
Latitudinal Characteristic Nodule Composition of Soybean-Nodulating Bradyrhizobia: Temperature-Dependent Proliferation in Soil or Infection?

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Locations
2.2. Soybean Cultivation
2.3. Nodule Sampling and Isolation of Rhizobia
2.4. Phylogenetic Analysis of the Rhizobia Based on Genes of 16S rRNA and 16S–23S rRNA Internal Transcribed Spacer (ITS) Region
2.5. Nucleotide Sequence Accession Numbers
2.6. Statistical Analysis
3. Results
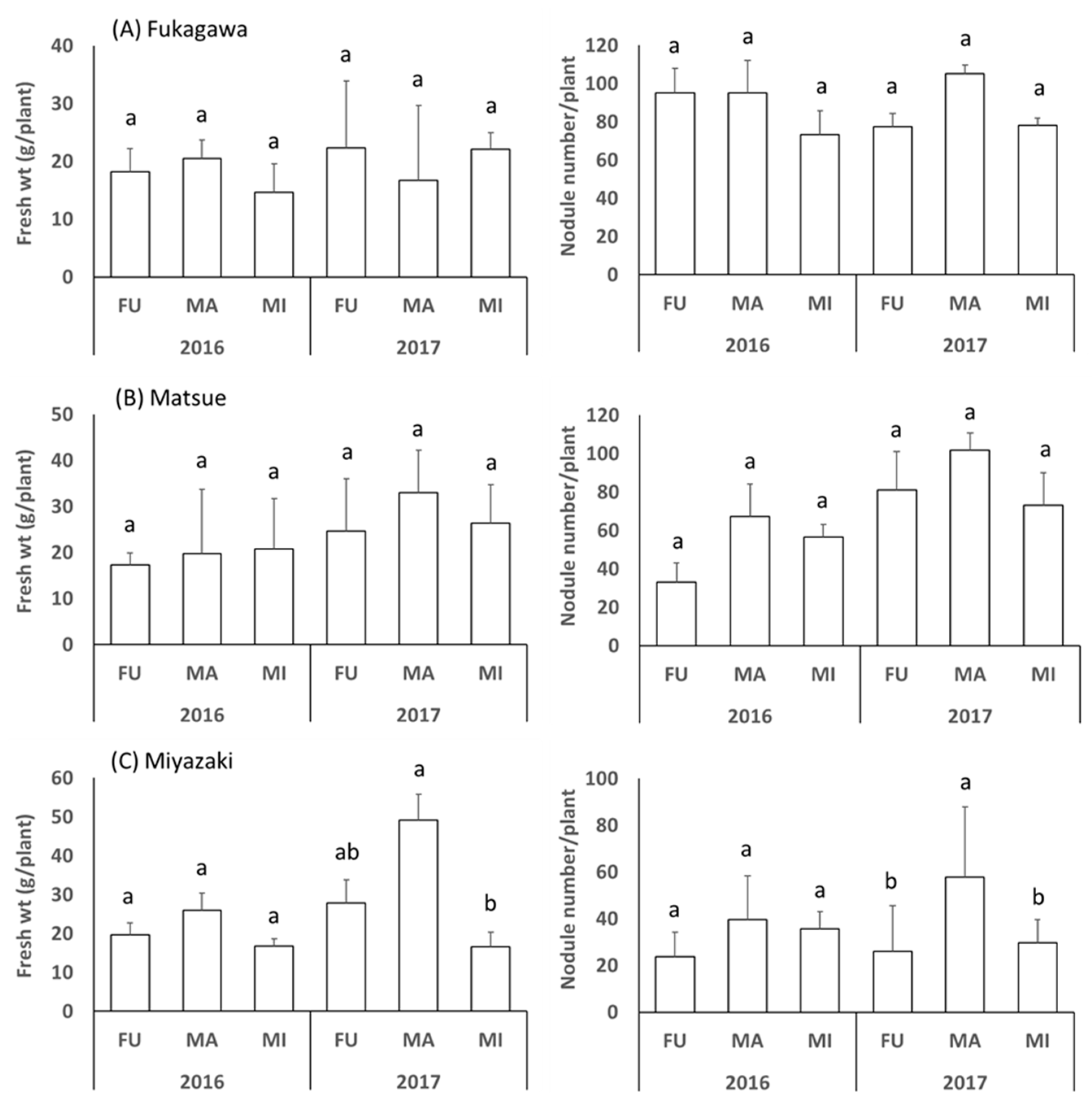
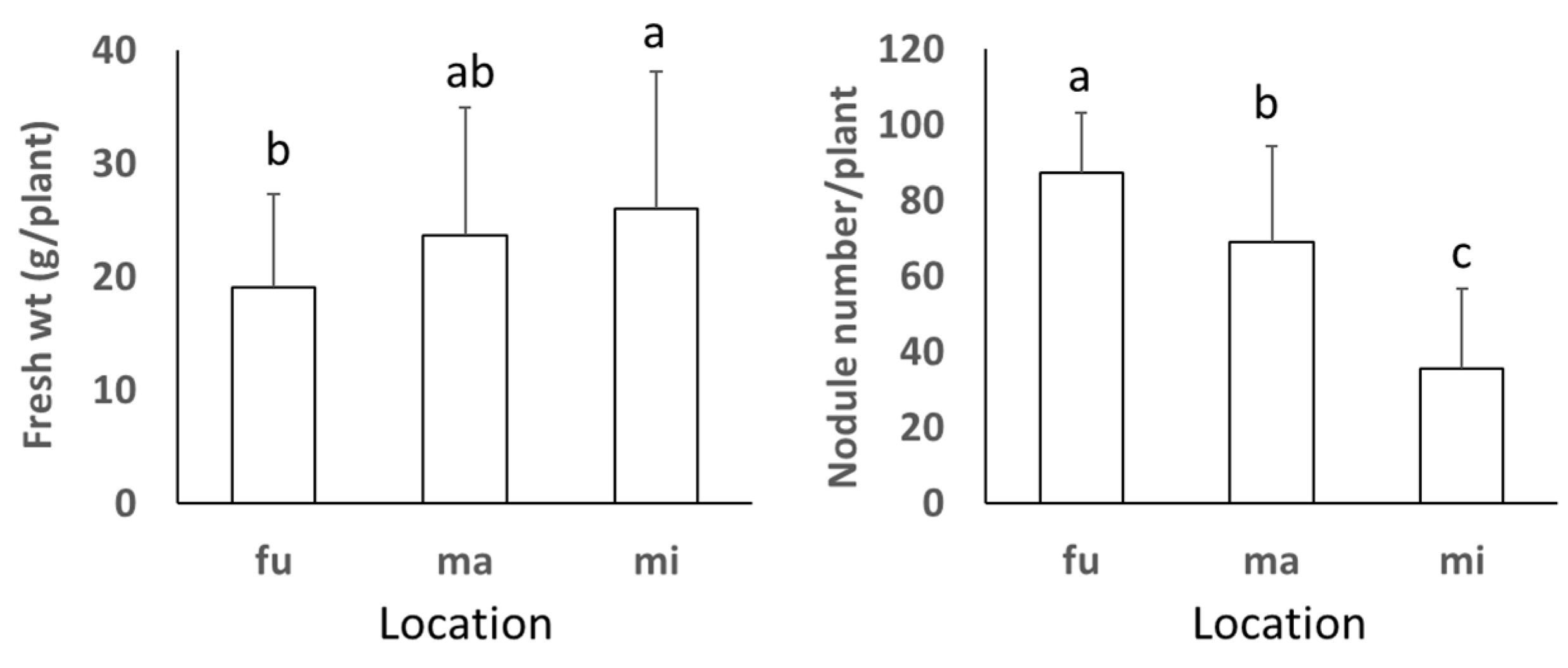
3.1. Fresh Plant Weight and Number of Nodules of Soybean
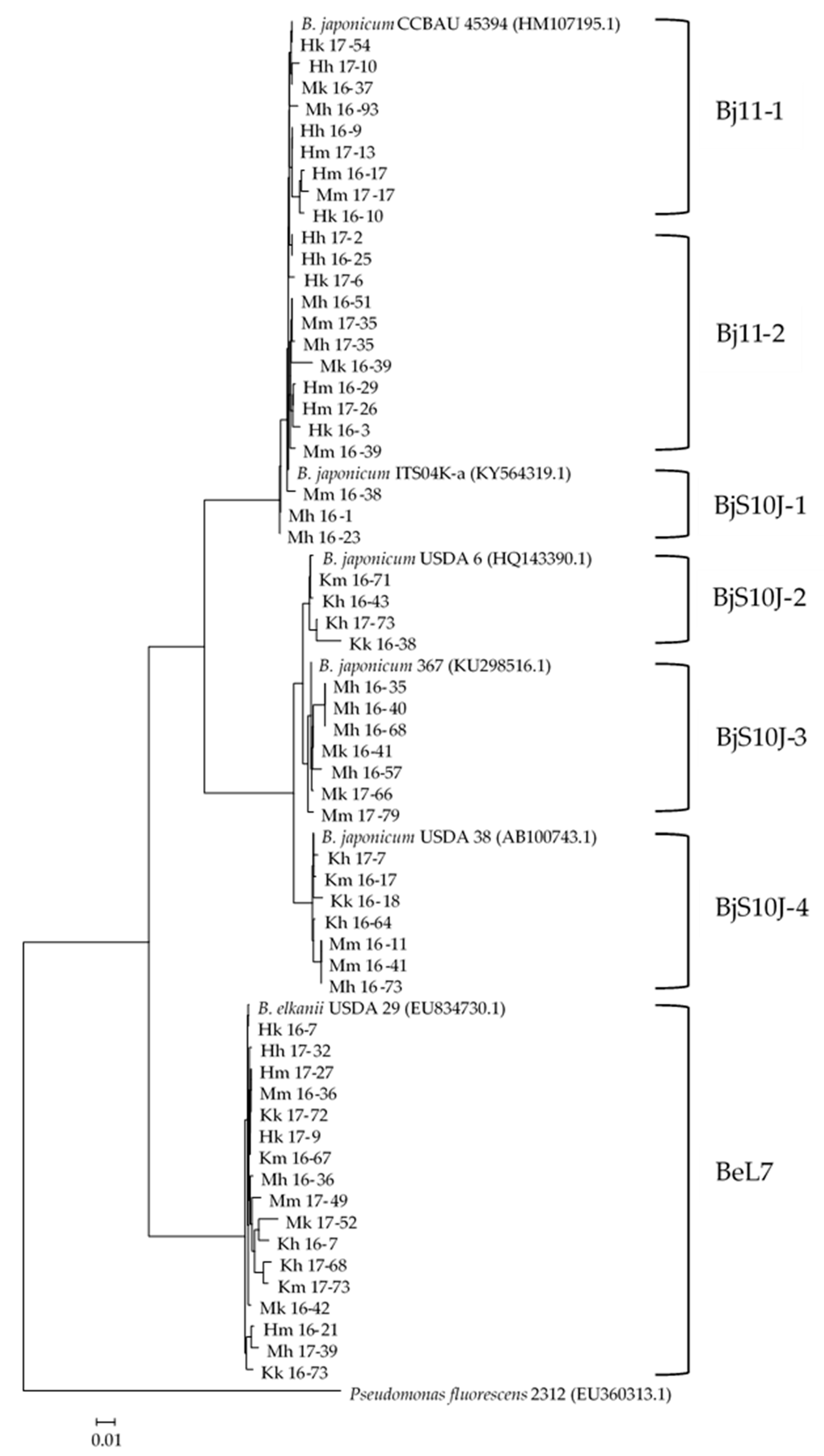
3.2. Phylogenetical Characterizations of the Rhizobia
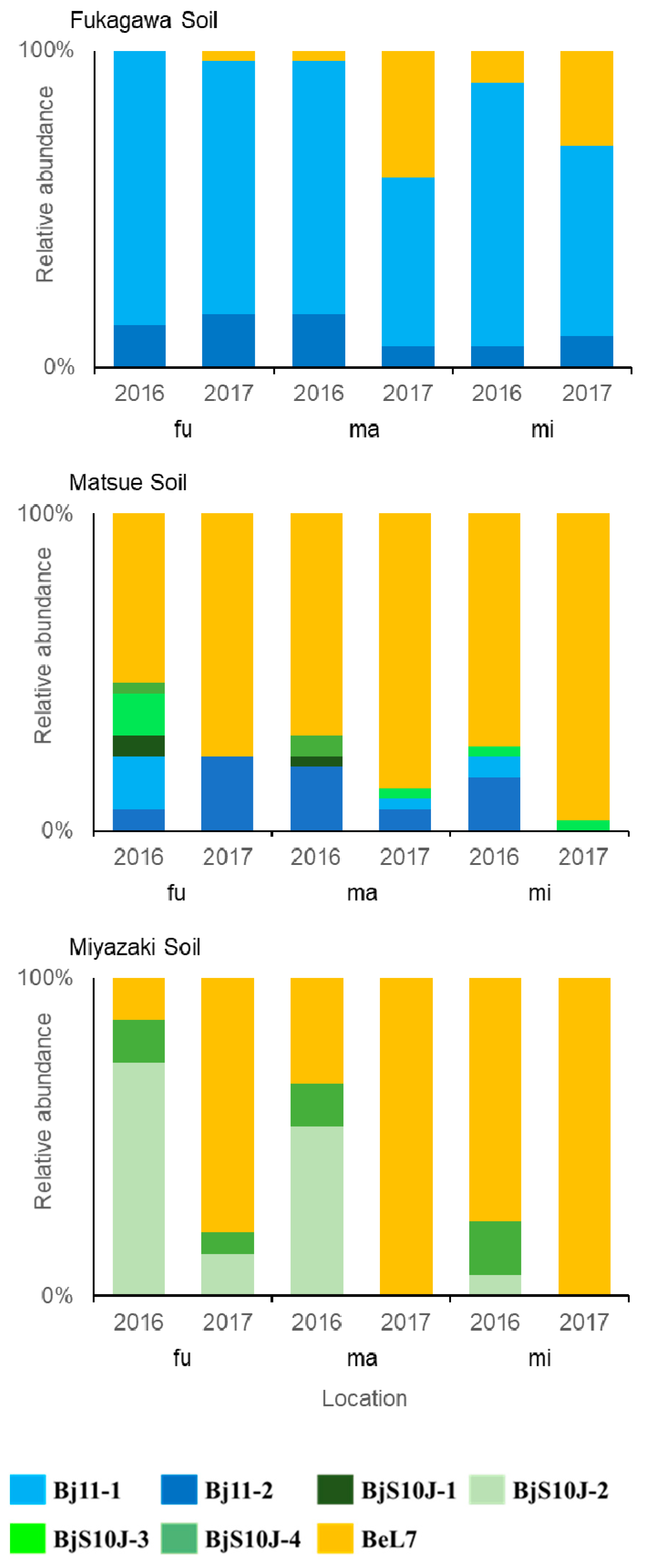
3.3. Relative Composition of the Strains in Relation to Soil and Climate in 2016 and 2017
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hymowitz, T.; Harlan, J.R. Introduction of soybean to north America by Samuel Bowen in 1765. Econ. Bot. 1983, 37, 371–379. [Google Scholar] [CrossRef]
- Dupare, B.U.; Billore, S.D.; Joshi, O.P.; Husain, S.M. Origin, domestication, introduction, and success of soybean in India. Asian Agrihist 2008, 12, 179–195. [Google Scholar]
- Khojely, D.M.; Ibrahim, S.E.; Sapey, E.; Han, T. History, current status, and prospects of soybean production and research in sub-Saharan Africa. Crop J. 2018, 6, 226–235. [Google Scholar] [CrossRef]
- Vinuesa, P.; Rojas-Jimenez, K.; Contreras-Moreira, B.; Mahna, S.K.; Prasad, B.N.; Moe, H.; Selvaraju, S.B.; Thierfelder, H.; Werner, D. Multilocus sequence analysis for assessment of the biogeography and evolutionary genetics of four Bradyrhizobium species that nodulate soybeans on the asiatic continent. Appl. Environ. Microbiol. 2008, 74, 6987–6996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Y.Z.; XU, X.D.; Wang, E.T.; Gao, J.I.; Martinez-Romero, E.; Chen, W.X. Phylogenetic and genetic relationships of Mesorhizobium tianshanense and related rhizobia. Int. J. Syst. Bacteriol. 1997, 47, 874–879. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wang, E.; Wang, S.; Li, Y.; Chen, X.; Li, Y. Characteristics of Rhizobium tianshanense sp. nov., a moderately and slowly growing root nodule bacterium isolated from an arid saline environment in Xinjiang, People’s Republic of China. Int. J. Syst. Bacteriol. 1995, 45, 153–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Streeter, J.G. Failure of inoculant rhizobia to overcome the dominance of indigenous strains for nodule formation. Can J. Microbiol. 1994, 40, 513–522. [Google Scholar] [CrossRef]
- Saeki, Y.; Aimi, N.; Tsukamoto, S.; Yamakawa, T.; Nagatomo, Y.; Akao, S. Diversity and geographical distribution of indigenous soybean-nodulating bradyrhizobia in Japan. Soil Sci. Plant Nutr. 2006, 52, 418–426. [Google Scholar] [CrossRef]
- Shiro, S.; Matsuura, S.; Saiki, R.; Sigua, G.C.; Yamamoto, A.; Umehara, Y.; Hayashi, M.; Saeki, Y. Genetic diversity and geographical distribution of indigenous soybean-nodulating bradyrhizobia in the United States. Appl. Environ. Microbiol. 2013, 79, 3610–3618. [Google Scholar]
- Adhikari, D.; Kaneto, M.; Itoh, K.; Suyama, K.; Pokharel, B.B.; Gaihre, Y.K. Genetic diversity of soybean-nodulating rhizobia in Nepal in relation to climate and soil properties. Plant Soil 2012, 357, 131–145. [Google Scholar] [CrossRef]
- Li, Q.Q.; Wang, E.T.; Zhang, Y.Z.; Zhang, Y.M.; Tian, C.F.; Sui, X.H.; Chen, W.F.; Chen, W.X. Diversity and biogeography of rhizobia isolated from root nodules of Glycine max grown in Hebei province, China. Microb. Ecol. 2011, 61, 917–931. [Google Scholar] [CrossRef] [PubMed]
- Kluson, R.A.; Kenworthy, W.J.; Weber, D.F. Soil temperature effects on competitiveness and growth of Rhizobium japonicum and on Rhizobium-induced chlorosis of soybeans. Plant Soil 1986, 95, 201–207. [Google Scholar] [CrossRef]
- Suzuki, Y.; Adhikari, D.; Itoh, K.; Suyama, K. Effects of temperature on competition and relative dominance of Bradyrhizobium japonicum and Bradyrhizobium elkanii in the process of soybean nodulation. Plant Soil 2014, 374, 915–924. [Google Scholar] [CrossRef]
- Shiro, S.; Kuranaga, C.; Yamamoto, A.; Sameshima-Saito, R.; Saeki, Y. Temperature-dependent expression of NodC and community structure of soybean-nodulating bradyrhizobia. Microbes Environ. 2016, 31, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Puri, R.R.; Adachi, F.; Omichi, M.; Saeki, Y.; Yamamoto, A.; Hayashi, S.; Itoh, K. Culture-dependent analysis of endophytic bacterial community of sweet potato (Ipomoea batatas) in different soils and climates. J. Adv. Microbiol. 2018, 13, 1–12. [Google Scholar]
- Vincent, J.M. A manual for practical study of root nodule bacteria. In IBP Handbook No. 15; Blackwell Scientific Publishers: Oxford, UK, 1970; p. 164. [Google Scholar]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar]
- Hiraishi, A.; Inagaki, K.; Tanimoto, Y.; Iwasaki, M.; Kishimoto, N.; Tanaka, I. Phylogenetic characterization of a new thermoacidophilic bacterium isolated from hot springs in Japan. J. Gen. Appl. Microbiol. 1997, 43, 295–304. [Google Scholar] [CrossRef] [Green Version]
- Freed, R. MSTAT: A software program for plant breeder. In Principles of Plant Genetics and Breeding, 2nd ed.; Acquaah, G., Ed.; Blackwell Publishing: Malden, MA, USA, 2007; Volume 1, pp. 426–431. [Google Scholar]
- Stoyanova, J. Growth, nodulation and nitrogen fixation in soybean as affected by air humidity and root temperature. Biol. Plant 1996, 38, 537–544. [Google Scholar] [CrossRef]
- Zhang, F.; Smith, D.L. Effects of low root zone temperatures on the early stages of symbiosis establishment between soybean [Glycine max (L.) Merr.] and Bradyrhizobium japonicum. J. Exp. Bot. 1994, 45, 1467–1473. [Google Scholar]
- Montañez, A.; Danso, S.K.A.; Hardarson, G. The effect of temperature on nodulation and nitrogen fixation by five Bradyrhizobium japonicum strains. Appl. Soil Ecol. 1995, 2, 165–174. [Google Scholar] [CrossRef]
- Hungria, M.; Vargas, M.A.T. Environmental factors affecting N2 fixation in grain legumes in the tropics, with an emphasis on Brazil. F. Crop Res. 2000, 65, 151–164. [Google Scholar] [CrossRef]
- Saeki, Y.; Aimi, N.; Hashimoto, M.; Tsukamoto, S.; Kaneko, A.; Yoshida, N.; Nagatomo, Y.; Akao, S. Grouping of bradyrhizobium usda strains by sequence analysis of 168 rRDNA and 168-238 rDNA internal transcribed spacer region. Soil Sci. Plant Nutr. 2004, 50, 517–525. [Google Scholar]
- Willems, A.; Munive, A.; de Lajudie, P.; Gillis, M. In most Bradyrhizobium groups sequence comparison of 16S-23S rDNA internal transcribed spacer regions corroborates DNA-DNA hybridizations. Syst. Appl. Microbiol. 2003, 26, 203–210. [Google Scholar] [CrossRef]
- Risal, C.P.; Yokoyama, T.; Ohkama-Ohtsu, N.; Djedidi, S.; Sekimoto, H. Genetic diversity of native soybean bradyrhizobia from different topographical regions along the southern slopes of the Himalayan Mountains in Nepal. Syst. Appl. Microbiol. 2010, 33, 416–425. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Latitude (°N) | Longitude (°E) | Temperature (°C) a | Rainfall (mm) a |
|---|---|---|---|---|
| Fukagawa | 43.71 | 142.01 | 16–26/16–26 (14–24/17–27) b | 432/243 |
| Matsue | 35.48 | 133.06 | 23–30/23–30 (22–29/25–32) | 177/481 |
| Miyazaki | 31.82 | 131.41 | 24–32/24–31 (25–32/25–33) | 240/860 |
| Closest 16S rDNA | ITS Group | Closest ITS a | Acc. No. b | Id. (%) | Remarks |
|---|---|---|---|---|---|
| B. japonicum Bj11 | Bj11-1 | Bj CCBAU 45394 | HM107195 | 98–99 | Slow grower Primarily Fukagawa soil |
| Bj11-2 | Bj CCBAU 45394 | HM107195 | 99–100 | Fast grower Fukagawa and Matsue soils | |
| B. japonicum S10J | BjS10J-1 | Bj ITS04K-a | KY564319 | 99–100 | Matsue soil |
| BjS10J-2 | Bj USDA 6 | HQ143390 | 98–99 | Miyazaki soil | |
| BjS10J-3 | Bj 367 | KU298516 | 96–100 | Matsue soil | |
| BjS10J-4 | Bj USDA 38 | AB100743 | 99–100 | Primarily Miyazaki soil | |
| B.elkanii L7 | BeL7 | Be USDA 29 | EU834730 | 97–100 | Ubiquitous in all soils |
| Soil/ Location | Year | Bj11 | BjS10J | BeL7 | ||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 1 | 2 | 3 | 4 | |||
| FU/fu | 2016 | 87 | 13 | - | - | - | - | - |
| 2017 | 80 | 17 | - | - | - | - | 3 | |
| FU/ma | 2016 | 80 | 17 | - | - | - | - | 3 |
| 2017 | 53 | 7 | - | - | - | - | 40 | |
| FU/mi | 2016 | 83 | 7 | - | - | - | - | 10 |
| 2017 | 60 | 10 | - | - | - | - | 30 | |
| MA/fu | 2016 | 17 | 7 | 7 | - | 13 | 3 | 53 |
| 2017 | - | 23 | - | - | - | - | 77 | |
| MA/ma | 2016 | - | 20 | 3 | - | - | 7 | 70 |
| 2017 | 3 | 7 | - | - | 3 | - | 87 | |
| MA/mi | 2016 | 7 | 17 | - | - | 3 | - | 73 |
| 2017 | - | - | - | - | 3 | - | 97 | |
| MI/fu | 2016 | - | - | - | 73 | - | 13 | 13 |
| 2017 | - | - | - | 13 | - | 7 | 80 | |
| MI/ma | 2016 | - | - | - | 53 | - | 13 | 33 |
| 2017 | - | - | - | - | - | - | 100 | |
| MI/mi | 2016 | - | - | - | 7 | - | 17 | 77 |
| 2017 | - | - | - | - | - | - | 100 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hafiz, M.H.R.; Salehin, A.; Adachi, F.; Omichi, M.; Saeki, Y.; Yamamoto, A.; Hayashi, S.; Itoh, K. Latitudinal Characteristic Nodule Composition of Soybean-Nodulating Bradyrhizobia: Temperature-Dependent Proliferation in Soil or Infection? Horticulturae 2021, 7, 22. https://doi.org/10.3390/horticulturae7020022
Hafiz MHR, Salehin A, Adachi F, Omichi M, Saeki Y, Yamamoto A, Hayashi S, Itoh K. Latitudinal Characteristic Nodule Composition of Soybean-Nodulating Bradyrhizobia: Temperature-Dependent Proliferation in Soil or Infection? Horticulturae. 2021; 7(2):22. https://doi.org/10.3390/horticulturae7020022
Chicago/Turabian StyleHafiz, Md Hafizur Rahman, Ahsanul Salehin, Fumihiko Adachi, Masayuki Omichi, Yuichi Saeki, Akihiro Yamamoto, Shohei Hayashi, and Kazuhito Itoh. 2021. "Latitudinal Characteristic Nodule Composition of Soybean-Nodulating Bradyrhizobia: Temperature-Dependent Proliferation in Soil or Infection?" Horticulturae 7, no. 2: 22. https://doi.org/10.3390/horticulturae7020022