
Identification of the Transcription Factors RAP2-13 Activating the Expression of CsBAK1 in Citrus Defence Response to Xanthomonas citri subsp. citri

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Bacterial Pathogen Preparation
2.3. RNA Isolation and Quantitative RT-PCR
2.4. Isolation and Bioinformatic Analysis of the CsBAK1 Gene in Citrus
2.5. Transient Expression of CsBAK1 in’Bingtang’ Sweet Orange Leaves
2.6. Construction of BAK1 Promoter Vectors and ‘Bingtang’ Sweet Orange Leaf Transformation
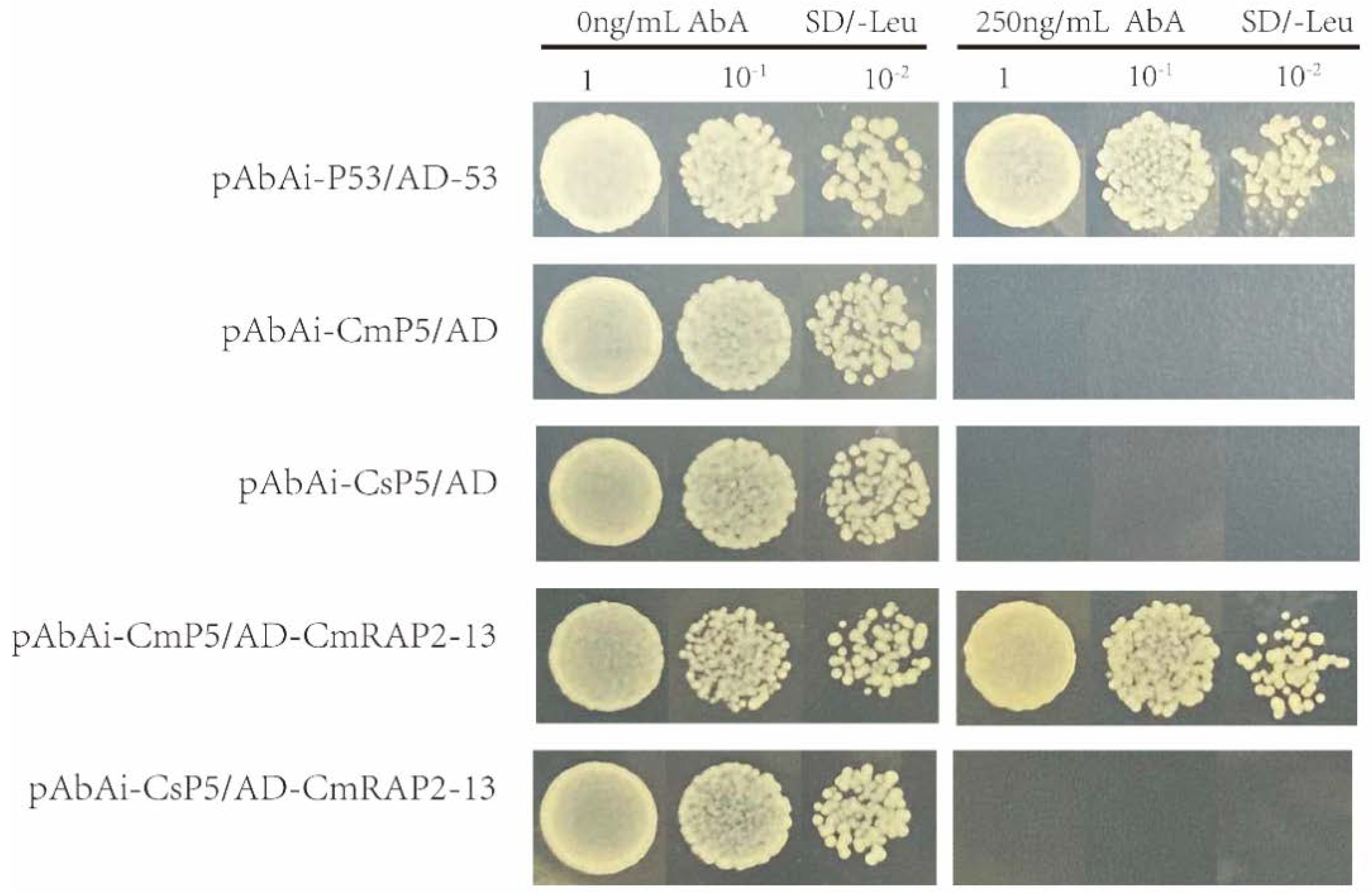
2.7. Yeast One-Hybrid (Y1H) Screening
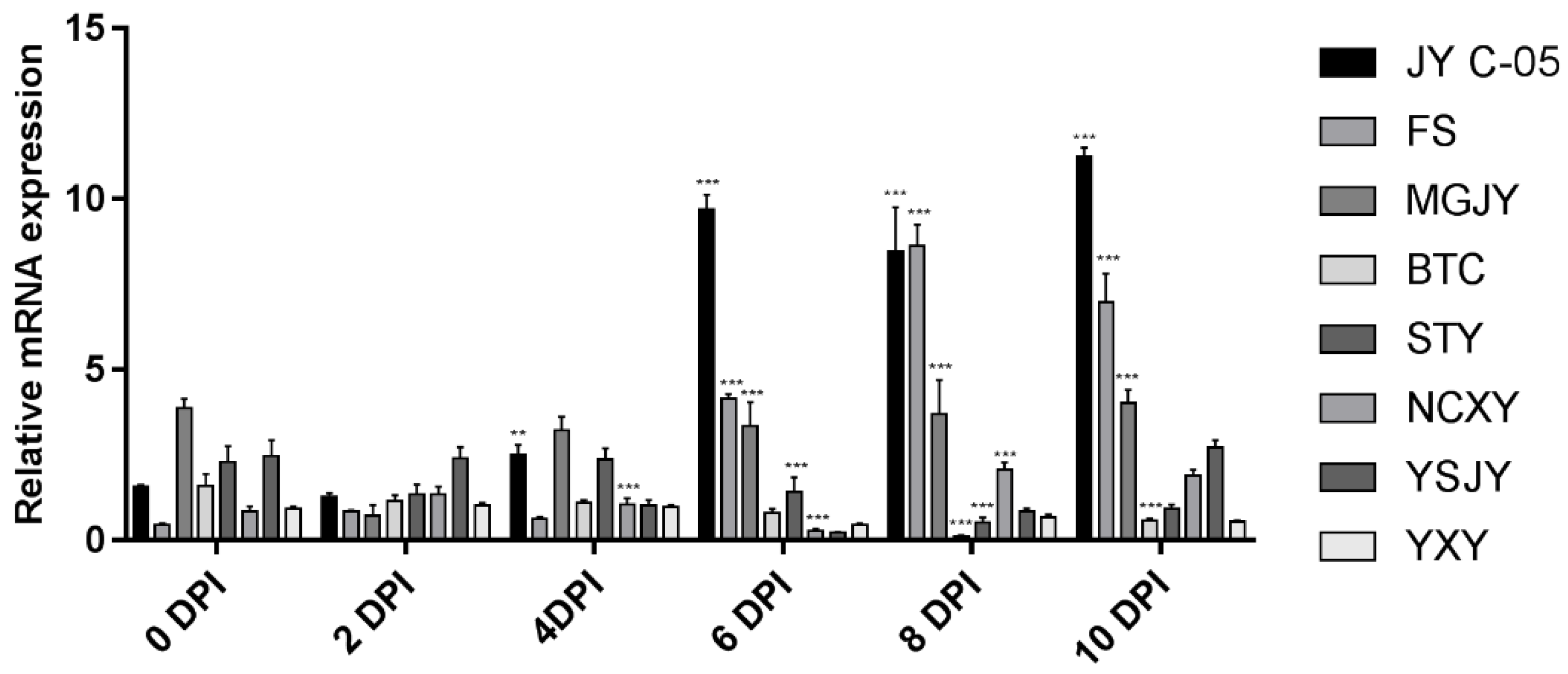
2.8. Expression Profiling of Transcription Factors
2.9. Data Processing
3. Results
3.1. Canker Symptom Development on Leaves of Susceptible and Resistant Sweet Orange Cultivars
3.2. Analysis of CsBAK1 Expression Levels in Citrus Leaves
3.3. Isolation and Phylogenetic Analysis of the BAK1 Gene from the Citron C-05 Cultivar
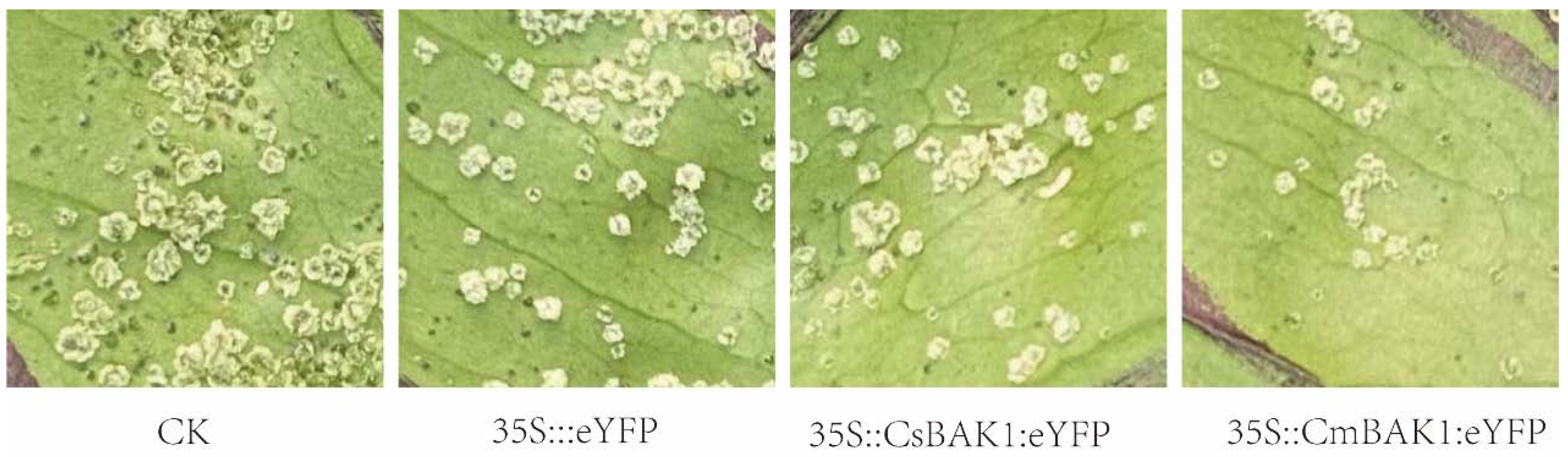
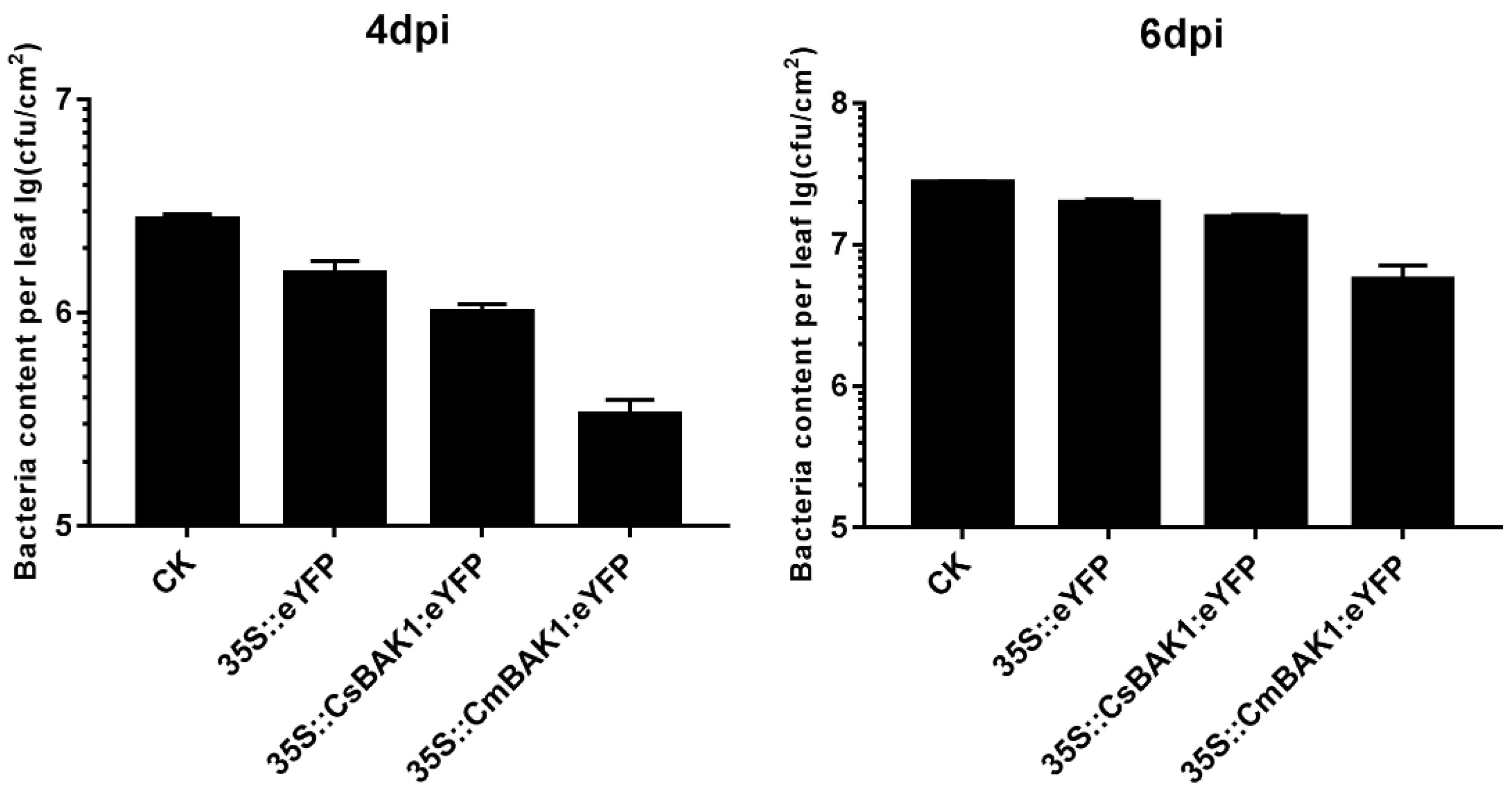
3.4. Transient Expression of CsBAK1 in ‘Bingtang’ Sweet Orange Leaves
3.5. Analysis of the BAK1 Promoter Region
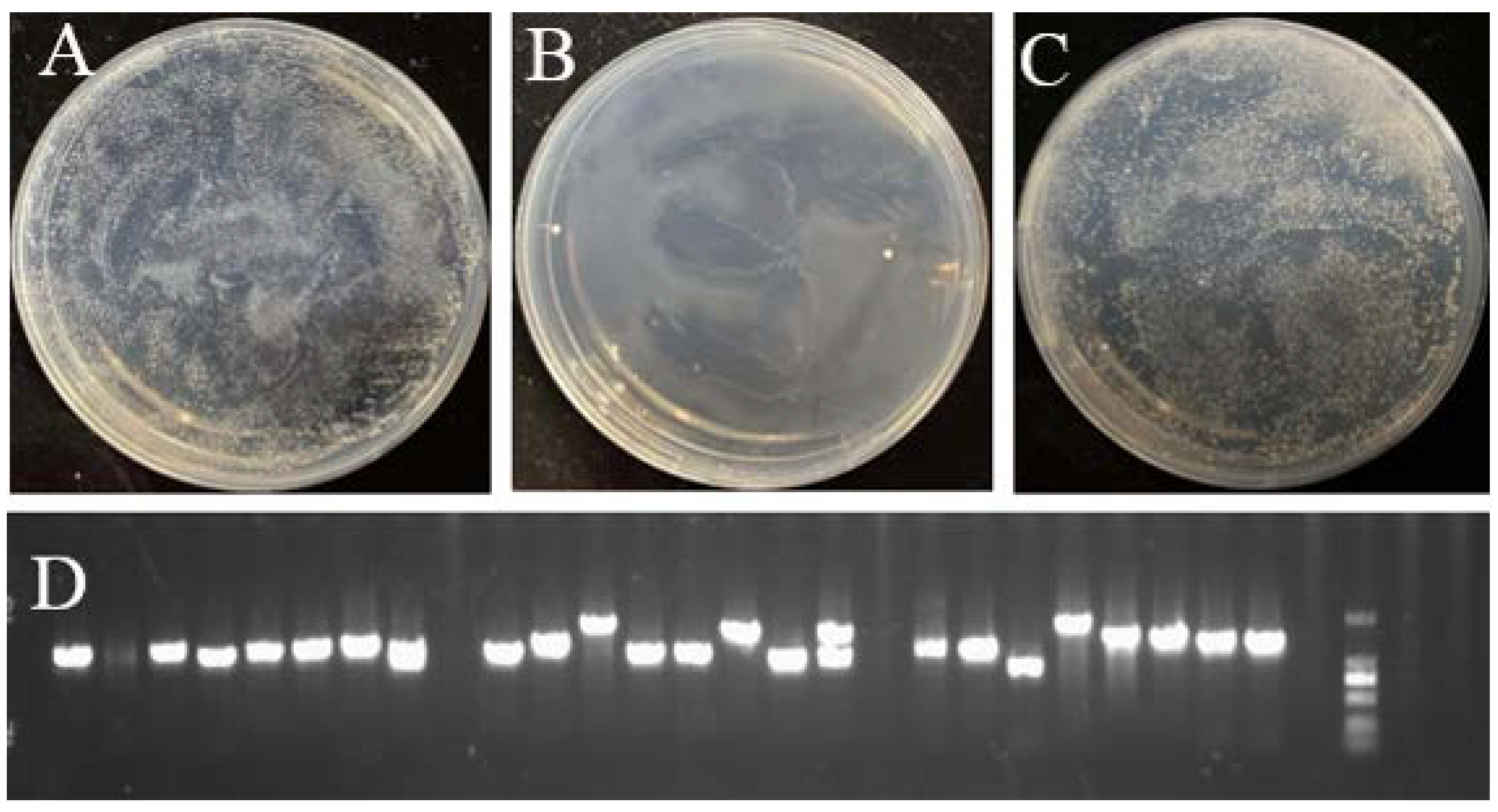
3.6. Identification of BAK1 Promoter Binding Proteins
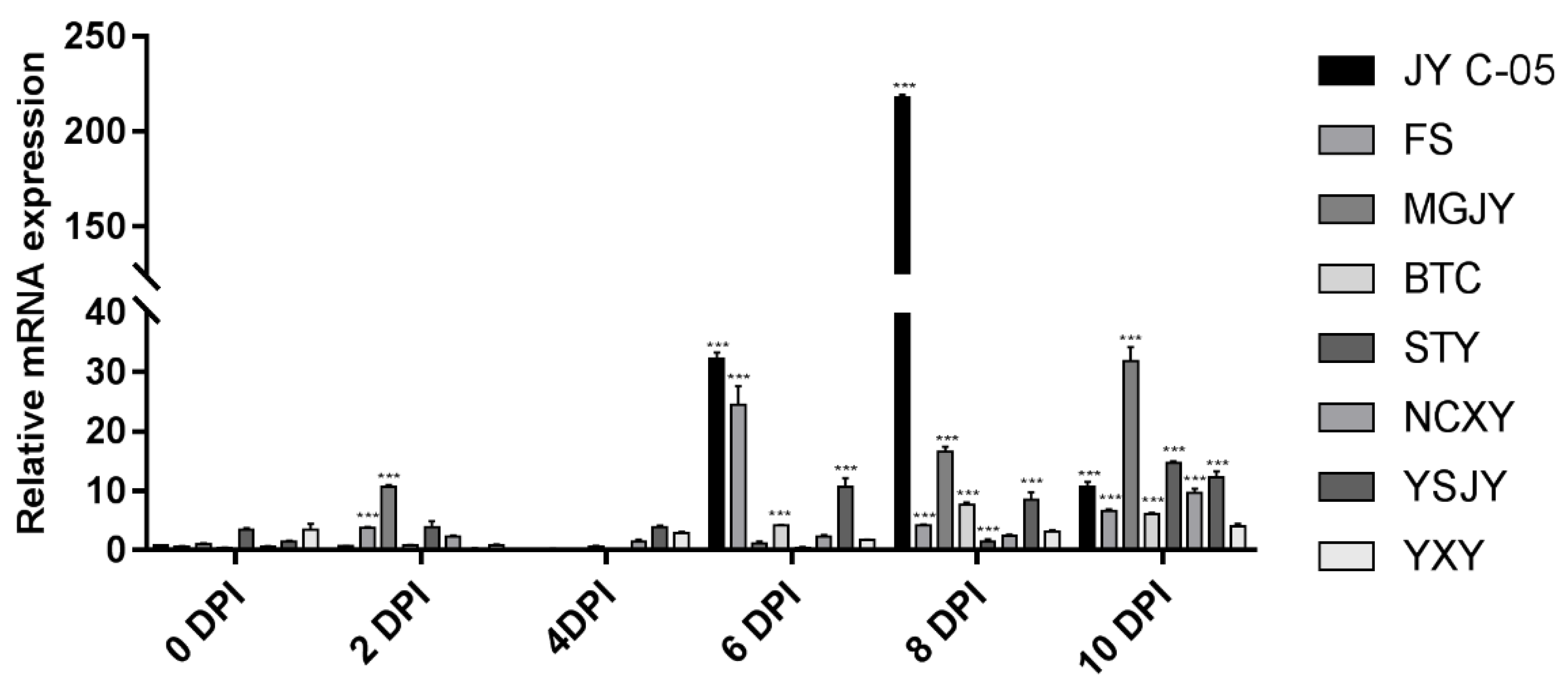
3.7. RAP2-13 Gene Expression in Citrus Leaves
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Rukshana, N.; Popy, B.; Medhi, K.K. Citrus Canker: Developments down the lane. Ann. Plant Soil Res. 2020, 22, 396–404. [Google Scholar]
- Gottwald, T.R.; Hughes, G.; Graham, J.H.; Sun, X.; Riley, T. The citrus canker epidemic in Florida: The scientific basis of regulatory eradication policy for an invasive species. Phytopathology 2001, 91, 30–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lúcia, B.C.; Caio FC, Z.; Luiz, L.S.; Carlos, R.P. Hexyl gallate for the control of citrus canker caused by Xanthomonas citri subsp citri. Microbiol. Open 2020, 9, e1104. [Google Scholar]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 23–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, A.; Nicholas, P.; Nicholas, N.; Xiao, L.J.; John, M.H.; Xing, H.Z. Genomic Analysis for Citrus Disease Detection. OBM Genet. 2021, 5, 124. [Google Scholar] [CrossRef]
- Shi, Q.; Febres, V.J.; Jones, J.B.; Moore, G.A. A survey of FLS2 genes from multiple citrus species identifies candidates for enhancing disease resistance to Xanthomona scitri ssp. citri. Hortic. Res. 2016, 3, 16022. [Google Scholar] [CrossRef] [Green Version]
- Ding-li, L.; Xiao, X.; Guo, W. Production of transgenic ‘anliucheng’ sweet orange (Citrus sinensis Osbeck) with Xa21 gene for potential canker resistance. J. Integr. Agric. 2014, 13, 2370–2377. [Google Scholar]
- Domínguez-Ferreras, A.; Kiss-Papp, M.; Jehle, A.K.; Felix, G.; Chinchilla, D. An Overdose of the Arabidopsis Coreceptor Brassinosteroid Insensitive1-Associated Receptor Kinase1 or Its Ectodomain Causes Autoimmunity in a Suppressor of Bir1-1-Dependent Manner. Plant Physiol. 2015, 168, 1106–1121. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.A.; Prochnik, S.; Jenkins, J.; Salse, J.; Hellsten, U.; Murat, F.; Perrier, X.; Ruiz, M.; Scalabrin, S.; Terol, J. Sequencing of diverse mandarin, pummelo and orange genomes reveals complex history of admixture during citrus domestication. Nat. Biotechnol. 2014, 32, 656–662. [Google Scholar] [CrossRef]
- Hecht, V.; Vielle-Calzada, J.P.; Hartog, M.V.; Schmidt, E.D.; Boutilier, K.; Grossniklaus, U.; de Vries, S.C. The Arabidopsis somatic embryogenesis receptor kinase 1 gene is expressed in developing ovules and embryos and enhances embryogenic competence in culture. Plant Physiol. 2001, 127, 803–816. [Google Scholar] [CrossRef]
- Chinchilla, D.; Zipfel, C.; Robatzek, S.; Kemmerling, B.; Nurnberger, T.; Jones, J.D.; Felix, G.; Boller, T. A flagellin-induced complex of the receptor FLS2 and BAK1 initiates plant defence. Nature 2007, 448, 497–500. [Google Scholar] [CrossRef] [Green Version]
- Heese, A.; Hann, D.R.; Gimenez-Ibanez, S.; Jones, A.M.; He, K.; Li, J.; Schroeder, J.I.; Peck, S.C.; Rathjen, J.P. The receptor-like kinase SERK3/BAK1 is a central regulator of innate immunity in plants. Proc. Natl. Acad. Sci. USA 2007, 104, 12217–12222. [Google Scholar] [CrossRef] [Green Version]
- Kemmerling, B.; Nürnberger, T. Brassinosteroid-independent functions of the BRI1-associated kinase BAK1/SERK3. Plant Signal Behav. 2008, 3, 116–118. [Google Scholar] [CrossRef] [PubMed]
- Vert, G. Plant signaling: Brassinosteroids, immunity and effectors are BAK1. Curr. Biol. 2008, 18, R963–R965. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.N.; Hu, C.H.; Xu, L.; Zhang, J.Y.; Yang, L.; Li, D.Z.; Li, N.; Liu, L.P. Strategies for citrus to acquire resistance to canker disease. Proc. Int. Soc. Citric. 2010, 2, 1021–1027. [Google Scholar]
- Fu, H.Y.; Zhao, M.M.; Xu, J.; Tan, L.M.; Han, J.; Li, D.Z.; Wang, M.J.; Xiao, S.Y.; Ma, X.F.; Deng, Z.N. Citron C-05 inhibits both the penetration and colonization of Xanthomonas citri subsp. citri to achieve resistance to citrus canker disease. Hortic. Res. 2020, 7, 58. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhang Jl Ji, H.G.; Sossod, D.; Lie, T.; Frommerd, W.B.; Bing, Y.; Frank FWhiteb Wang, N.; Jones, J.B. Lateral organ boundaries 1 is a disease susceptibility gene for citrus bacterial canker disease. Proc. Natl. Acad. Sci. USA 2014, 114, E521–E529. [Google Scholar] [CrossRef] [Green Version]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef]
- Dodds, P.N.; Rathjen, J.P. Plant immunity: Towards an integrated view of plant-pathogen interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef]
- Dangl, J.L.; Horvath, D.M.; Staskawicz, B.J. Pivoting the plant immune system from dissection to deployment. Science 2013, 341, 746–751. [Google Scholar] [CrossRef] [Green Version]
- Bohm, H.; Albert, I.; Reinhard, A. Nurnberger, Immune receptor complexes at the plant cell surface. Curr. Opin. Plant Biol. 2014, 20, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Couto, D.; Zipfel, C. Regulation of pattern recognition receptor signalling in plants. Nat. Rev. Immunol. 2006, 16, 537–555. [Google Scholar] [CrossRef] [PubMed]
- Saijo, Y.; Loo, E.P.; Yasuda, S. Pattern recognition receptors and signaling in plant-microbe interactions. Plant J. 2018, 93, 592–613. [Google Scholar] [CrossRef] [PubMed]
- Roux, M.; Schwessinger, B.; Albrecht, C.; Chinchilla, D.; Jones, A.; Holton, N.; Malinovsky, F.G.; Tor, M.; de Vries, S.; Zipfel, C. The A rabidopsis leucine-rich repeat receptor-like kinases BAK1/SERK3 and BKK1/SERK4 are required for innate im m unity to hem ibiotrophic and biotrophic pathogens. Plant Cell 2011, 23, 2440–2455. [Google Scholar] [CrossRef]
- Sun, Y.; Han, Z.; Tang, J.; Hu, Z.; Chai, C.; Zhou, B.; Chai, J. Structure reveals that BAK1 as a co-receptor recognizes the BRI1-bound brassinolide. Cell Res. 2013, 23, 1326–1329. [Google Scholar] [CrossRef]
- Schulze, B.; Mentzel, T.; Jehle, A.K.; Mueller, K.; Beeler, S.; Boller, T.; Felix, G.; Chinchilla, D. Rapid heteromerization and phosphorylation of ligand-activated plant transmembrane receptors and their associated kinase BAK1. J. Biol. Chem. 2010, 285, 9444–9451. [Google Scholar] [CrossRef] [Green Version]
- Schwessinger, B.; Bahar, O.; Thomas, N.; Holton, N.; Nekrasov, V.; Ruan, D.; Canlas, P.E.; Daudi, A.; Petzold, C.J.; Singan, V.R.; et al. Transgenic expression of the dicotyledonous pattern recognition receptor EFR in rice leads to ligand dependent activation of defense responses. PLoS Pathog. 2015, 11, e1004809. [Google Scholar]
- Petutschnig, E.K.; Jones, A.M.; Serazetdinova, L.; Lipka, U.; Lipka, V. The lysin motif receptor-like kinase (LysM-RLK) CERK1 is a major chitin-binding protein in Arabidopsis thaliana and subject to chitin-induced phosphorylation. J. Biol. Chem. 2010, 285, 28902–28911. [Google Scholar] [CrossRef] [Green Version]
- Mazarei, M.; Teplova, I.; Hajimorad, M.R.; Stewart, C.N. Pathogen phytosensing: Plants to report plant pathogens. Sensors 2008, 8, 2628–2641. [Google Scholar] [CrossRef] [Green Version]
- Smirnova, O.G.; Ibragimova, S.S.; Kochetov, A.V. Simple data base to select promoters for plant transgenesis. Transgenic Res. 2012, 21, 429–437. [Google Scholar] [CrossRef]
- Zhou, J.M.; Zhang, Y. Plant immunity: Danger perception and signaling. Cell 2020, 181, 978–989. [Google Scholar] [CrossRef] [PubMed]
- Jakoby, M.; Weisshaar, M.J.B.; Dröge-Laser, W.; Vicente-Carbajosa, J.; Tiedemann, J.; Kroj, T.; Parcy, F. bZIP transcription factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–111. [Google Scholar] [CrossRef]
- Song, N.; Ma, L.; Wang, W.; Sun, H.; Wang, L.; Baldwin, I.T.; Wu, J. An ERF2-like transcription factor regulates production of the defense sesquiterpene capsidiol upon Alternaria alternata in-fection. J. Exp. Bot. 2019, 70, 5895–5908. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| BAK1 | ACCAGAATACGGGCAGACCT | AAGGTTCCCGTCAACTCGTTA |
| EF1-α | GTAACCAAGTCTGCTGCCAAG | GACCCAAACACCCAACACATT |
| Gene | Primer (5′-3′) |
|---|---|
| gBAK1-F | GGACGAGCTCGGTACCATGTCGGACGATGAAAACGA |
| gBAK1-R | GCCCTTGCTCACCATATCACAGTGAGGCAGTATCTGATT |
| Gene | Primer (5′-3′) |
|---|---|
| CmP1-F | AGGATCCCCAATACTTTGTTCAAAGCTGGGTCAAACC |
| CmP1-R | TGGATCCCCAATACTGTTCATCCAACTAATCTGATCTTCT |
| CmP2-F | AGGATCCCCATACTGACATCATCAATTCATAGTTCAGGT |
| CmP3-F | AGGATCCCCAATACTGTAATTTCCAACGTCGCACTTT |
| CmP4-F | AGGATCCCCAATACTCGTGAACACTAAATAACAACATTT |
| CmP-R | TGGATCCCCAATACTTATTGAAATACAAGTAATGCAGGT |
| Gene | Primer (5′-3′) |
|---|---|
| BAK1-CmP4-F | gaaaagcttgaattcgTAAGTATTATTGTATTGTTCAAAGCTGG |
| BAK1-CmP4-R | atacagagcacatgccATATTGAAATACAAGTAATTTGCAGGT |
| Gene | Primer (5′-3′) |
|---|---|
| RAP2-13-AD-F | gtaccagattacgctcaATGGCGGCTACAATGGATTT |
| RAP2-13-AD-R | atgcccacccgggtggTTAAGATAATATTGAAGCCCAATCAAT |
| Gene | Primer (5′-3′) |
|---|---|
| RAP2-13-F | CCGTTTGGTGGTGAACTTATGG |
| RAP2-13-R | GAGGAGGTGGGTAAGACTGGTAAT |
| Cisacting Element | Position (bp) | |||
|---|---|---|---|---|
| Susceptible Citrus Germplasm | Resistant Citrus | |||
| BTC | YSXY | MGXY | JY-C05 | |
| CCAAT-box | −207 | |||
| TCT-motif | +1074, +1452 | +636, +1004, +1452 | ||
| WUN-motif | +868, +1676 | +903, +1672 | −106, +1671 | +883, +1671 |
| MYB | +946 | +302 | ||
| ABRE | +1570 | -- | -- | -- |
| G-box | +784, +1569 | +819, +1105 | −1573 | +799, +1086 |
| Box 4 | +59, +188, +192, −1374, −1707 | +93, +222, +226, −1379, −1703, −1709, −1713 | +68, +270, +511, +640, +644, −1270, −1708, −1712 | +73, +202, +206, −1378, −1708, −1702, −1712 |
| DRE | −1511 | −1510 | −1510 | |
| ARE | +52, +1391, +1068, −1806, −1921 | +470, +1390,+1806, −1921 | +32, +1364, +1390, +1608, −1806, −1921 | |
| P-box | +97 | +131 | +549 | +111 |
| W box | −14, −198, +742, −1638 | −48, −232, +777, −1633 | −446, −650, −1195, −1632 | −28, −212, +757, −1632 |
| TCA-element | +709, +769 | +1162 | ||
| AT-rich element | +1862 | +1862 | +1862 | |
| MYC | +797 | +832 | + | +812 |
| ERE | +203, +1210, −1279 | +237, −1226, −1309 | +655 | +217, −1207, −1290 |
| STRE | +250, +1640 | +668, +1639 | +230, +1639 | |
| TC-rich repeats | −35, −589 | −624 | −487, −1042 | −49, −1197 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Q.; Zhao, M.; Li, Y.; Li, D.; Ma, X.; Deng, Z. Identification of the Transcription Factors RAP2-13 Activating the Expression of CsBAK1 in Citrus Defence Response to Xanthomonas citri subsp. citri. Horticulturae 2022, 8, 1012. https://doi.org/10.3390/horticulturae8111012
Wu Q, Zhao M, Li Y, Li D, Ma X, Deng Z. Identification of the Transcription Factors RAP2-13 Activating the Expression of CsBAK1 in Citrus Defence Response to Xanthomonas citri subsp. citri. Horticulturae. 2022; 8(11):1012. https://doi.org/10.3390/horticulturae8111012
Chicago/Turabian StyleWu, Qi, Mingming Zhao, Yi Li, Dazhi Li, Xianfeng Ma, and Ziniu Deng. 2022. "Identification of the Transcription Factors RAP2-13 Activating the Expression of CsBAK1 in Citrus Defence Response to Xanthomonas citri subsp. citri" Horticulturae 8, no. 11: 1012. https://doi.org/10.3390/horticulturae8111012