
Genome-Wide Analysis of the AP2/ERF Family in Oily Persimmon (Diospyros oleifera) and Their Preliminary Roles Exploration in Response to Polyamines for Adventitious Root Formation in Cultivated Persimmon (D. kaki)
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of AP2/ERF Gene Family Members
2.2. Domain Identification and Gene Structure Analysis of AP2/ERF Family Members
2.3. AP2/ERF Gene Family Cluster Analysis
2.4. Prediction of cis-Acting Sites of AP2/ERF Gene Family
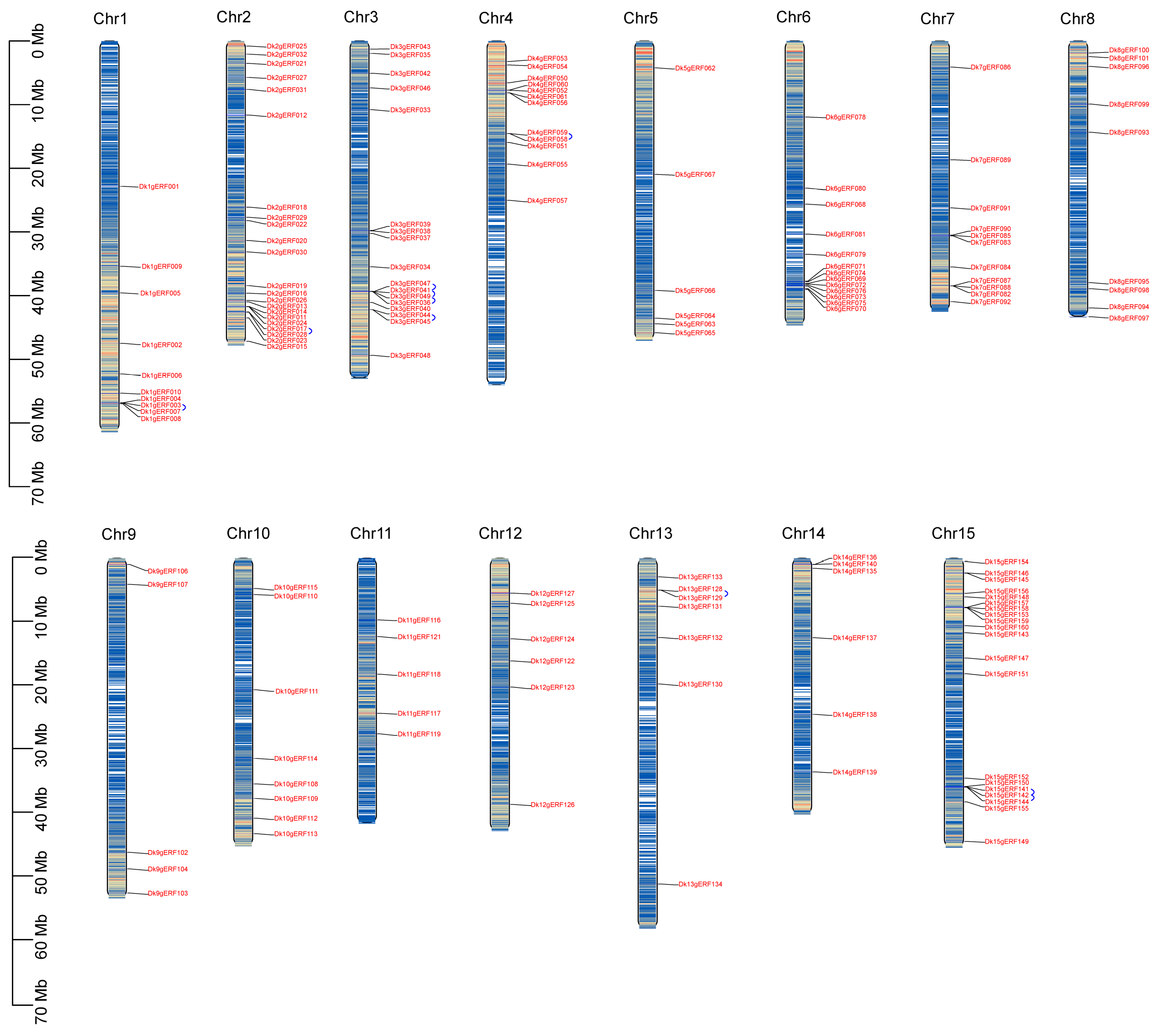
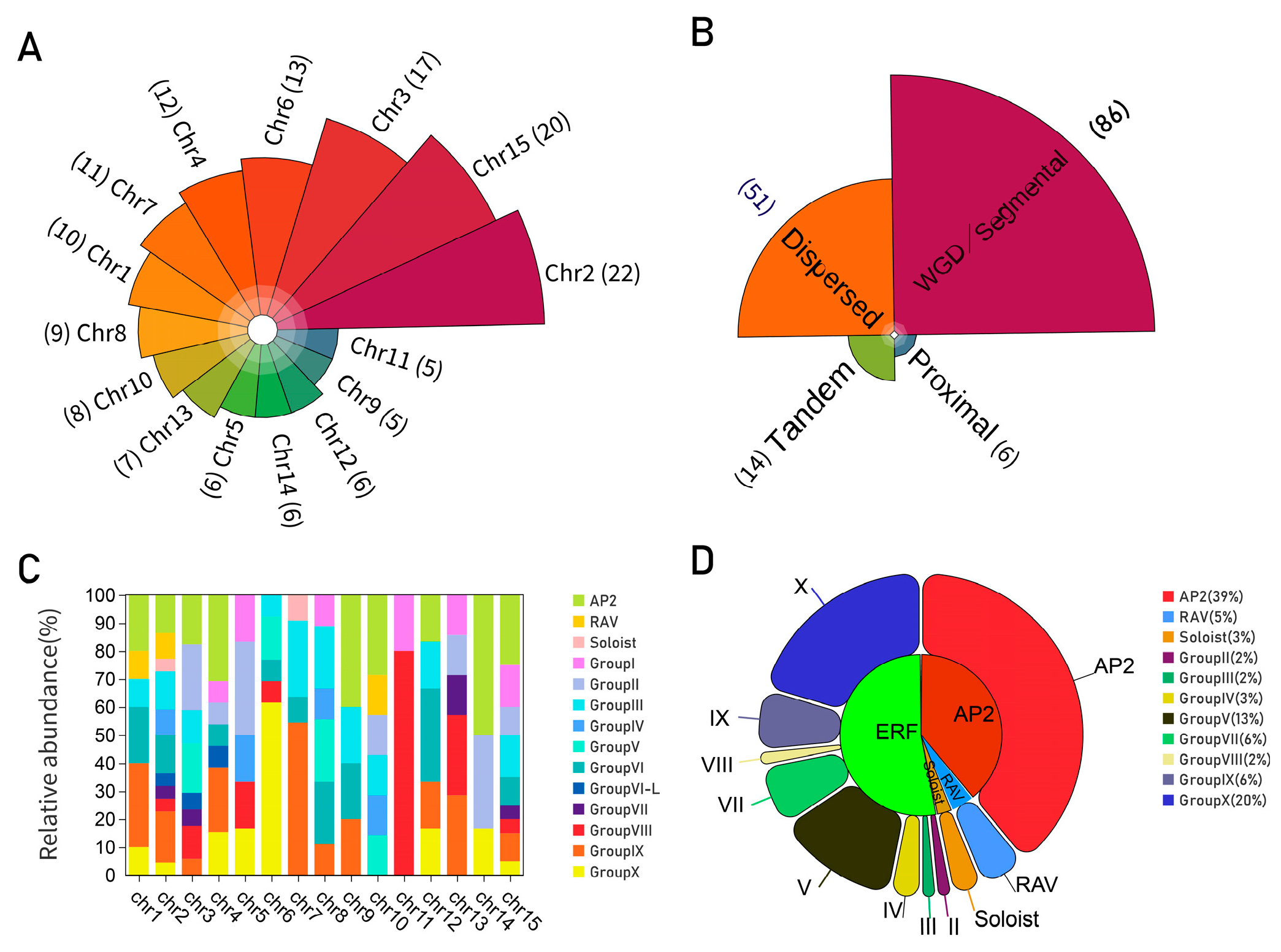
2.5. Chromosomal Distribution and Duplication of AP2/ERF Gene
2.6. Tissue-Specific Expression Analysis and Real-Time QUANTITATIVE Fluorescence PCR (qRT-PCR) Expression Detection
2.7. GO Enrichment and Protein Interaction (PPI) Analysis of AP2/ERF
3. Results
3.1. Identification and Sequence Information of AP2/ERF Gene Family Members in Oily Persimmon
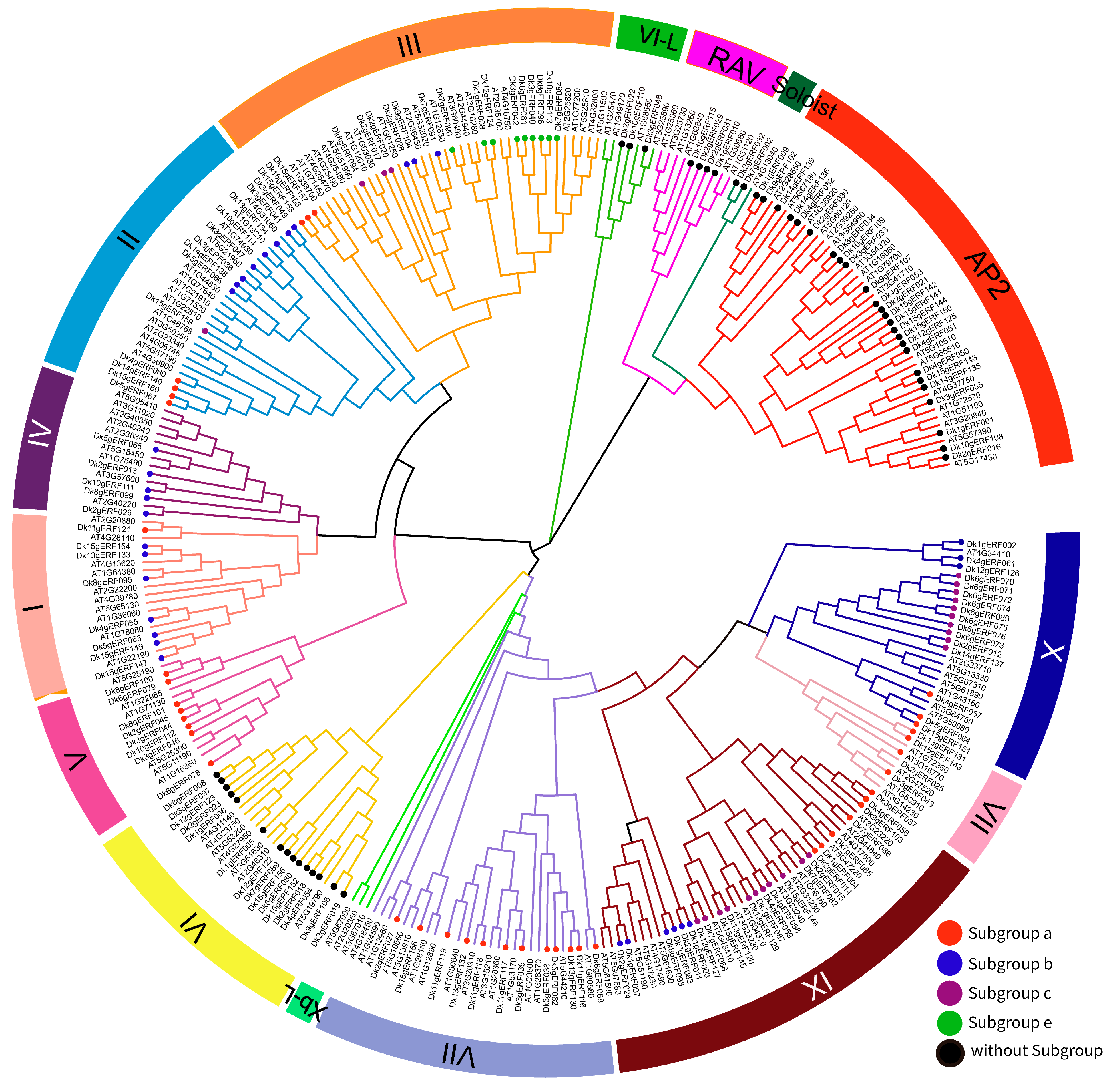
3.2. DkAP2/ERF Gene Family Classification and Evolutionary Analysis
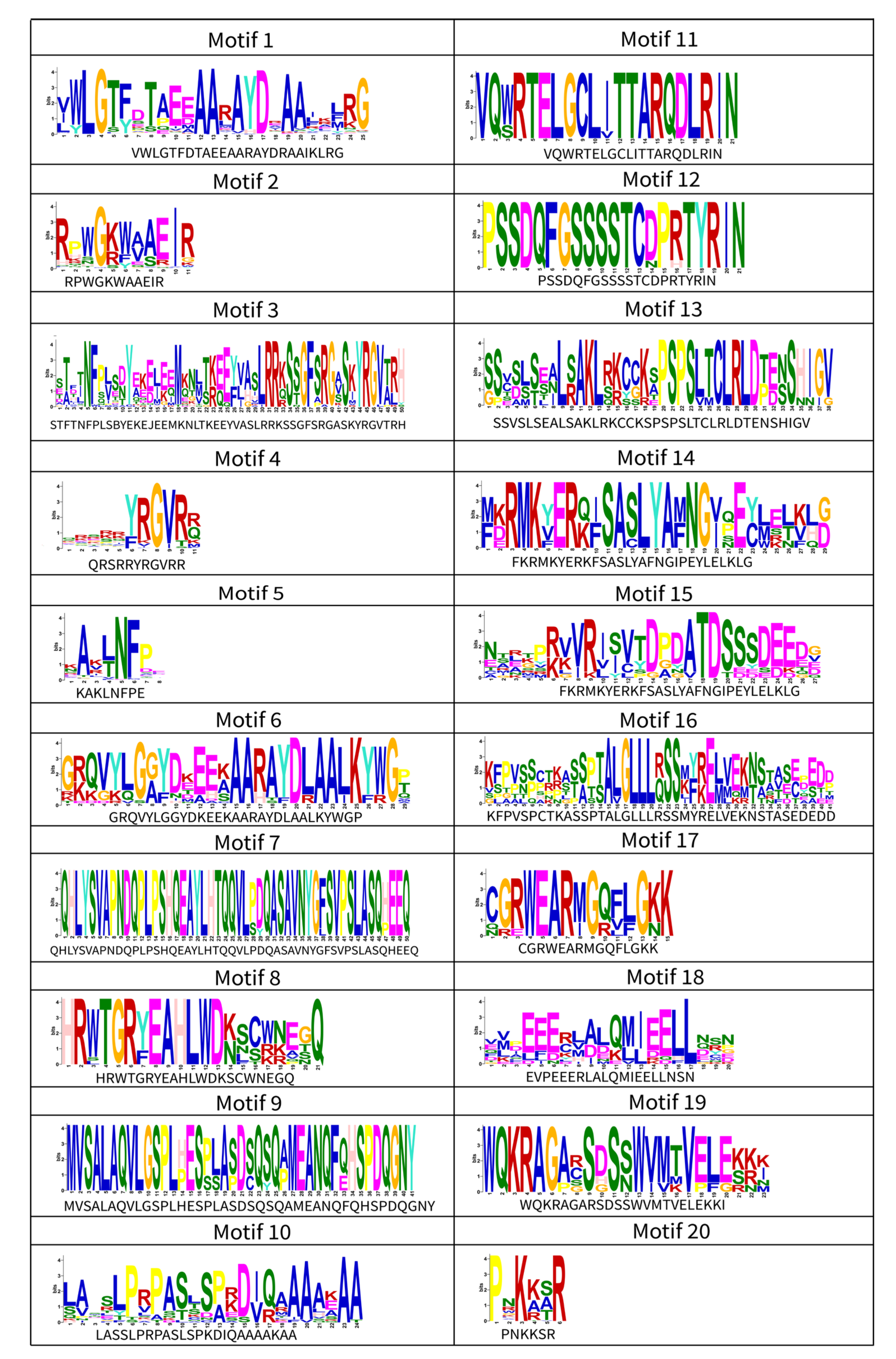
3.3. Gene Conserved Motif and Structure Analysis of the DkAP2/ERF Family
3.4. Analysis of cis-Acting Elements of DkAP2/ERF Gene Family
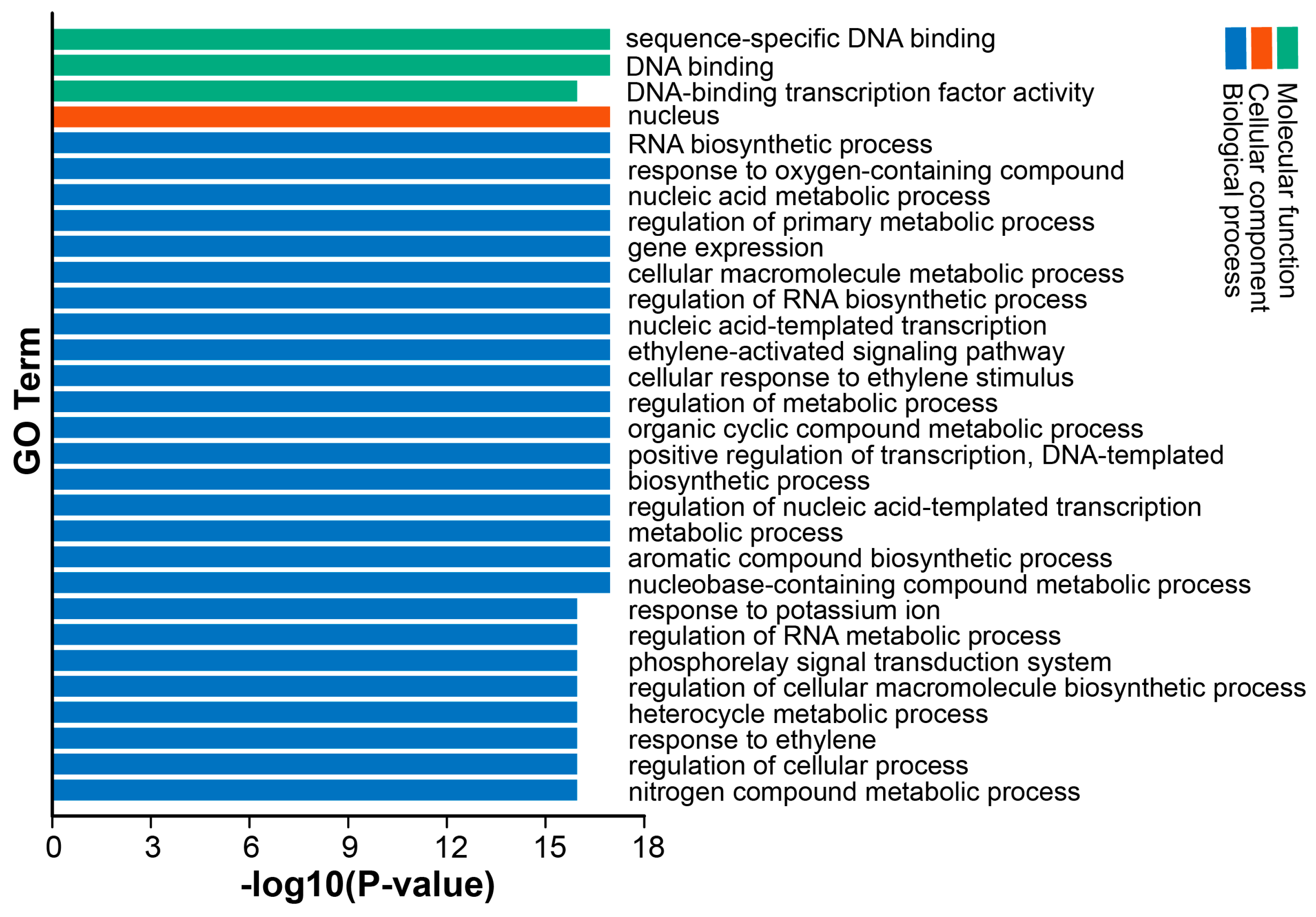
3.5. GO Analysis of DkAP2/ERF
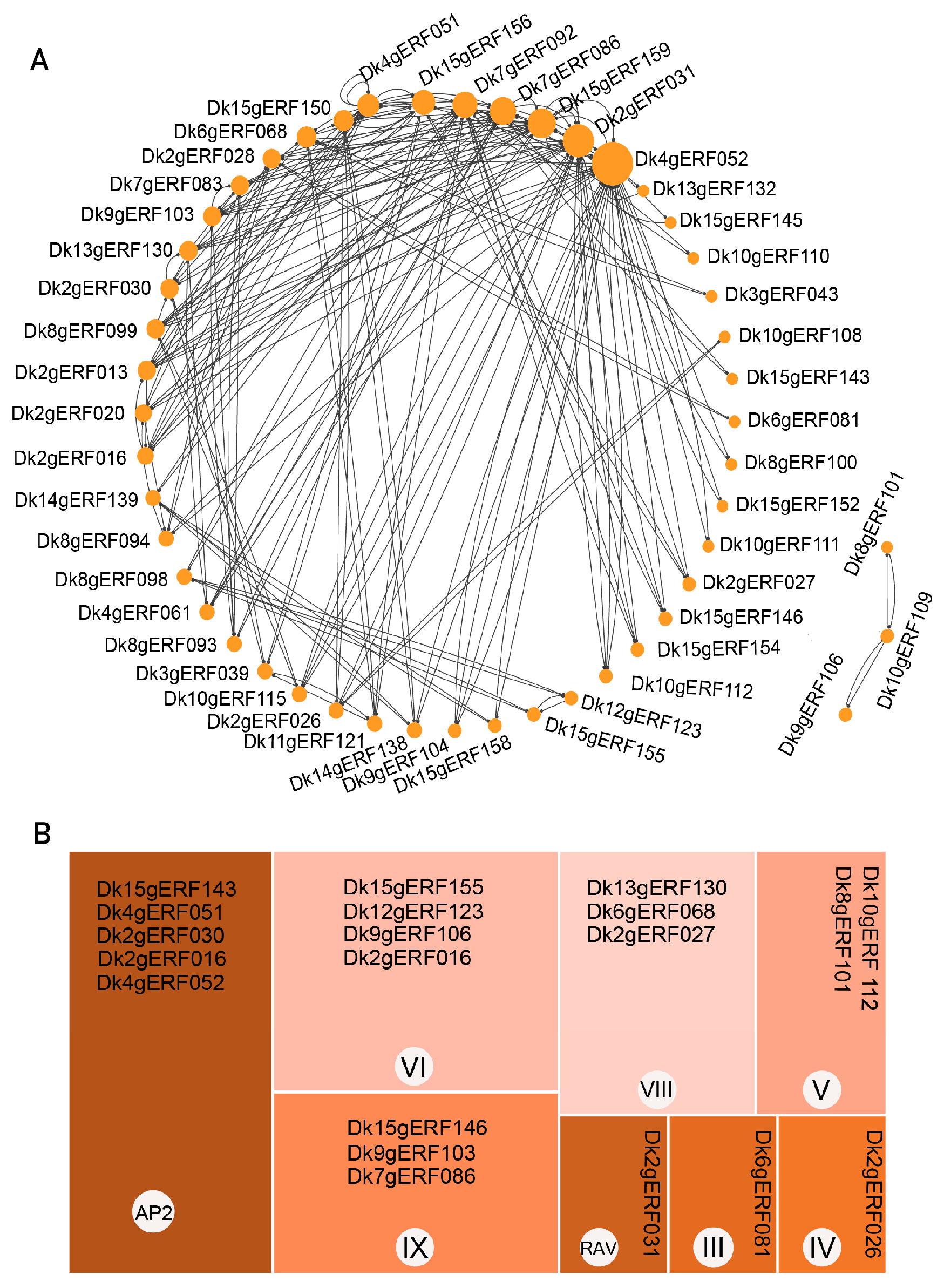
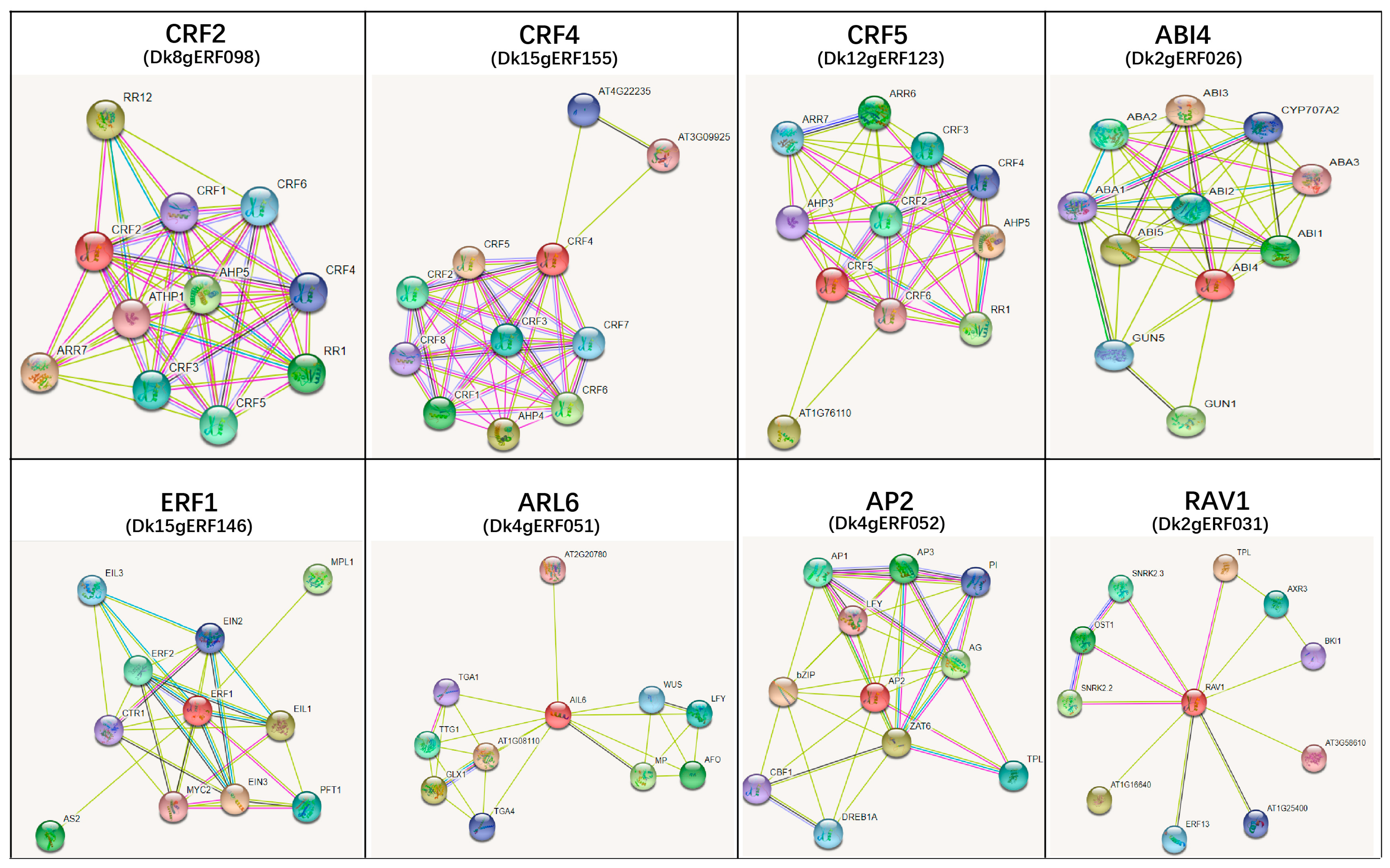
3.6. PPI Protein Prediction
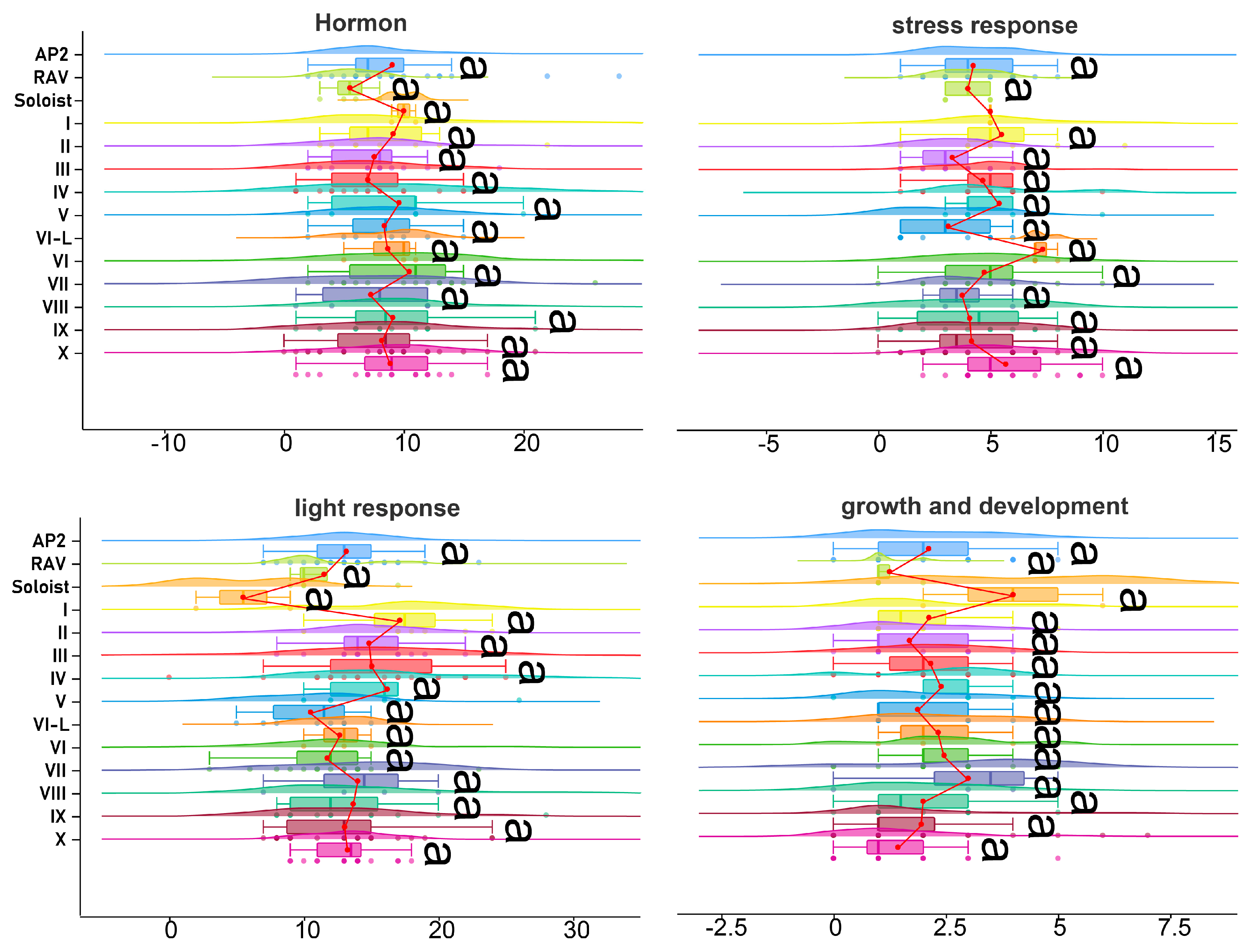
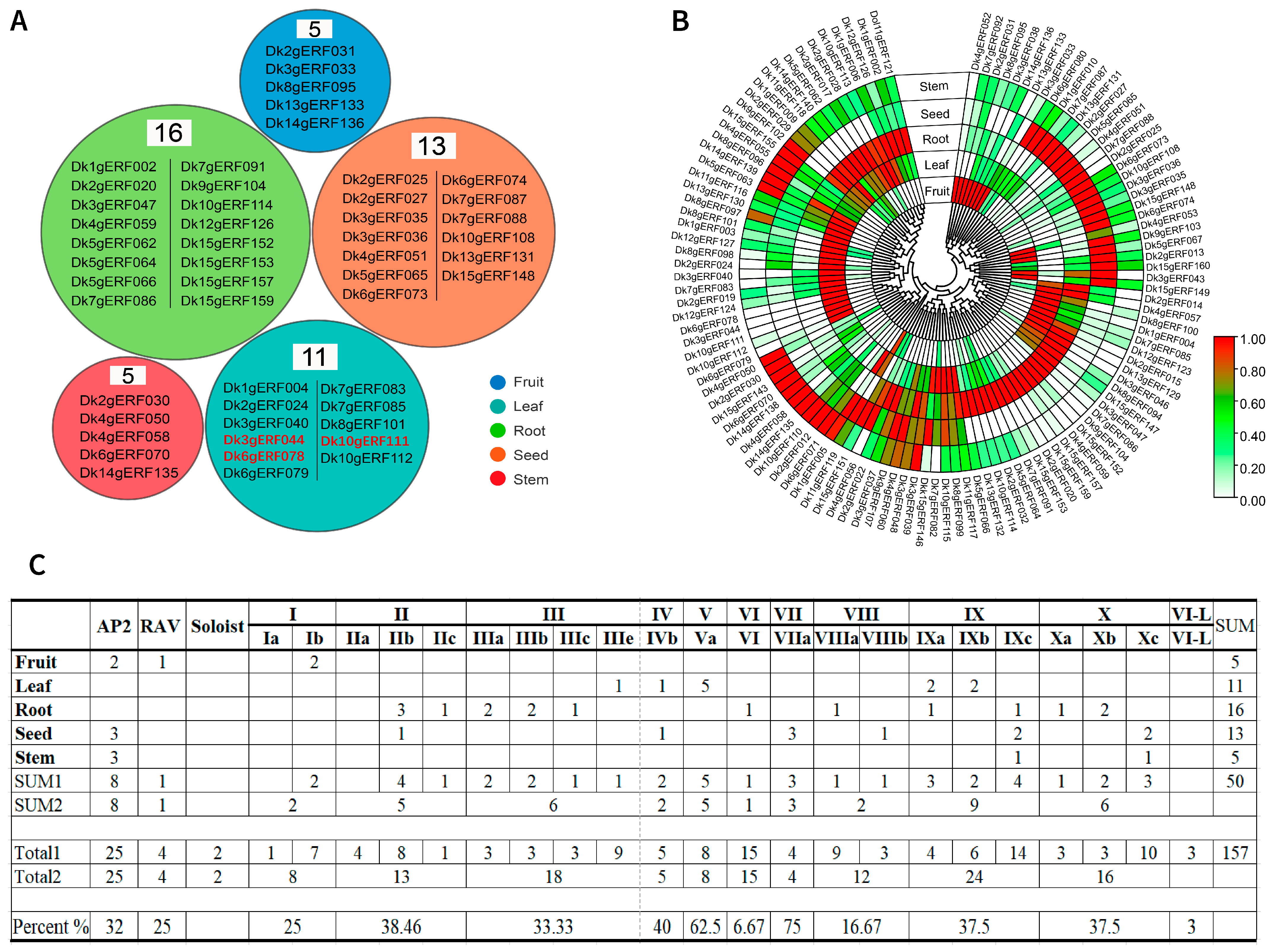
3.7. Tissue-Specific Expression Analysis of DkAP2/ERF
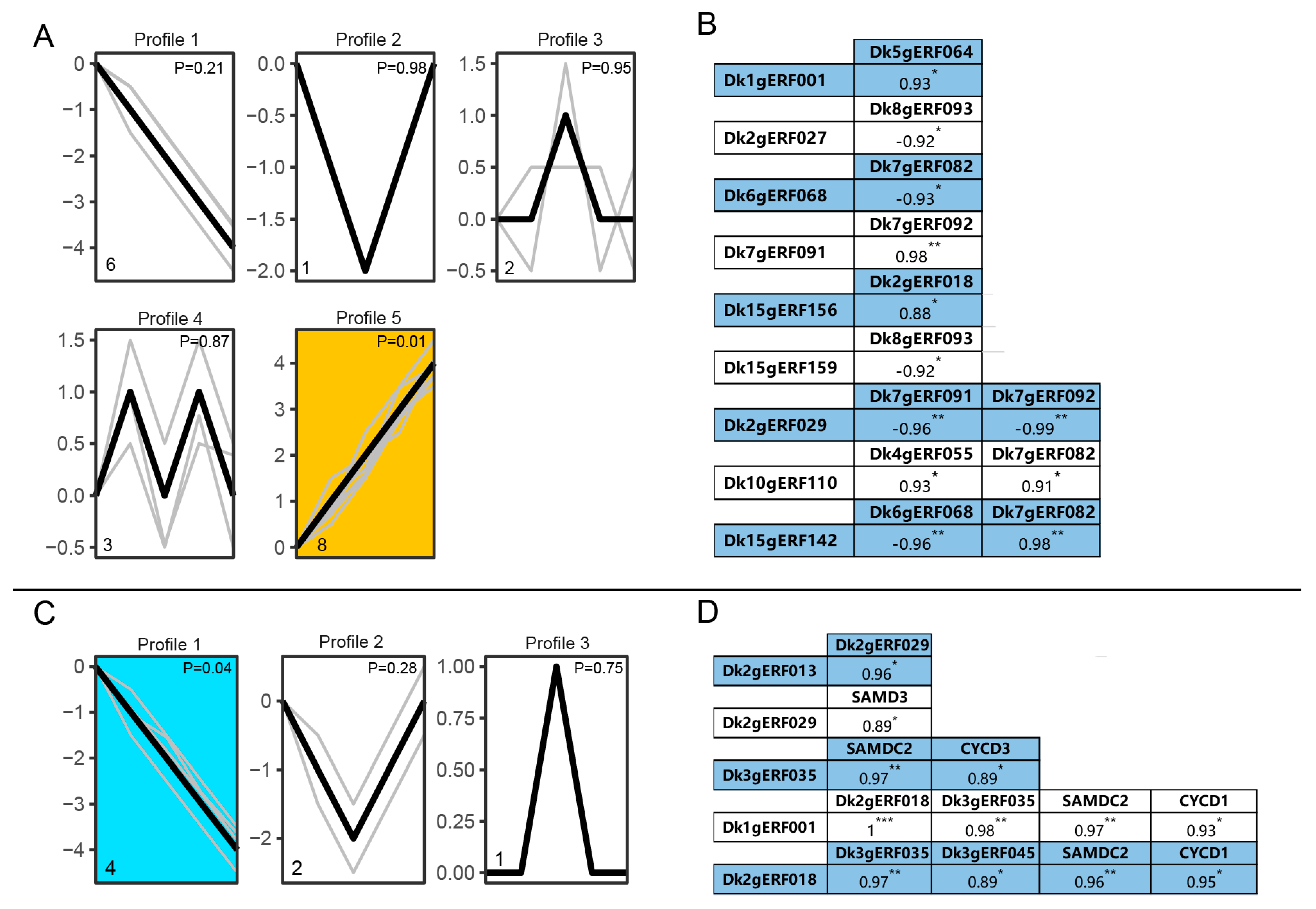
3.8. Expression Analysis of DkAP2/ERF Genes during Rooting and Development in Tissue Culture SEEDLINGS Treated with Polyamine
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Akagi, T.; Katayama-Ikegami, A.; Yonemori, K. Proanthocyanidin biosynthesis of persimmon (Diospyros Kaki Thunb). Fruit. Sci. Hortic--Amst. 2011, 130, 373–380. [Google Scholar] [CrossRef]
- Feng, K.; Hou, X.L.; Xing, G.M.; Liu, J.X.; Duan, A.Q.; Xu, Z.S.; Li, M.Y.; Zhuang, J.; Xiong, A.S. Advances in AP2/ERF super-family transcription factors in plant. Crit. Rev. Biotechnol. 2020, 40, 750–776. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-wide analysis of the ERF gene family in Arabidopsis and Rice. Plant Physiol. 2006, 140, 411–432. [Google Scholar] [CrossRef]
- Licausi, F.; Ohme-Takagi, M.; Perata, P. APETALA2/Ethylene responsive factor (AP2/ERF) transcription factors: Mediators of stress responses and developmental programs. New Phytol. 2013, 199, 639–649. [Google Scholar] [CrossRef]
- Karanja, B.K.; Xu, L.; Wang, Y.; Tang, M.; M’mbone Muleke, E.; Dong, J.; Liu, L. Genome-wide characterization of the AP2/ERF gene family in Radish (Raphanus Sativus L.): Unveiling evolution and patterns in response to abiotic stresses. Gene 2019, 718, 144048. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, K.; Peterson, K.; Jack, T. The plant B3 superfamily. Trends Plant Sci. 2008, 13, 647–655. [Google Scholar] [CrossRef]
- Najafi, S.; Sorkheh, K.; Nasernakhaei, F. Characterization of the APETALA2/Ethylene-responsive factor (AP2/ERF) transcription factor family in Sunflower. Sci. Rep. 2018, 8, 11576. [Google Scholar] [CrossRef]
- Kagale, S.; Rozwadowski, K. EAR motif-mediated transcriptional repression in plants: An underlying mechanism for epigenetic regulation of gene expression. Epigenetics 2011, 6, 141–146. [Google Scholar] [CrossRef]
- Tiwari, S.B.; Belachew, A.; Ma, S.F.; Young, M.; Ade, J.; Shen, Y.; Marion, C.M.; Holtan, H.E.; Bailey, A.; Stone, J.K.; et al. The EDLL motif: A potent plant transcriptional activation domain from AP2/ERF transcription factors. Plant J. 2012, 70, 855–865. [Google Scholar] [CrossRef]
- Tahir, M.M.; Mao, J.; Li, S.; Li, K.; Liu, Y.; Shao, Y.; Zhang, D.; Zhang, X. Insights into factors controlling adventitious root formation in Apples. Horticulturae 2022, 8, 276. [Google Scholar] [CrossRef]
- Trupiano, D.; Yordanov, Y.; Regan, S.; Meilan, R.; Tschaplinski, T.; Scippa, G.S.; Busov, V. Identification, characterization of an AP2/ERF transcription factor that promotes adventitious, lateral root formation in Populus. Planta 2013, 238, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Hirota, A.; Kato, T.; Fukaki, H.; Aida, M.; Tasaka, M. The auxin-regulated AP2/EREBP gene PUCHI is required for morphogenesis in the early lateral root primordium of Arabidopsis. Plant Cell 2007, 19, 2156–2168. [Google Scholar] [CrossRef] [PubMed]
- Neogy, A.; Garg, T.; Kumar, A.; Dwivedi, A.K.; Singh, H.; Singh, U.; Singh, Z.; Prasad, K.; Jain, M.; Yadav, S.R. Genome-wide transcript profiling reveals an auxin-responsive transcription factor, OsAP2/ERF-40, promoting rice adventitious root development. Plant Cell Physiol. 2019, 60, 2343–2355. [Google Scholar] [CrossRef] [PubMed]
- Rigal, A.; Yordanov, Y.S.; Perrone, I.; Karlberg, A.; Tisserant, E.; Bellini, C.; Busov, V.B.; Martin, F.; Kohler, A.; Bhalerao, R.; et al. The AINTEGUMENTA LIKE1 homeotic transcription factor PtAIL1 controls the formation of adventitious root primordia in Poplar. Plant Physiol. 2012, 160, 1996–2006. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.P.; Ravishankar, G.A. Role of polyamines in the ontogeny of plants and their biotechnological applications. Plant Cell, Tissue Organ Cult. 2002, 69, 1–34. [Google Scholar] [CrossRef]
- Suo, Y.; Sun, P.; Cheng, H.; Han, W.; Diao, S.; Li, H.; Mai, Y.; Zhao, X.; Li, F.; Fu, J. A high-quality chromosomal genome assembly of Diospyros Oleifera Cheng. Gigascience 2020, 9, giz164. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Bailey, T.L.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1994, 2, 28–36. [Google Scholar]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Yin, X.R.; Shi, Y.N.; Min, T.; Luo, Z.R.; Yao, Y.C.; Xu, Q.; Ferguson, I.; Chen, K.S. Expression of ethylene response genes during persimmon fruit astringency removal. Planta 2012, 235, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Min, T.; Yin, X.R.; Shi, Y.N.; Luo, Z.R.; Yao, Y.C.; Grierson, D.; Ferguson, I.B.; Chen, K.S. Ethylene-responsive transcription factors interact with promoters of ADH and PDC involved in persimmon (Diospyros kaki) fruit de-astringency. J. Exp. Bot. 2012, 63, 6393–6405. [Google Scholar] [CrossRef] [PubMed]
- Min, T.; Fang, F.; Ge, H.; Shi, Y.N.; Luo, Z.R.; Yao, Y.C.; Grierson, D.; Yin, X.Y.; Chen, K.S. Two novel anoxia-induced ethylene response factors that interact with promoters of deastringency-related genes from persimmon. PLoS ONE 2014, 9, e7043. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Choi, S.C.; Jung, S.; Cho, B.K.; Ahn, G.H.; Ryu, S.B. A transcriptome approach towards understanding fruit softening in persimmon. Front. Plant Sci. 2017, 8, 1556. [Google Scholar]
- Allen, M.D.; Yamasaki, K.; Ohme-Takagi, M.; Tateno, M.; Suzuki, M. A novel mode of DNA recognition by a beta-sheet revealed by the solution structure of the GCC-box binding domain in complex with DNA. EMBO J. 1998, 17, 5484–5496. [Google Scholar] [CrossRef] [PubMed]
- Licausi, F.; Giorgi, F.M.; Zenoni, S.; Osti, F.; Pezzotti, M.; Perata, P. Genomic and transcriptomic analysis of the AP2/ERF superfamily in Vitis Vinifera. BMC Genom. 2010, 11, 719. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, Y.; Wu, M.; Li, L.; Li, C.; Han, Z.; Yuan, J.; Chen, C.; Song, W.; Wang, C. Genome-Wide identification of AP2/ERF transcription factors in Cauliflower and expression profiling of the ERF Family under salt and drought stresses. Front. Plant Sci. 2017, 8, 946. [Google Scholar] [CrossRef]
- Liu, M.; Sun, W.; Ma, Z.; Zheng, T.; Huang, L.; Wu, Q.; Zhao, G.; Tang, Z.; Bu, T.; Li, C.; et al. Genome-wide investigation of the AP2/ERF gene family in tartary buckwheat (Fagopyum Tataricum). BMC Plant Biol. 2019, 19, 84. [Google Scholar] [CrossRef]
- Wang, H.; Zhou, P.; Liu, Y.; Han, J.J.; You, Q.; Chai, M.J.; Li, Y.; Tian, C.F.; Yang, Q.S.; Lu, J.X. Genome-wide analysis of AP2/ERF gene family in Chrysanthemums. Mol. Plant Breed. 2022, in press. [Google Scholar]
- Sharoni, A.M.; Nuruzzaman, M.; Satoh, K.; Shimizu, T.; Kondoh, H.; Sasaya, T.; Choi, I.R.; Omura, T.; Kikuchi, S. Gene structures, classification and expression models of the AP2/EREBP transcription factor family in Rice. Plant Cell Physiol. 2011, 52, 344–360. [Google Scholar] [CrossRef]
- Li, P.; Chai, Z.; Lin, P.; Huang, C.; Huang, G.; Xu, L.; Deng, Z.; Zhang, M.; Zhang, Y.; Zhao, X. Genome-wide identification and expression analysis of AP2/ERF transcription factors in Sugarcane (Saccharum Spontaneum L.). BMC Genom. 2020, 21, 685. [Google Scholar] [CrossRef]
- Kitomi, Y.; Ito, H.; Hobo, T.; Aya, K.; Kitano, H.; Inukai, Y. The Auxin responsive AP2/ERF transcription factor CROWN ROOTLESS5 is involved in crown root initiation in rice through the Induction of OsRR1, a type-A response regulator of cytokinin signaling. Plant J. 2011, 67, 472–484. [Google Scholar] [CrossRef] [PubMed]
- Andriankaja, A.; Boisson-Dernier, A.; Frances, L.; Sauviac, L.; Jauneau, A.; Barker, D.G.; de Carvalho-Niebel, F. AP2-ERF Transcription factors mediate nod factor dependent Mt ENOD11 activation in root hairs via a novel Cis-regulatory motif. Plant Cell 2007, 19, 2866–2885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.W. Molecular bases for the regulation of adventitious root generation in Plants. Front. Plant Sci. 2021, 12, 614072. [Google Scholar] [CrossRef]
- Chen, H.; Bullock, D.A.; Alonso, J.M.; Stepanova, A.N. To fight or to grow: The balancing role of ethylene in plant abiotic stress responses. Plants 2021, 11, 33. [Google Scholar] [CrossRef] [PubMed]
- Aukerman, M.J.; Sakai, H. Regulation of flowering time and floral organ identity by a microRNA and its APETALA2-like aarget genes. Plant Cell 2003, 15, 2730–2741. [Google Scholar] [CrossRef] [PubMed]
- Jofuku, K.D.; Omidyar, P.K.; Gee, Z.; Okamuro, J.K. Control of seed mass and Seed yield by the floral homeotic gene APETALA2. Proc. Natl. Acad. Sci. USA 2005, 102, 3117–3122. [Google Scholar] [CrossRef]
- Moose, S.P.; Sisco, P.H. Glossy15, an APETALA2-like gene from maize that regulates leaf epidermal cell identity. Genes Dev. 1996, 10, 3018–3027. [Google Scholar] [CrossRef]
- Shinshi, H.; Usami, S.; Ohme-Takagi, M. Identification of an ethylene-responsive region in the promoter of a tobacco class I chitinase gene. Plant Mol. Biol. 1995, 27, 923–932. [Google Scholar] [CrossRef]
- Müller, M.; Munné-Bosch, S. Ethylene Response Factors: A key regulatory hub in hormone and stress signaling. Plant Physiol. 2015, 169, 32–41. [Google Scholar] [CrossRef]
- Hao, D.; Ohme-Takagi, M.; Sarai, A. Unique Mode of GCC box recognition by the DNA-binding domain of ethylene-responsive element-binding factor (ERF domain) in plant. J. Biol. Chem. 1998, 273, 26857–26861. [Google Scholar] [CrossRef] [PubMed]
- Gutterson, N.; Reuber, T.L. Regulation of disease resistance pathways by AP2/ERF transcription factors. Curr. Opin. Plant Biol. 2004, 7, 465–471. [Google Scholar] [CrossRef]
- Sohn, K.H.; Lee, S.C.; Jung, H.W.; Hong, J.K.; Hwang, B.K. Expression and functional roles of the pepper pathogen-induced transcription factor RAV1 in bacterial disease resistance, and drought and salt Stress tolerance. Plant Mol. Biol. 2006, 61, 897–915. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, S.; Somssich, M.; Nakata, M.T.; Unda, F.; Atsuzawa, K.; Kaneko, Y.; Wang, T.; Bågman, A.M.; Gaudinier, A.; Yoshida, K.; et al. Complete substitution of a secondary cell wall with a primary cell wall in Arabidopsis. Nat. Plants 2018, 4, 777–783. [Google Scholar] [CrossRef]
- Tsafouros, A.; Denaxa, N.K.; Roussos, P.A. Chapter 12—Role of Polyamines in Adventitious root formation. In Environmental, Physiological and Chemical Controls of Adventitious Rooting in Cuttings; Plant Biology, Sustainability and Climate Change; Husen, A., Ed.; Academic Press: Cambridge, MA, USA, 2022; pp. 289–313. ISBN 978−0−323−90636−4. [Google Scholar]
- Du, X.Y.; Zhang, Q.L.; Luo, Z.R. Comparison of four molecular markers for genetic analysis in Diospyros L. (Ebenaceae). Plant Syst. Evol. 2009, 281, 171–181. [Google Scholar] [CrossRef]
- Fu, J.M.; Liu, H.M.; Hu, J.J.; Liang, Y.Q.; Liang, J.J.; Wuyun, T.N.; Tan, X.F. Five complete chloroplast genome sequences from Diospyros: Genome organization and comparative analysis. PLoS ONE 2016, 11, e0159566. [Google Scholar] [CrossRef] [PubMed]
- Guan, C.F.; Liu, S.Y.; Wang, M.K.; Ji, H.; Ruan, X.F.; Wang, R.Z.; Yang, Y. Comparative transcriptomic analysis reveals genetic divergence and domestication genes in Diospyros. BMC Plant Biol. 2019, 19, 227. [Google Scholar] [CrossRef]
- Werner, T.; Motyka, V.; Laucou, V.; Smets, R.; Van Onckelen, H.; Schmülling, T. Cytokinin-deficient transgenic Arabidopsis plants show multiple developmental alterations indicating opposite functions of cytokinins in the regulation of shoot and root meristem activity. Plant Cell 2003, 15, 2532–2550. [Google Scholar] [CrossRef]
- Lohar, D.P.; Schaff, J.E.; Laskey, J.G.; Kieber, J.J.; Bilyeu, K.D.; Bird, D.M. Cytokinins play opposite roles in lateral root formation, and nematode and rhizobial symbioses. Plant J. 2004, 38, 203–214. [Google Scholar] [CrossRef]
- Riefler, M.; Novak, O.; Strnad, M.; Schmülling, T. Arabidopsis cytokinin receptor mutants reveal functions in shoot growth, leaf senescence, seed size, germination, root development, and cytokinin metabolism. Plant Cell 2006, 18, 40–54. [Google Scholar] [CrossRef]
- Suzuki, T.; Sakurai, K.; Ueguchi, C.; Mizuno, T. Two types of putative nuclear factors that physically interact with histidine-containing phosphotransfer (hpt) domains, signaling mediators in Histo-asp phosphorelay, in Arabidopsis thaliana. Plant Cell Physiol. 2001, 42, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Hwang, L.; Chen, H.C.; Sheen, J. Two-component signal transduction pathways in Arabidopsis. Plant Physiol. 2002, 129, 500–515. [Google Scholar] [CrossRef] [PubMed]
- Schaller, G.E.; Kieber, J.J.; Shiu, S.H. Two-component signaling elements and histidyl-aspartyl phosphorelays. Arab. Book 2008, 6, e0112. [Google Scholar] [CrossRef] [Green Version]
- Jeon, J.; Cho, C.; Lee, M.R.; Van Binh, N.; Kim, J. CYTOKININ RESPONSE FACTOR2 (CRF2) and CRF3 regulate lateral root development in response to cold stress in Arabidopsis. Plant Cell 2016, 28, 1828–1843. [Google Scholar] [CrossRef]
- Druege, U.; Franken, P.; Lischewski, S.; Ahkami, A.H.; Zerche, S.; Hause, B.; Hajirezaei, M.R. Transcriptomic analysis reveals ethylene as stimulator and auxin as regulator of adventitious root formation in Petunia cuttings. Front. Plant Sci. 2014, 5, 494. [Google Scholar] [CrossRef]
- Druege, U.; Franken, P.; Hajirezaei, M.R. Plant hormone homeostasis, signaling, and function during adventitious root formation in cuttings. Front. Plant Sci. 2016, 7, 381. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GeneID | Protein Identifier | Interaction Protein in Network | Function Annotation |
|---|---|---|---|
| Dk8gERF098 | CRF2 | RR12/ATHP1 | Cytokinin signaling pathway |
| Dk12gERF123 | CRF5 | CRF2/CRF3/CRF5/CRF6/ARR6/ARR7/AHP3/AHP5/RR1 | Cytokinin signaling pathway |
| Dk15gERF155 | CRF4 | CRF1/CRF2/CRF3/CRF4/CRF5/CRF6/CRF7/CRF8/AHP4 | Cytokinin signaling pathway |
| Dk10gERF112 | SHN1 | Wax biosynthesis | |
| Dk9gERF106 | SHN2 | Wax biosynthesis | |
| Dk8gERF101 | SHN3 | Wax biosynthesis | |
| Dk15gERF143 | ANT | CYCD3;1/STM/WUS | Organs initiation and development |
| Dk6gERF081 | tny | Stimulate cytokinin biosynthesis | |
| Dk13gERF130 | ERF4 | TPL | Root elongation |
| Dk2gERF030 | TOE2 | BRX | Cell proliferation and elongation of the root |
| Dk9gERF103 | ERF-1 | Ethylene-induced inhibition of root growth | |
| Dk2gERF016 | BBM | AGL15/MYB18/WUS | Root development |
| Dk2gERF026 | ABI4 | ABI1/ABI2/ABI3/ABI5/ABA1/ABA2/ABA3 | ABA signaling pathway |
| Dk2gERF027 | ESR1 | MP/CUC1/WUS/CYP707A2 | Auxin/ABA metabolism |
| Dk15gERF146 | ERF1 | EIN2/EIN3/EIL1/CTR1/ERF2 | Ethylene signaling pathway |
| Dk6gERF068 | ERF8 | HKT1/MOR1 | Growth of root/cytokinin signaling pathway |
| Dk4gERF051 | AIL6 | AFO/WUS/MP/TG4/TGA1 | In response to AUX/IAA |
| Dk2gERF031 | RAV1 | SNRK2.2/SNRK2.3/OST1/AXR3 | In response to ABA/AUX/IAA |
| Dk4gERF052 | AP2 | TPL/ZAT6/CBF1/BZIP | Auxin signaling pathway |
| Dk7gERF086 | ERF13 | MYB15/AUX1/MYC2 | Root development/in response to ABA signaling pathway |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Du, X.; Liu, M.; Liu, X.; Zhao, L.; Cao, L.; Zhang, S.; Song, L.; Sun, Y.; Liu, D.; et al. Genome-Wide Analysis of the AP2/ERF Family in Oily Persimmon (Diospyros oleifera) and Their Preliminary Roles Exploration in Response to Polyamines for Adventitious Root Formation in Cultivated Persimmon (D. kaki). Horticulturae 2023, 9, 191. https://doi.org/10.3390/horticulturae9020191
Wang Y, Du X, Liu M, Liu X, Zhao L, Cao L, Zhang S, Song L, Sun Y, Liu D, et al. Genome-Wide Analysis of the AP2/ERF Family in Oily Persimmon (Diospyros oleifera) and Their Preliminary Roles Exploration in Response to Polyamines for Adventitious Root Formation in Cultivated Persimmon (D. kaki). Horticulturae. 2023; 9(2):191. https://doi.org/10.3390/horticulturae9020191
Chicago/Turabian StyleWang, Yanbo, Xiaoyun Du, Minxiao Liu, Xueqing Liu, Lingling Zhao, Luyao Cao, Shuo Zhang, Laiqing Song, Yanxia Sun, Daliang Liu, and et al. 2023. "Genome-Wide Analysis of the AP2/ERF Family in Oily Persimmon (Diospyros oleifera) and Their Preliminary Roles Exploration in Response to Polyamines for Adventitious Root Formation in Cultivated Persimmon (D. kaki)" Horticulturae 9, no. 2: 191. https://doi.org/10.3390/horticulturae9020191