, M.; Shin, S.H.; Park, J.M.; Lee, S.N.; Lee, M.Y.; Ryu, K.H.; Paek, K.Y.; Harn, C.H. Detection of transgene in early developmental stage by GFP monitoring enhances the efficiency of genetic transformation of pepper. Plant Biotechnol. Rep. 2011, 5, 157–167. [Google Scholar] [CrossRef]
- Park, S.I.; Kim, H.B.; Jeon, H.J.; Kim, H. Agrobacterium-mediated Capsicum annuum gene editing in two cultivars, hot pepper CM334 and bell pepper dempsey. Int. J. Mol. Sci. 2021, 22, 3921. [Google Scholar] [CrossRef] [PubMed]
- Ravishankar, G.A.; Suresh, B.; Giridhar, P.; Rao, S.R.; Johnson, T.S. Biotechnological studies on Capsicum for metabolite production and plant improvement. In Capsicum: The Genus Capsicum, 1st ed.; De, A.K., Ed.; Taylor & Francis Inc.: New York, NY, USA, 2003; pp. 96–128. [Google Scholar]
- Kumar, A.; Kumar, S.; Anju, T.; Ramchiary, N. Genetic, epigenetic, and hormonal regulation of fruit development and ripening in Capsicum L. species. Annu. Plant Rev. 2018, 4, 295–356. [Google Scholar]
- Bagga, S.; Lucero, Y.; Apodaca, K.; Rajapakse, W.; Lujan, P.; Ortega, J.L.; Sengupta-Gopalan, C. Chile (Capsicum annuum) plants transformed with the RB gene from Solanum bulbocastanum are resistant to Phytophthora capsici. PLoS ONE 2019, 14, e0223213. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Jung, M.; Shin, S.H.; Lee, J.H.; Choi, S.H.; Her, N.H.; Lee, J.H.; Ryu, K.H.; Paek, K.Y.; Harn, C.H. Transgenic peppers that are highly tolerant to a new CMV pathotype. Plant Cell Rep. 2009, 28, 223–232. [Google Scholar] [CrossRef]
- Kim, H.; Choi, J.; Won, K.H. A stable DNA-free screening system for CRISPR/RNPs-mediated gene editing in hot and sweet cultivars of Capsicum annuum. BMC Plant Biol. 2020, 20, 449. [Google Scholar] [CrossRef]
- Christopher, T.; Rajam, M.V. Effect of genotype, explant and medium on in vitro regeneration of red pepper. Plant Cell Tissue Organ Cult. 1996, 46, 245–250. [Google Scholar] [CrossRef]
- Kim, J.Y.; Jung, M.; Kim, H.S.; Lee, Y.H.; Choi, S.H.; Lim, Y.P.; Min, B.W.; Yang, S.G.; Harn, C.H. A new selection system for pepper regeneration by mannose. J. Plant Biotechnol. 2002, 4, 129–134. [Google Scholar]
- Gyeongsangbuk-do Agricultural Research & Extension Services. Available online: https://www.gba.go.kr/index.do?menu_id=00000146 (accessed on 30 August 2023).
- Lee, M.J.; Kwon, T.R.; Pae, D.H.; Kim, D.U.; Choi, B.S. Characteristics of major local cultivars of red pepper (Capsicum annuum) grown in Youngyang region. J. Korean Soc. Hortic. Sci. 1999, 40, 158–162. [Google Scholar]
- Villa, T.C.C.; Maxted, N.; Scholten, M.; Ford-Lloyd, B. Defining and identifying crop landraces. Plant Genet. Resour. 2005, 3, 373–384. [Google Scholar] [CrossRef]
- Van Leur, J.A.G.; Ceccarelli, S.; Grando, S. Diversity for disease resistance in barley landraces from Syria and Jordan. Plant Breed. 1989, 103, 324–335. [Google Scholar] [CrossRef]
- Negri, V.; Maxted, N.; Veteläinen, M. European landrace conservation: An introduction. In European Landraces: On-Farm Conservation, Management and Use; Veteläinen, M., Negri, V., Maxted, N., Eds.; Bioversity International: Italy, Rome, 2009; pp. 1–22. [Google Scholar]
- Scarano, A.; Olivieri, F.; Gerardi, C.; Liso, M.; Chiesa, M.; Chieppa, M.; Frusciante, L.; Barone, A.; Santino, A.; Rigano, M.M. Selection of tomato landraces with high fruit yield and nutritional quality under elevated temperatures. J. Sci. Food Agric. 2020, 100, 2791–2799. [Google Scholar] [CrossRef] [PubMed]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: De novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 2017, 45, e18. [Google Scholar] [PubMed]
- Shi, L.; Chen, H.; Jiang, M.; Wang, L.; Wu, X.; Huang, L.; Liu, C. CPGAVAS2, an integrated plastome sequence annotator and analyzer. Nucleic Acids Res. 2019, 47, W65–W73. [Google Scholar] [CrossRef] [PubMed]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) version 1.3. 1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E. DnaSP v6: DNA sequence polymorphism analysis of large datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef]
- Brundo, M.; Malde, S.; Poliakov, A.; Do, C.B.; Couronne, O.; Dubchak, I.; Batzoglou, S. Global Alignment: Finding rearrangements during alignment. Bioinformatics 2003, 19, i54–i62. [Google Scholar]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z. KaKs_calculator 3.0: Calculating selective pressure on coding and non-coding sequences. Genom. Proteom. Bioinform. 2022, 20, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Lane, R.P.; McCarter, S.M.; Kuhn, C.W.; Deom, C.M. ‘Dempsey’, a virus-and bacterial spot-resistant bell pepper. Hortscience 1997, 32, 333–334. [Google Scholar] [CrossRef]
- Palmer, J.D. Comparative organization of chloroplast genomes. Annu. Rev. Genet. 1985, 19, 325–354. [Google Scholar] [CrossRef]
- Ravi, V.; Khurana, J.P.; Tyagi, A.K.; Khurana, P. An update on chloroplast genomes. Plant Syst. Evol. 2008, 271, 101–122. [Google Scholar] [CrossRef]
- Kuittinen, H.; Niittyvuopio, A.; Rinne, P.; Savolainen, O. Natural variation in Arabidopsis lyrata vernalization requirement conferred by a FRIGIDA indel polymorphism. Mol. Biol. Evol. 2008, 25, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Liao, L.; Zhou, H.; Gu, C.; Wang, L.; Han, Y. A small indel mutation in an anthocyanin transporter causes variegated colouration of peach flowers. J. Exp. Bot. 2015, 66, 7227–7239. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cheng, Y.; Xu, S.; Ma, H.; Han, J.; Zhang, Y. Tree peony variegated flowers show a small insertion in the F3′H gene of the acyanic flower parts. BMC Plant Biol. 2020, 20, 211. [Google Scholar] [CrossRef] [PubMed]
- Robles, P.; Quesada, V. Unveiling the functions of plastid ribosomal proteins in plant development and abiotic stress tolerance. Plant Physiol. Biochem. 2022, 189, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Pacheco-Olvera, A.; Hernández-Verdugo, S.; Rocha-Ramírez, V.; González-Rodríguez, A.; Oyama, K. Genetic diversity and structure of pepper (Capsicum annuum L.) from Northwestern Mexico analyzed by microsatellite markers. Crop Sci. 2012, 52, 231–241. [Google Scholar] [CrossRef]
- Williams, C.E.; Clair, D.A.S. Phenetic relationships and levels of variability detected by restriction fragment length polymorphism and random amplified polymorphic DNA analysis of cultivated and wild accessions of Lycopersicon esculentum. Genome 1993, 36, 619–630. [Google Scholar] [CrossRef]
- Wang, Y.; Rashid, M.A.R.; Li, X.; Yao, C.; Lu, L.; Bai, J.; Li, Y.; Xu, N.; Yao, Q.; Zhang, L.; et al. Collection and evaluation of genetic diversity and population structure of potato landraces and varieties in China. Front. Plant Sci. 2019, 10, 139. [Google Scholar] [CrossRef]
- Xiao, S.; Xu, P.; Deng, Y.; Dai, X.; Zhao, L.; Heider, B.; Zhang, A.; Zhou, Z.; Cao, Q. Comparative analysis of chloroplast genomes of cultivars and wild species of sweetpotato (Ipomoea batatas [L.] Lam). BMC Genom. 2021, 22, 262. [Google Scholar]
- Heidmann, I.; De Lange, B.; Lambalk, J.; Angenent, G.C.; Boutilier, K. Efficient sweet pepper transformation mediated by the BABY BOOM transcription factor. Plant Cell Rep. 2011, 30, 1107–1115. [Google Scholar] [CrossRef]
- Xu, C.; Dong, W.; Li, W.; Lu, Y.; Xie, X.; Jin, X.; Shi, J.; He, K.; Suo, Z. Comparative analysis of six Lagerstroemia complete chloroplast genomes. Front. Plant Sci. 2017, 8, 15. [Google Scholar] [CrossRef]
- Yun, S.; Kim, S.C. Comparative plastomes and phylogenetic analysis of seven Korean endemic Saussurea (Asteraceae). BMC Plant Biol. 2022, 22, 550. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, F.; Shahzadi, I.; Ahmed, I.; Waheed, M.T.; Mirza, B. Characterization of Withania somnifera chloroplast genome and its comparison with other selected species of Solanaceae. Genomics 2020, 112, 1522–1530. [Google Scholar] [CrossRef] [PubMed]
- Kalia, R.K.; Rai, M.K.; Kalia, S.; Singh, R.; Dhawan, A.K. Microsatellite markers: An overview of the recent progress in plants. Euphytica 2011, 177, 309–334. [Google Scholar] [CrossRef]
- Provan, J.; Powell, W.; Hollingsworth, P.M. Chloroplast microsatellites: New tools for studies in plant ecology and evolution. Trends Ecol. Evol. 2001, 16, 142–147. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, L.; Wang, W.; Zhang, Z.; Du, H.; Qu, Z.; Li, X.Q.; Xiang, H. Differences in codon usage bias between photosynthesis-related genes and genetic system-related genes of chloroplast genomes in cultivated and wild solanum species. Int. J. Mol. Sci. 2018, 19, 3142. [Google Scholar] [CrossRef]
- Cho, K.S.; Cheon, K.S.; Hong, S.Y.; Cho, J.H.; Im, J.S.; Mekapogu, M.; Yu, Y.S.; Park, T.H. Complete chloroplast genome sequences of Solanum commersonii and its application to chloroplast genotype in somatic hybrids with Solanum tuberosum. Plant Cell Rep. 2016, 35, 2113–2123. [Google Scholar] [CrossRef]
- Mehmood, F.; Ubaid, Z.; Shahzadi, I.; Ahmed, I.; Waheed, M.T.; Poczai, P.; Mirza, B. Plastid genomics of Nicotiana (Solanaceae): Insights into molecular evolution, positive selection and the origin of the maternal genome of Aztec tobacco (Nicotiana rustica). PeerJ 2020, 8, e9552. [Google Scholar] [CrossRef]
- Saldaña, C.L.; Chávez-Galarza, J.C.; De la Cruz, G.; Jhoncon, J.H.; Guerrero-Abad, J.C.; Vásquez, H.V.; Maicelo, J.L.; Arbizu, C.I. Revealing the complete chloroplast genome of an Andean horticultural crop, sweet cucumber (Solanum muricatum), and its comparison with other Solanaceae species. Data 2022, 7, 123. [Google Scholar] [CrossRef]
- Peng, L.; Yamamoto, H.; Shikanai, T. Structure and biogenesis of the chloroplast NAD (P) H dehydrogenase complex. Biochim. Biophys. Acta (BBA)-Bioenerg. 2011, 1807, 945–953. [Google Scholar] [CrossRef]
- Kapralov, M.V.; Filatov, D.A. Widespread positive selection in the photosynthetic Rubisco enzyme. BMC Evol. Biol. 2007, 7, 73. [Google Scholar] [CrossRef]
- Bisson, G.P.; Mehaffy, C.; Broeckling, C.; Prenni, J.; Rifat, D.; Lun, D.S.; Burgos, M.; Weissman, D.; Karakousis, P.C.; Dobos, K. Upregulation of the phthiocerol dimycocerosate biosynthetic pathway by rifampin-resistant, rpoB mutant Mycobacterium tuberculosis. J. Bacteriol. 2012, 194, 6441–6452. [Google Scholar] [CrossRef] [PubMed]
- Pickersgill, B. Relationships between weedy and cultivated forms in some species of chili peppers (genus Capsicum). Evolution 1971, 25, 683–691. [Google Scholar] [PubMed]
- McLeod, M.J.; Guttman, S.I.; Eshbaugh, W.H.; Rayle, R.E. An electrophoretic study of evolution in Capsicum (Solanaceae). Evolution 1983, 37, 562–574. [Google Scholar] [CrossRef] [PubMed]
- Paran, I.; Aftergoot, E.; Shifriss, C. Variation in Capsicum annuum revealed by RAPD and AFLP markers. Euphytica 1998, 99, 167–173. [Google Scholar] [CrossRef]
- Ince, A.G.; Karaca, M.; Onus, A.N. Genetic relationships within and between Capsicum species. Biochem. Genet. 2010, 48, 83–95. [Google Scholar] [CrossRef]
- Geleta, L.F.; Labuschagne, M.T.; Viljoen, C.D. Genetic variability in pepper (Capsicum annuum L.) estimated by morphological data and amplified fragment length polymorphism markers. Biodivers. Conserv. 2005, 14, 2361–2375. [Google Scholar] [CrossRef]
- Baral, J.; Bosland, P.W. Genetic diversity of a Capsicum germplasm collection from Nepal as determined by randomly amplified polymorphic DNA markers. J. Am. Soc. Hortic. Sci. 2002, 127, 318–324. [Google Scholar] [CrossRef]
- Nicolaï, M.; Cantet, M.; Lefebvre, V.; Sage-Palloix, A.M.; Palloix, A. Genotyping a large collection of pepper (Capsicum spp.) with SSR loci brings new evidence for the wild origin of cultivated C. annuum and the structuring of genetic diversity by human selection of cultivar types. Genet. Resour. Crop Evol. 2013, 60, 2375–2390. [Google Scholar] [CrossRef]
- Rai, V.P.; Kumar, R.; Kumar, S.; Rai, A.; Kumar, S.; Singh, M.; Singh, S.P.; Rai, A.B.; Paliwal, R. Genetic diversity in Capsicum germplasm based on microsatellite and random amplified microsatellite polymorphism markers. Physiol. Mol. Biol. Plants 2013, 19, 575–586. [Google Scholar] [CrossRef]
- Jeong, H.J.; Jo, Y.D.; Park, S.W.; Kang, B.C. Identification of Capsicum species using SNP markers based on high resolution melting analysis. Genome 2010, 53, 1029–1040. [Google Scholar] [CrossRef]
- Tong, N.; Bosland, P.W. Capsicum tovarii, a new member of the Capsicum baccatum complex. Euphytica 1999, 109, 71–77. [Google Scholar] [CrossRef]
- KIPRIS. Transgenic Pepper with Enhanced Tolerance to PepMoV and Production Method Thereof. Available online: http://www.kipris.or.kr/khome/main.jsp (accessed on 11 September 2023).
- Jung, M. Development of Genetically Modified Peppers Tolerant to Pepper Mottle Virus through the Establishment of Highly Efficient Transformation System. Ph.D. Thesis, Chungbuk National University, Cheongju, Republic of Korea, 2011. [Google Scholar]
- Brickell, C.D.; Alexander, C.; Cubey, J.J.; David, J.C.; Hoffman, M.H.A.; Lslie, A.C.; Malécot, V.; Jin, X. International Code of Nomenclature for Cultivated Plants, 9th ed.; ISHS: Leuven, Belgium, 2016; pp. 6–10. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | C15 | CM334 | Younggo 5 | Younggo 4/10/11 | Dempsey | Ferrari |
|---|---|---|---|---|---|---|
| Total size (bp) | 156,821 | 156,881 | 156,922 | 156,878 | 156,895 | 156,826 |
| LSC (bp) | 87,384 | 87,256 | 87,391 | 87,347 | 87,395 | 87,380 |
| SSC (bp) | 17,853 | 17,939 | 17,929 | 17,929 | 17,920 | 17,862 |
| IR (bp) | 25,792 | 25,843 | 25,801 | 25,801 | 25,790 | 25,792 |
| Total GC content (%) | 37.7 | 37.7 | 37.7 | 37.7 | 37.7 | 37.7 |
| LSC (%) | 35.7 | 35.8 | 35.7 | 35.7 | 35.7 | 35.7 |
| SSC (%) | 32.0 | 32.0 | 32.0 | 32.0 | 32.0 | 32.0 |
| IR (%) | 43.1 | 43.0 | 43.1 | 43.1 | 43.1 | 43.1 |
| Number of total genes | 113 | 113 | 113 | 113 | 113 | 113 |
| Number of protein-coding genes | 79 | 79 | 79 | 79 | 79 | 79 |
| Number of tRNA genes | 30 | 30 | 30 | 30 | 30 | 30 |
| Number of rRNA genes | 4 | 4 | 4 | 4 | 4 | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yun, S.; Kim, H. Insight into the Phylogenetic Relationships and Evolutionary History of Pepper Cultivars (Capsicum annuum L.) through Comparative Analyses of Plastomes. Horticulturae 2023, 9, 1092. https://doi.org/10.3390/horticulturae9101092
Yun S, Kim H. Insight into the Phylogenetic Relationships and Evolutionary History of Pepper Cultivars (Capsicum annuum L.) through Comparative Analyses of Plastomes. Horticulturae. 2023; 9(10):1092. https://doi.org/10.3390/horticulturae9101092
Chicago/Turabian StyleYun, Seona, and Hyeran Kim. 2023. "Insight into the Phylogenetic Relationships and Evolutionary History of Pepper Cultivars (Capsicum annuum L.) through Comparative Analyses of Plastomes" Horticulturae 9, no. 10: 1092. https://doi.org/10.3390/horticulturae9101092