
Identification of Phenological Growth Stages of Four Morus Species Based on the Extended BBCH-Scale and Its Application in Fruit Development with Morphological Profiles and Color Characteristics


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
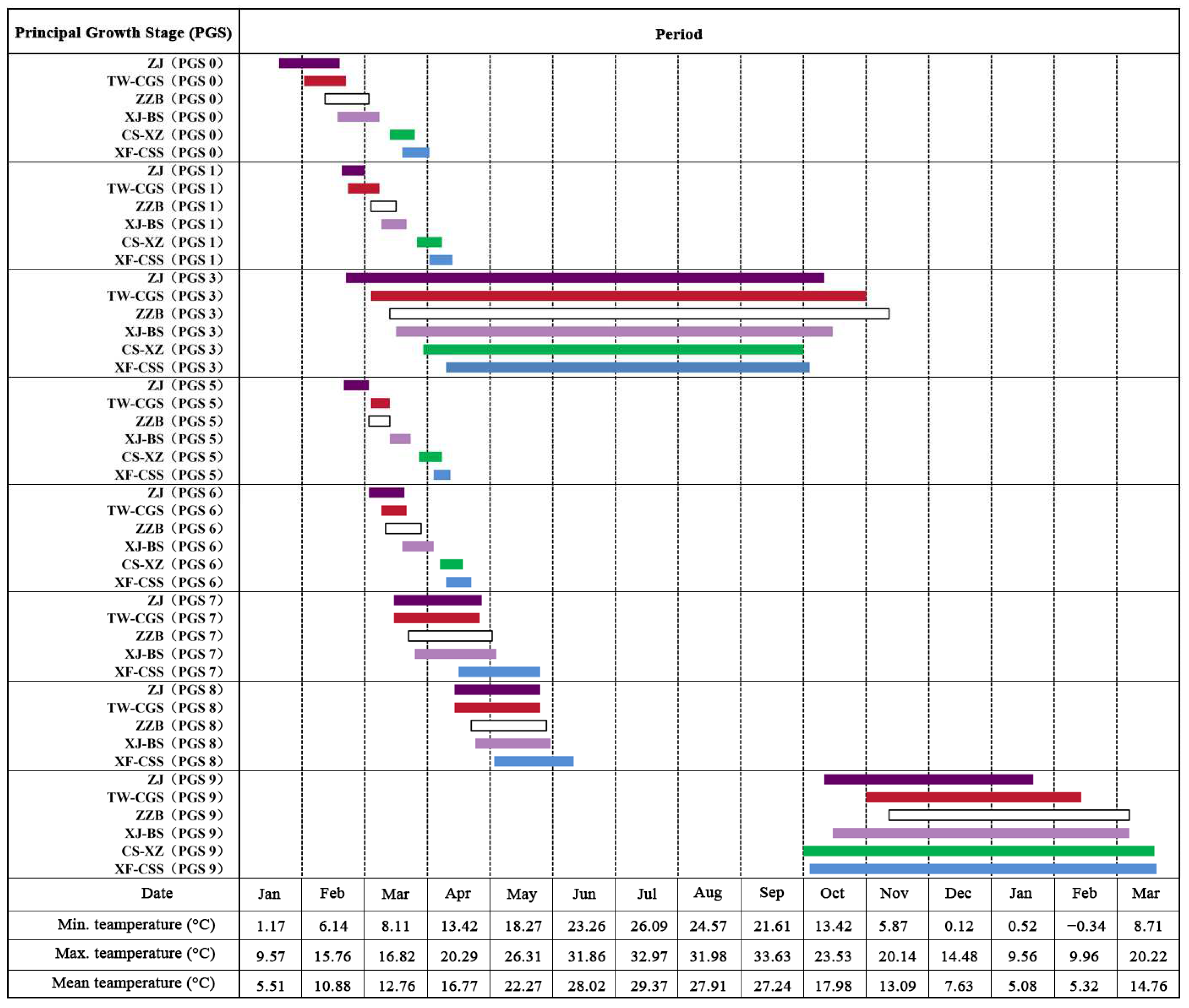
3.1. Principal Growth Stage 0: Bud Development (Figure 1)
3.2. Principal Growth Stage 1: Leaf Development (Figure 2)
3.3. Principal Growth Stage 3: Shoot Development (Figure 3)
3.4. Principal Growth Stage 5: Inflorescence Emergence (Figure 4)
3.5. Principal Growth Stage 6: Flowering (Figure 5)
3.6. Principal Growth Stage 7: Fruit Development (Figure 6)
3.7. Principal Growth Stage 8: Maturity of Fruit (Figure 6)
3.8. Principal Growth Stage 9: Senescence and Beginning of the Rest Period
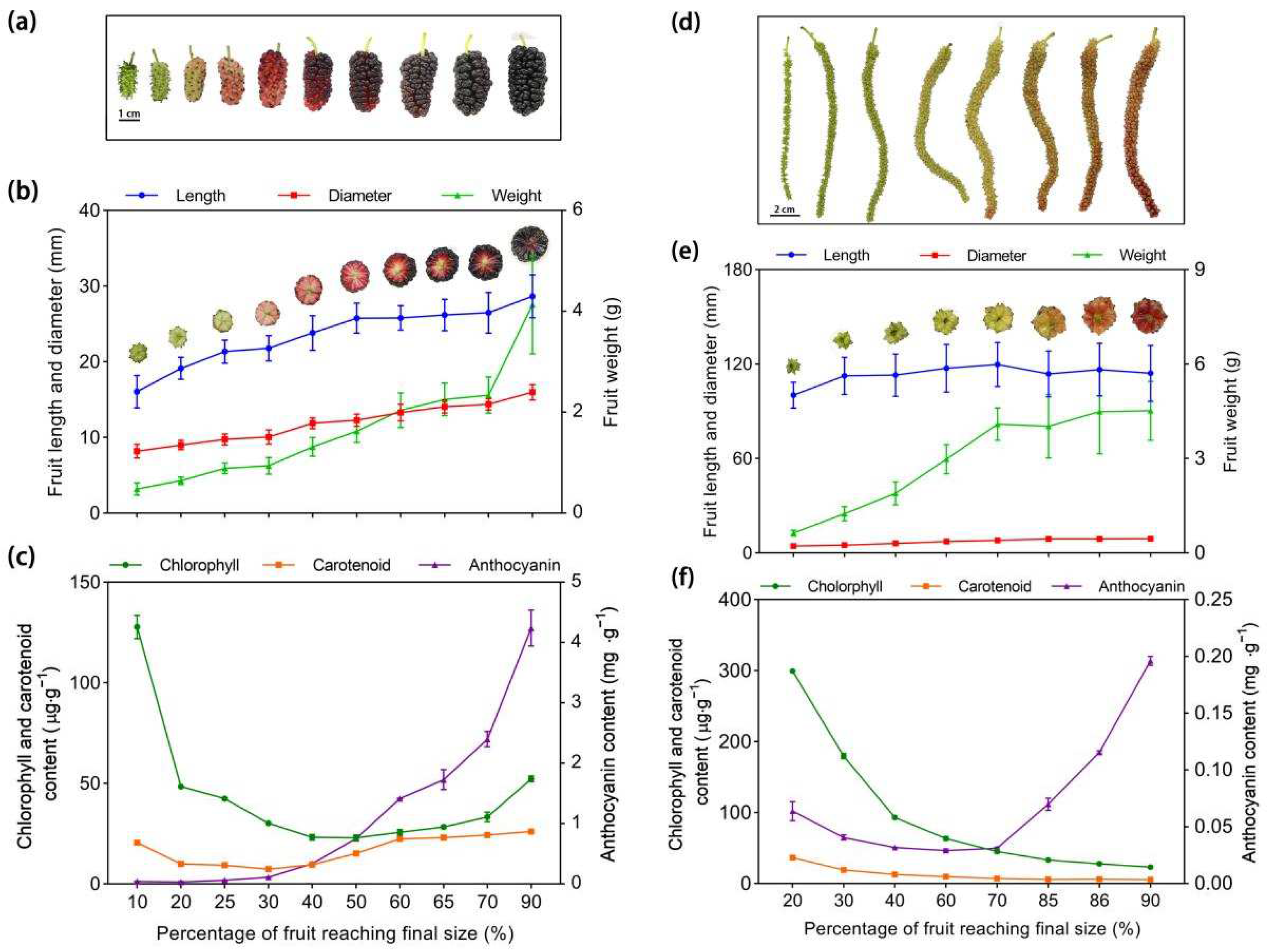
3.9. Morphological Profiles and Color Characters of the Mulberry Fruits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vijayan, K.; Ravikumar, G.; Tikader, A. Mulberry (Morus spp.) breeding for higher fruit production. In Advances in Plant Breeding Strategies: Fruits; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer: New York, NY, USA, 2018; Volume 3, pp. 89–130. [Google Scholar]
- Sharma, A.; Sharma, R.; Machii, H. Assessment of genetic diversity in a Morus germplasm collection using fluorescence-based AFLP markers. Theor. Appl. Genet. 2000, 101, 1049–1055. [Google Scholar] [CrossRef]
- Singhal, B.K.; Khan, M.A.; Dhar, A.; Baqua, F.M.; Bindroo, B.B. Approaches to industrial exploitation of Mulberry (Mulberry sp.) fruits. J. Fruit Ornam. Plant Res. 2010, 18, 83–99. [Google Scholar]
- Gray, E. Evidence of phenotypic plasticity in mulberry (Morus L.). Castanea 1990, 55, 272–281. [Google Scholar]
- Dandin, S.B. Mulberry: A versatile biosource in the service of mankind. Acta Sericol. Sin. 1998, 24, 109–113. [Google Scholar]
- Bhattacharya, E.; Dandin, S.B.; Ranade, S.A. Single primer amplification reaction methods reveal exotic and indigenous mulberry varieties are similarly diverse. J. Biosci. 2005, 30, 669–677. [Google Scholar] [CrossRef]
- Zeng, Q.; Chen, H.; Zhang, C.; Han, M.; Li, T.; Qi, X.; Xiang, Z.; He, N. Definition of eight mulberry species in the genus Morus by internal transcribed spacer-based phylogeny. PLoS ONE 2015, 10, e0135411. [Google Scholar] [CrossRef] [Green Version]
- Linné, C. Morus. In Species Plantarum; Linné, C., Ed.; Stockholm Impensis Laurentii Salvii: Stockholm, Sweden, 1753; Volume 2, p. 968. [Google Scholar]
- Bureau, E. Moraceae. In Prodromus Systematis Naturalis Regni Vegetabilis; De Candolle, A., Ed.; Tuettel and Wurtz: Paris, France, 1873; Volume 17, pp. 211–288. [Google Scholar]
- Koidzume, G. Taxonomical discussion on Morus plants. Bull. Imp. Sericult. Exp. Stat. 1917, 3, 1–62. [Google Scholar]
- Zhou, Z.; Gilbert, M.G. Moraceae. In Flora of China; Wu, Z., Raven, P.H., Hong, D., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: Saint Louis, MO, USA, 2003; Volume 5, pp. 22–73. [Google Scholar]
- Chang, J.C. Taxonomy and cultivar improvement of mulberry in Taiwan. J. Taiwan Soc. Hort. Sci. 2006, 52, 377–392. [Google Scholar]
- Nepal, M.P.; Ferguson, C.J. Phylogenetics of Morus (Moraceae) inferred from ITS and trnL-trnF sequence data. Syst. Bot. 2012, 37, 442–450. [Google Scholar] [CrossRef] [Green Version]
- Taylor, P.E.; Card, G.; House, J.; Dickinson, M.H.; Flagan, R.C. High-speed pollen release in the white mulberry tree, Morus alba L. Sex. Plant Reprod. 2006, 19, 19–24. [Google Scholar] [CrossRef]
- Squirrell, J.; Hollingsworth, P.M.; Bateman, R.M.; Dickson, J.H.; Light, M.H.S.; Macconnaill, M.; Tebbit, M.C. Partitioning and diversity of nuclear and organelle markers in native and introduced populations of Epipactis helleborine (Orchidaceae). Am. J. Bot. 2001, 88, 1409–1418. [Google Scholar] [CrossRef]
- Valdes, L.L.S.; Borroto, O.G.; Perez, G.F. Effect of mulberry (Morus alba) on the rumen ecosystem. In Mulberry, Moringa and Tithonia in Animal Feed, and Other Uses. Results in Latin America and the Caribbean; Lourdes, L., Savon, V., Odilia, G.B., Gustavo, F.P., Eds.; Food and Agriculture Organization of the United Nations Instituto de Ciencia Animal: Havana, Cuba, 2017; pp. 87–92. [Google Scholar]
- Jolly, M.S.; Dandin, S.B.; Ravindran, S.; Kumar, R. Sexual polymorphism in the genus Morus L. Proc. Indian Acad. Sci. 1986, 96, 315–320. [Google Scholar] [CrossRef]
- Minamizawa, K. Experimental studies on the sex differentiation in mulberry. Bull. Fac. Agri. 1963, 7, 1–47. [Google Scholar]
- Bajpai, P.K.; Warghat, A.R.; Yadav, A.; Kant, A.; Srivastava, R.B.; Stobdan, T. High phenotypic variation in Morus alba L. along an altitudinal gradient in the Indian trans-Himalaya. J. Mt. Sci. 2015, 12, 446–455. [Google Scholar] [CrossRef]
- Lee, Y.; Hwang, K.T. Changes in physicochemical properties of mulberry fruits (Morus alba L.) during ripening. Sci. Hortic. 2017, 217, 189–196. [Google Scholar] [CrossRef]
- Liu, Y.; Willison, J.H.M. Prospects for cultivating white mulberry (Morus alba) in the drawdown zone of the Three Gorges Reservoir, China. Environ. Sci. Pollut. Res. 2013, 20, 7142–7151. [Google Scholar] [CrossRef]
- Zhang, H.; Li, X.; Zhang, S.; Yin, Z.; Zhu, W.; Li, J.; Meng, L.; Zhong, H.; Xu, N.; Wu, Y.; et al. Rootstock alleviates salt stress in grafted mulberry seedlings: Physiological and PSII function responses. Front. Plant Sci. 2018, 9, 1806. [Google Scholar] [CrossRef]
- Meier, U. Growth Stages of Mono- and Dicotyledonous Plants. BBCH Monograph, 2nd ed.; Federal Biological Research Centre for Agriculture and Forestry: Quedlinburg, Germany, 2001; p. 158. [Google Scholar]
- Leather, S.R. Precise knowledge of plant growth stages enhances applied and pure research. Ann. Appl. Biol. 2010, 157, 159–161. [Google Scholar] [CrossRef]
- Yang, C.; Zhan, Q.; Gao, S.; Liu, H. Characterizing the spatial and temporal variation of the land surface temperature hotspots in Wuhan from a local scale. Geo-Spat. Inf. Sci. 2020, 23, 327–340. [Google Scholar] [CrossRef]
- Xu, H.; Chen, H.; Zhou, X.; Wu, Y.; Liu, Y. Research on the relationship between urban morphology and air temperature based on mobile measurement: A case study in Wuhan, China. Urban Clim. 2020, 34, 100671. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. Characterization and measurement of anthocyanins by UV–visible spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F1.2.1–F1.2.12. [Google Scholar] [CrossRef]
- Richardson, E.A.; Seely, S.D.; Walker, D.R. A model for estimating the completion of rest for ‘Redhaven’ and ‘Elberta’ peach trees. HortScience 1974, 9, 331–332. [Google Scholar] [CrossRef]
- Xu, W.; Wang, F.; Wang, R.; Sui, Y.; Zhou, Z.; Xie, J.; Xiang, Z. Seasonal Characterization of the endophytic fungal microbiome of mulberry (Morus spp.) cultivars resistant and susceptible to sclerotiniosis. Microorganisms 2021, 9, 2052. [Google Scholar] [CrossRef]
- Martínez-Calvo, J.; Badenes, M.L.; Llácer, G.; Bleiholder, H.; Hack, H.; Meier, U. Phenological growth stages of loquat tree (Eriobotrya japonica (Thunb.) Lindl.). Ann. Appl. Biol. 1999, 134, 353–357. [Google Scholar] [CrossRef]
- Salinero, M.C.; Vela, P.; Sainz, M.J. Phenological growth stages of kiwifruit (Actinidia deliciosa ‘Hayward’). Sci. Hortic. 2009, 121, 27–31. [Google Scholar] [CrossRef]
- Hernández, F.C.A.; Legua, P.; Melgarejo, P.; Martínez, R.; Martínez, J.J. Phenological growth stages of jujube tree (Ziziphus jujube): Codification and description according to the BBCH scale. Ann. Appl. Biol. 2015, 166, 136–142. [Google Scholar] [CrossRef]
- Sánchez-Salcedo, E.M.; Martínez-Nicolás, J.J.; Hernández, F. Phenological growth stages of mulberry tree (Morus sp.) codification and description according to the BBCH scale. Ann. Appl. Biol. 2017, 171, 441–450. [Google Scholar] [CrossRef]
- Martínez, R.; Legua, P.; Martínez-Nicolás, J.J.; Melgarejo, P. Phenological growth stages of “Pero de Cehegín” (Malus domestica Borkh): Codification and description according to the BBCH scale. Sci. Hortic. 2019, 246, 826–834. [Google Scholar] [CrossRef]
- Guan, C.; Che, Q.; Zhang, P.; Huang, J.; Chachar, S.; Ruan, X.; Wang, R.; Yang, Y. Codification and description of growth stages in persimmon (Diospyros kaki Thunb.) using the extended BBCH scale. Sci. Hortic. 2021, 280, 109895. [Google Scholar] [CrossRef]
- Caranqui-Aldaz, J.M.; Romero-Saltos, H.; Hernández, F.; Martínez, R. Reproductive phenology of Vaccinium floribundum Kunth (Ericaceae) and codification according to the BBCH scale based on evidence from the volcano Chimborazo paramo (Ecuador). Sci. Hortic. 2022, 303, 111207. [Google Scholar] [CrossRef]
- Chu, Y.C.; Chang, J.C. Codification and description of the phenological growth stages of red-fleshed pitaya (Hylocereus polyrhizus) using the extended BBCH scale-with special reference to spines, areole, and flesh color development under field conditions. Sci. Hortic. 2022, 293, 110752. [Google Scholar] [CrossRef]
- Ferrer-Blanco, C.; Hormaza, J.I.; Lora, J. Phenological growth stages of “pawpaw” [Asimina triloba (L.) Dunal, Annonaceae] according to the BBCH scale. Sci. Hortic. 2022, 295, 110853. [Google Scholar] [CrossRef]
- Paradinas, A.; Ramade, L.; Mulot-Greffeuille, C.; Hamidi, R.; Thomas, M.; Toillon, J. Phenological growth stages of ‘Barcelona’hazelnut (Corylus avellana L.) described using an extended BBCH scale. Sci. Hortic. 2022, 296, 110902. [Google Scholar] [CrossRef]
- Yahiro, M.; Shinjo, T.; Yasuhiro, N. Chilling-requirements for breaking the dormancy in muberry-winter-bud in subtropics III. Effect of moderate low temperature at 17 °C or 20 °C for breaking the dormancy. Jpn. J. Trop. Agric. 1986, 30, 79–81. [Google Scholar]
- Yahiro, M.; Shinjo, T.; Tashiro, K. Chilling requirements in the breaking of dormancy in winter buds of four mulberry varieties. J. Sericult. Sci. Jpn. 1988, 57, 284–288. [Google Scholar]
- Makhoul, G.; Choumane, W.; Tiba, K.; Baroudi, H. Effect of chilling hour requirements in breaking buds dormancy in some al-shami mulberry (Morus nigra L.) genotypes. Tishreen Univ. J. Bio. Sci. Ser. 2021, 43, 211–227. [Google Scholar]
- Liu, Z.; Zhu, H.; Abbott, A. Dormancy behaviors and underlying regulatory mechanisms: From perspective of pathways to epigenetic regulation. In Advances in Plant Dormancy; Anderson, J.V., Ed.; Springer: New York, NY, USA, 2015; pp. 75–105. [Google Scholar]
- Chang, L.Y.; Li, K.T.; Yang, W.J.; Chang, J.C.; Chang, M.W. Phenotypic classification of mulberry (Morus) species in Taiwan using numerical taxonomic analysis through the characterization of vegetative traits and chilling requirements. Sci. Hortic. 2014, 176, 208–217. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Prueger, J.H. Temperature extremes: Effect on plant growth and development. Weather Clim. Extrem. 2015, 10, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Grainger, J. Studies upon the time of flowering of plants: Anatomical, floristic and phenological aspects of the problem. Ann. Appl. Biol. 1939, 26, 684–704. [Google Scholar] [CrossRef]
- Chang, J.C.; Chang, M.W. ‘Elongated fruit No. 1’ mulberry: An elite cultivar for fresh consumption. J. Am. Pomol. Soc. 2010, 64, 101–105. [Google Scholar]
- Chang, J.C. ‘Miaoli No. 1’ mulberry: A new cultivar for berry production. HortScience 2008, 43, 1594–1595. [Google Scholar] [CrossRef] [Green Version]
- Gungor, N.; Sengul, M. Antioxidant activity, total phenolic content and selected physicochemical properties of white mulberry (Morus alba L.) fruits. Int. J. Food Prop. 2008, 11, 44–52. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Li, L.; Xiao, G.; Limwachiranon, J.; Xu, Y.; Lu, H.; Yang, D.; Luo, Z. Effects of elevated CO2 on energy metabolism and γ-aminobutyric acid shunt pathway in postharvest strawberry fruit. Food Chem. 2018, 265, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef]
- Zhu, X.Y.; Chen, J.Y.; Qiu, K.; Kuai, B. Phytohormone and light regulation of chlorophyll degradation. Front. Plant Sci. 2017, 8, 1911. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Yang, Z.; Zeng, Q.W.; Wang, S.B.; Luo, Y.W.; Huang, Y.; Xin, Y.C.; He, N.J. Abnormal expression of bHLH3 disrupts aflavonoid homeostasis network, causing differences in pigment composition among mulberry fruits. Hortic. Res. 2020, 7, 83. [Google Scholar] [CrossRef]
- Yu, C.; Dong, Z.X.; Jemaa, E.; Zhu, Z.X.; Mo, R.L.; Li, Y.; Deng, W.; Hu, X.M.; Zhang, C.; Han, G.M. A feature selection approach guided an early prediction of anthocyanin accumulation using massive untargeted metabolomics data in mulberry. Plant Cell Physiol. 2022, 63, 671–682. [Google Scholar] [CrossRef]
- Mo, R.L.; Han, G.M.; Zhu, Z.X.; Essemine, J.; Dong, Z.X.; Li, Y.; Deng, W.; Qu, M.N.; Zhang, C.; Yu, C. The Ethylene Response Factor ERF5 Regulates Anthocyanin Biosynthesis in ‘Zijin’ Mulberry Fruits by Interacting with MYBA and F3H Genes. Int. J. Mol. Sci. 2022, 23, 7615. [Google Scholar] [CrossRef]
- Mo, R.L.; Zhang, N.; Li, J.X.; Jin, Q.; Zhu, Z.X.; Dong, Z.X.; Li, Y.; Zhang, C.; Yu, C. Transcriptomic analysis provides insights into anthocyanin accumulation in mulberry fruits. Horticulturae 2022, 8, 920. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Species | Abbreviation | Accession Number | Origin | Pedigree | Sexuality | Style Length | Sorosis Length | Sorosis Color |
|---|---|---|---|---|---|---|---|---|---|
| Zijing | M. multicaulis Perr. | ZJ | hbgs0001 | Wuhan, China | ‘Zhushan 3’ × ‘Yueyou 78’ | Female | Absent | Medium | Black |
| Zhenzhubai | M. alba L. | ZZB | hbgs0011 | Liqing, China | Local cultivar | Female | Very short | Medium | White |
| Xinjiang Baisang | M. alba L. | XJ-BS | hbzy0003 | Aksu, China | Local cultivar | Female | Very short | Medium | Purple |
| Taiwan Changguosang | M. laevigata Wall. | TW-CGS | hbzy0002 | Taiwan, China | Local cultivar | Female | Very short | Long | Red |
| Xianfeng Changsuisang | M. wittiorum var. mawa Koidz. | XF-CSS | hbzy0005 | Enshi, China | Local cultivar | Female | Short | Long | Red |
| Changsui Xiongzhu | M. laevigata Wall. | CS-XZ | hbzy0009 | Unknown, China | Local cultivar | Male |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mo, R.; Zhang, N.; Hu, D.; Jin, Q.; Li, J.; Dong, Z.; Zhu, Z.; Li, Y.; Zhang, C.; Yu, C. Identification of Phenological Growth Stages of Four Morus Species Based on the Extended BBCH-Scale and Its Application in Fruit Development with Morphological Profiles and Color Characteristics. Horticulturae 2022, 8, 1140. https://doi.org/10.3390/horticulturae8121140
Mo R, Zhang N, Hu D, Jin Q, Li J, Dong Z, Zhu Z, Li Y, Zhang C, Yu C. Identification of Phenological Growth Stages of Four Morus Species Based on the Extended BBCH-Scale and Its Application in Fruit Development with Morphological Profiles and Color Characteristics. Horticulturae. 2022; 8(12):1140. https://doi.org/10.3390/horticulturae8121140
Chicago/Turabian StyleMo, Rongli, Na Zhang, Die Hu, Qiang Jin, Jinxin Li, Zhaoxia Dong, Zhixian Zhu, Yong Li, Cheng Zhang, and Cui Yu. 2022. "Identification of Phenological Growth Stages of Four Morus Species Based on the Extended BBCH-Scale and Its Application in Fruit Development with Morphological Profiles and Color Characteristics" Horticulturae 8, no. 12: 1140. https://doi.org/10.3390/horticulturae8121140