Alleviation of Heat Stress in Tomato by Exogenous Application of Sulfur

 ,
,  ,
,  ,
,  ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Conditions
2.2. Treatments
2.3. Morphological Variables
2.4. Physiological Variables
2.5. Biochemical Variables
2.6. Statistical Analysis
3. Results
3.1. Roma (Thermotolerant)
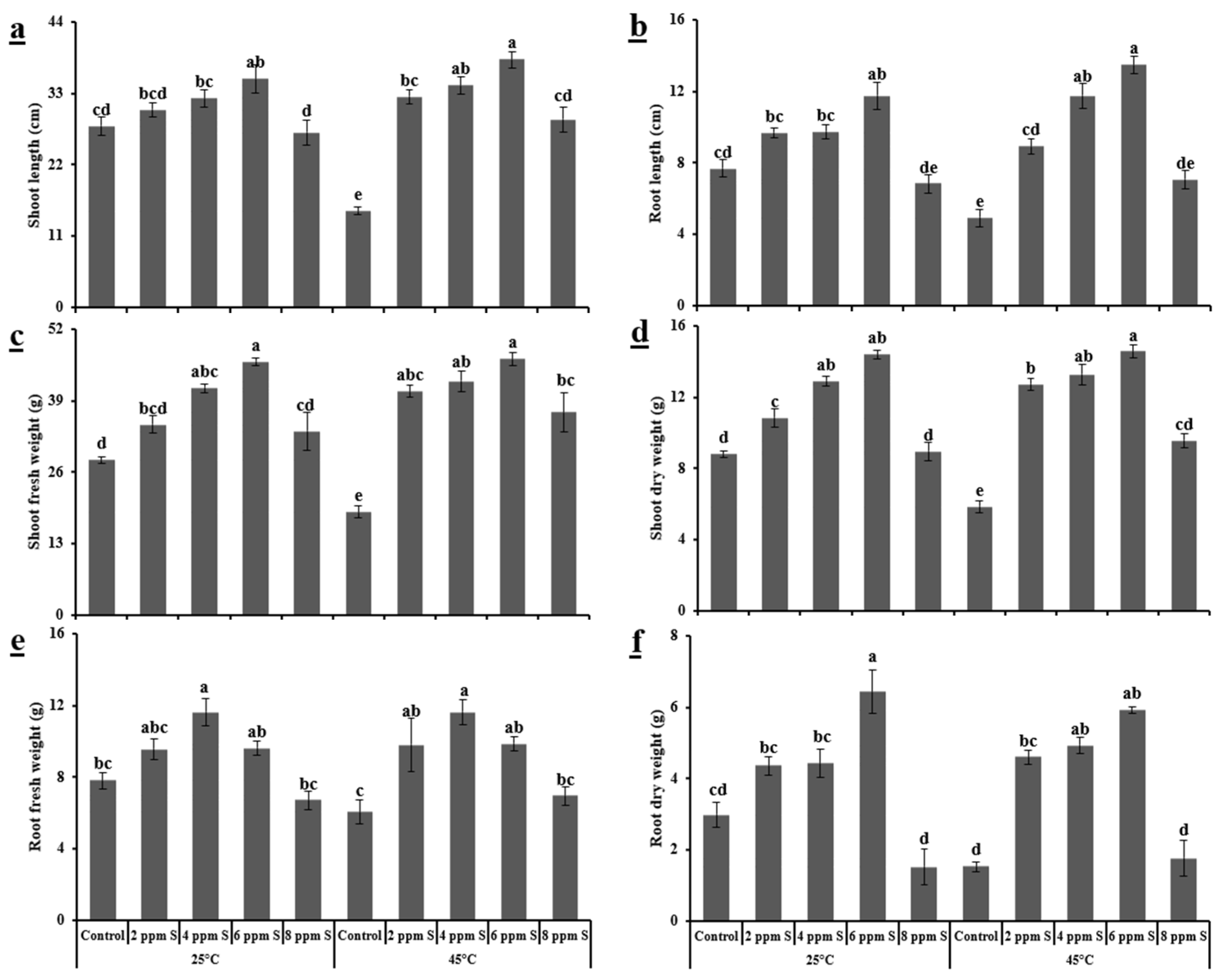
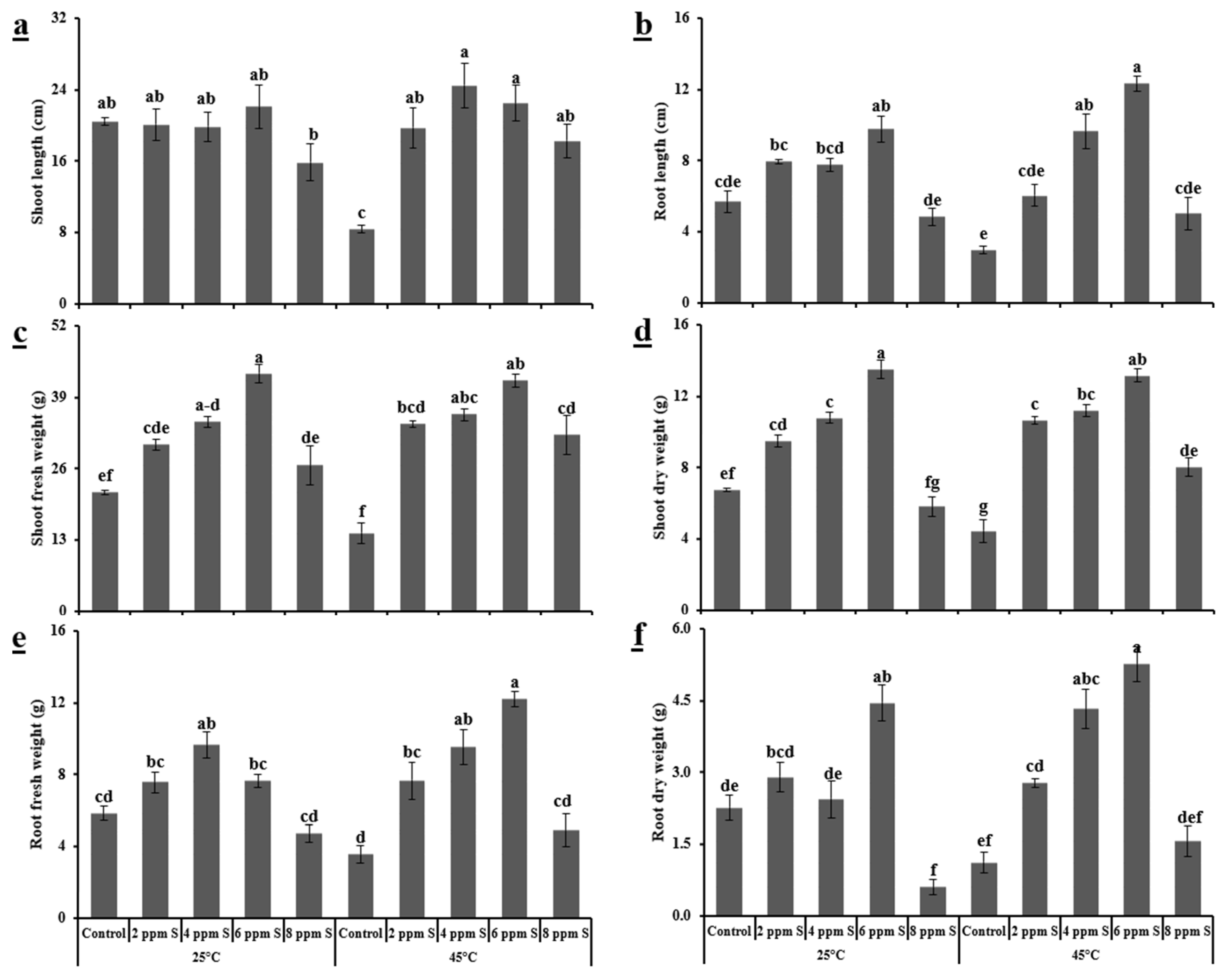
3.1.1. Morphological Variables
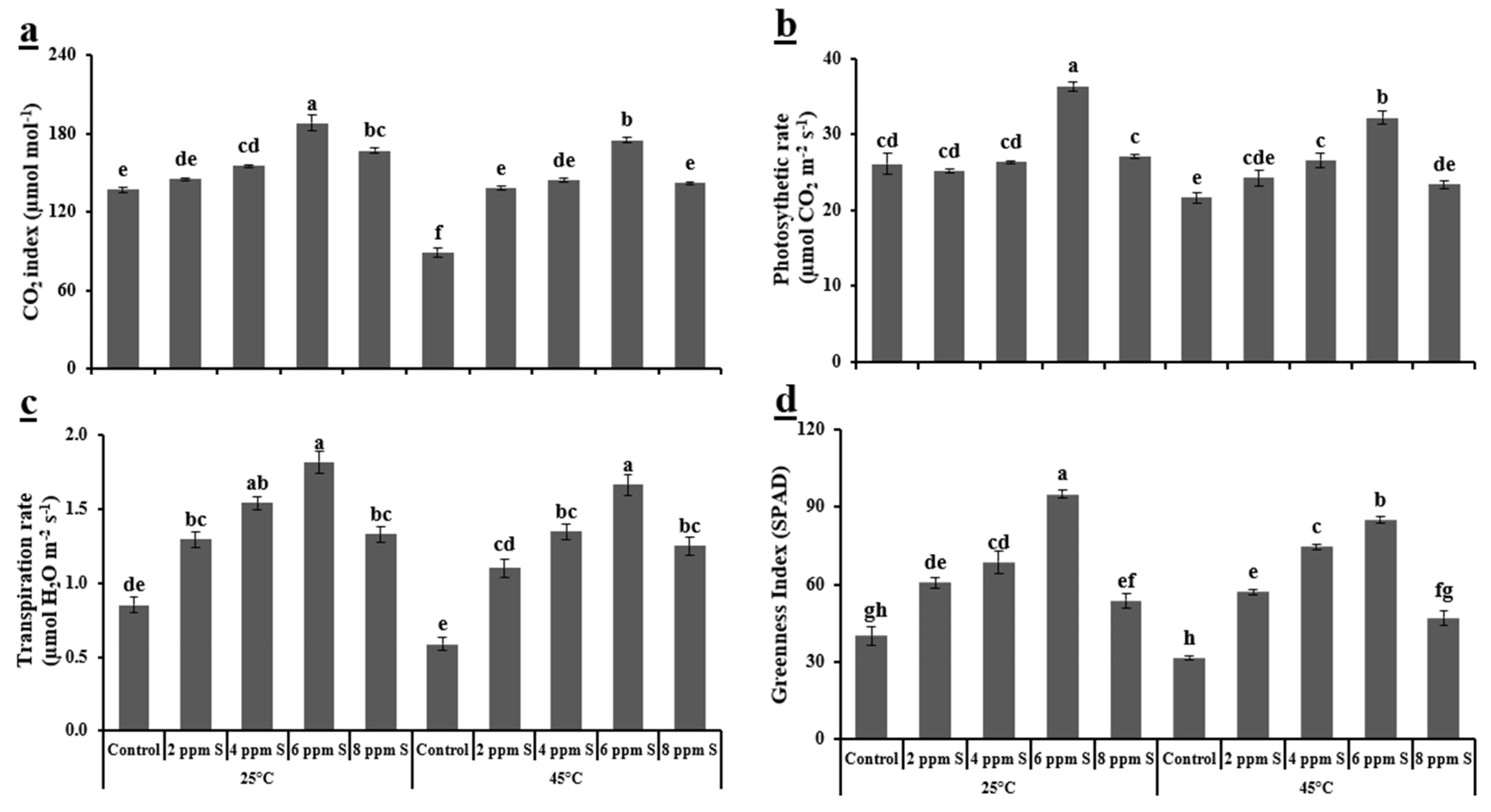
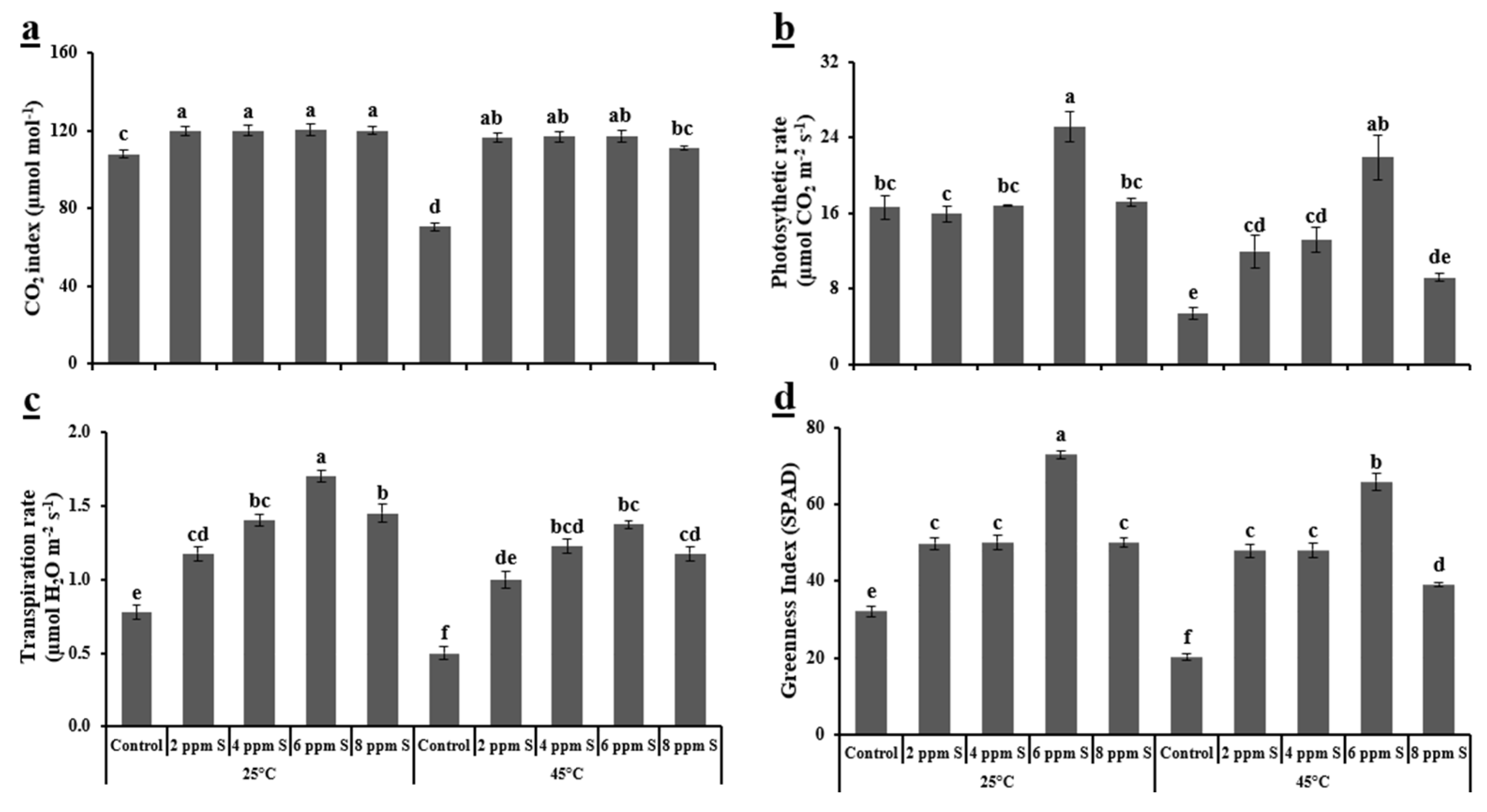
3.1.2. Physiological Variables
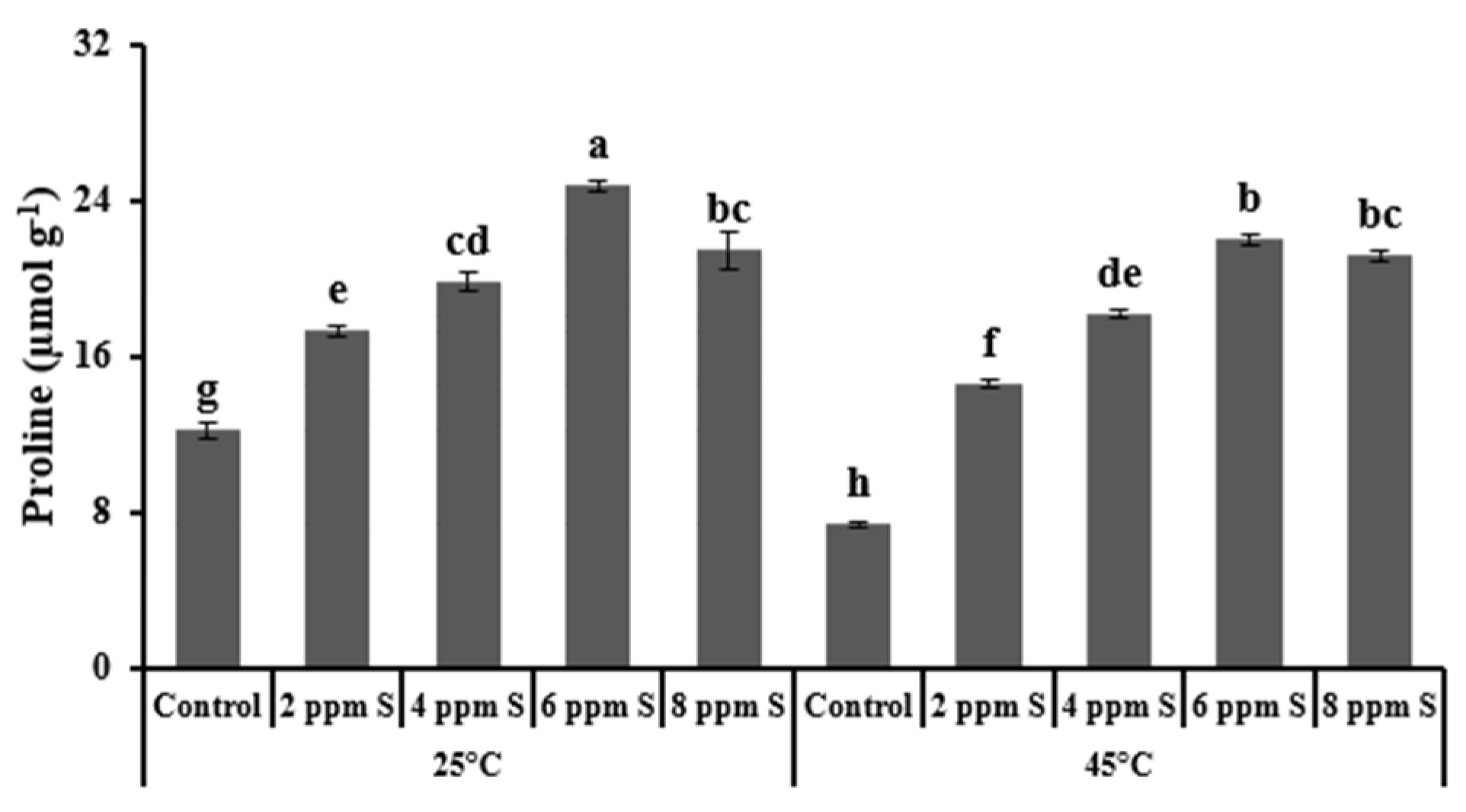
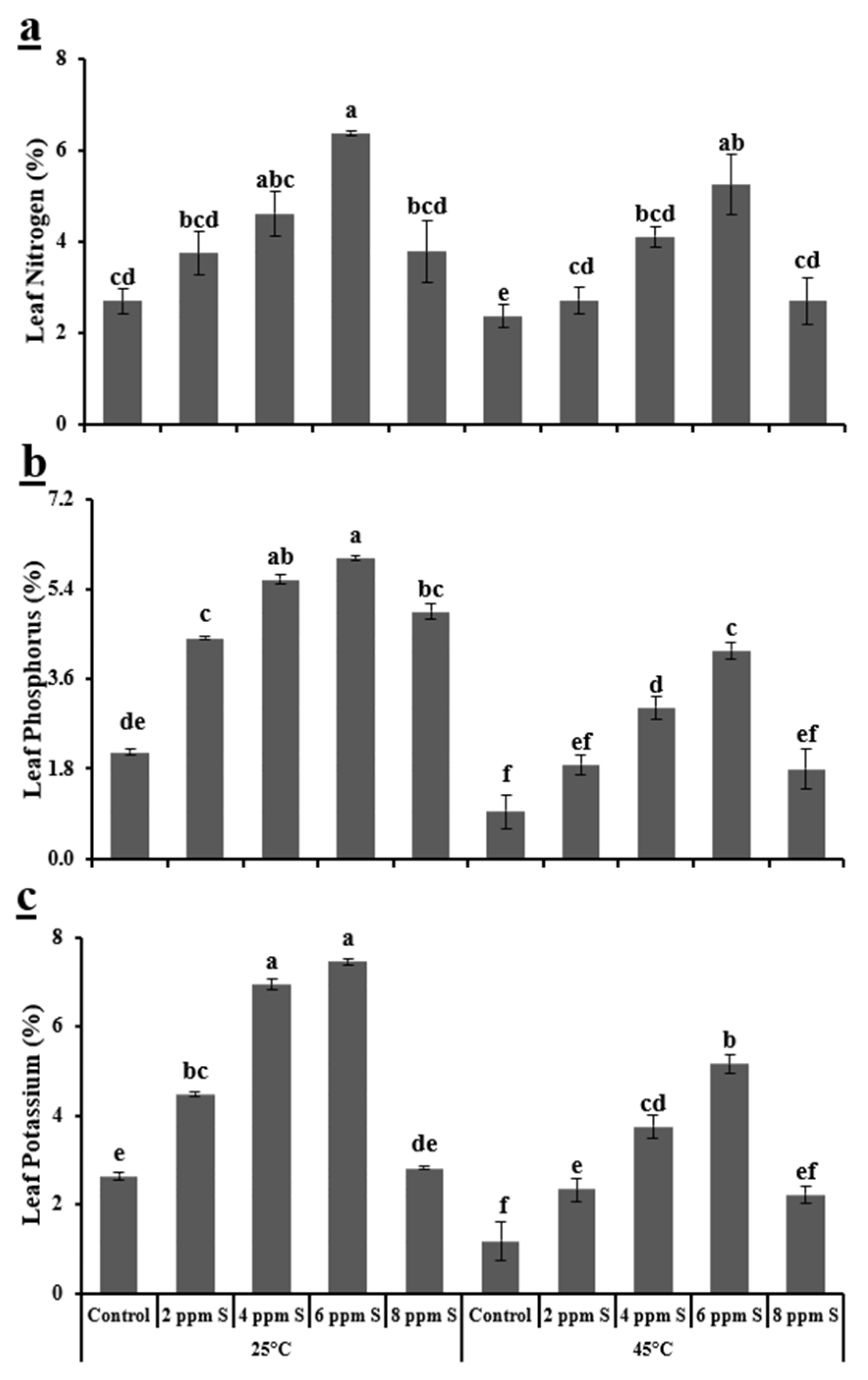
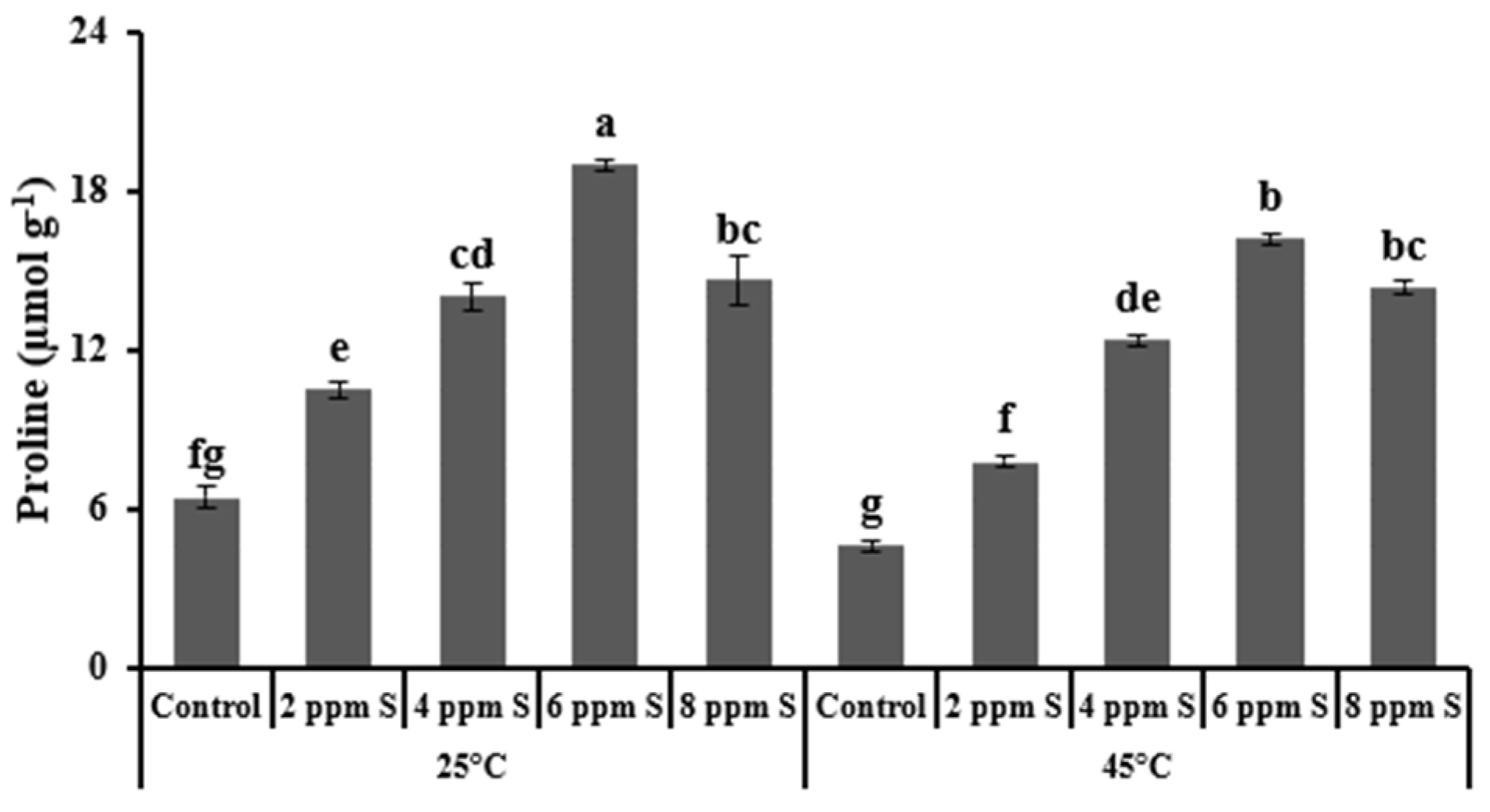
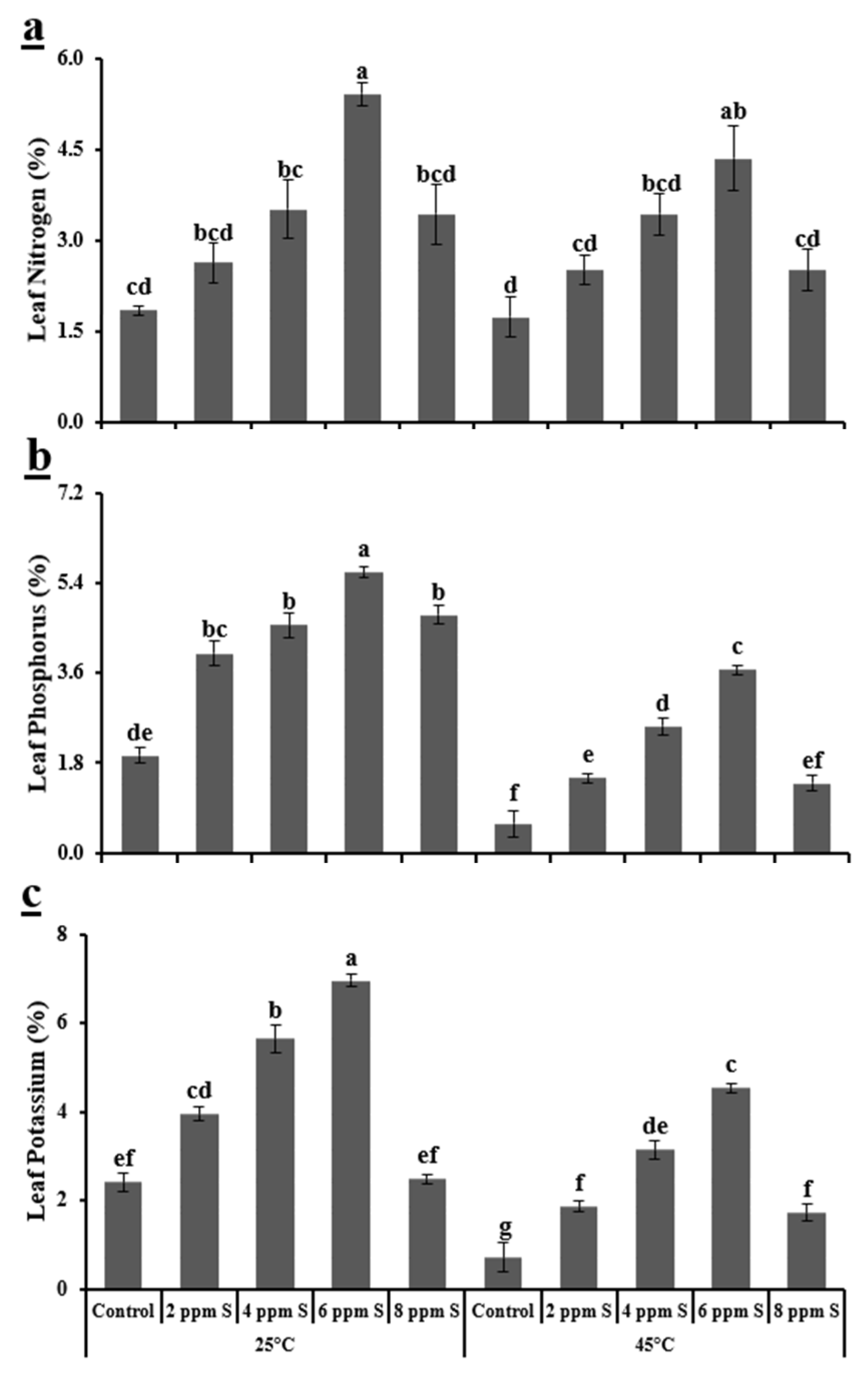
3.1.3. Biochemical Variables
3.2. Ahmar (Thermotolerant)
3.2.1. Morphological Variables
3.2.2. Physiological Variables
3.2.3. Biochemical Variables
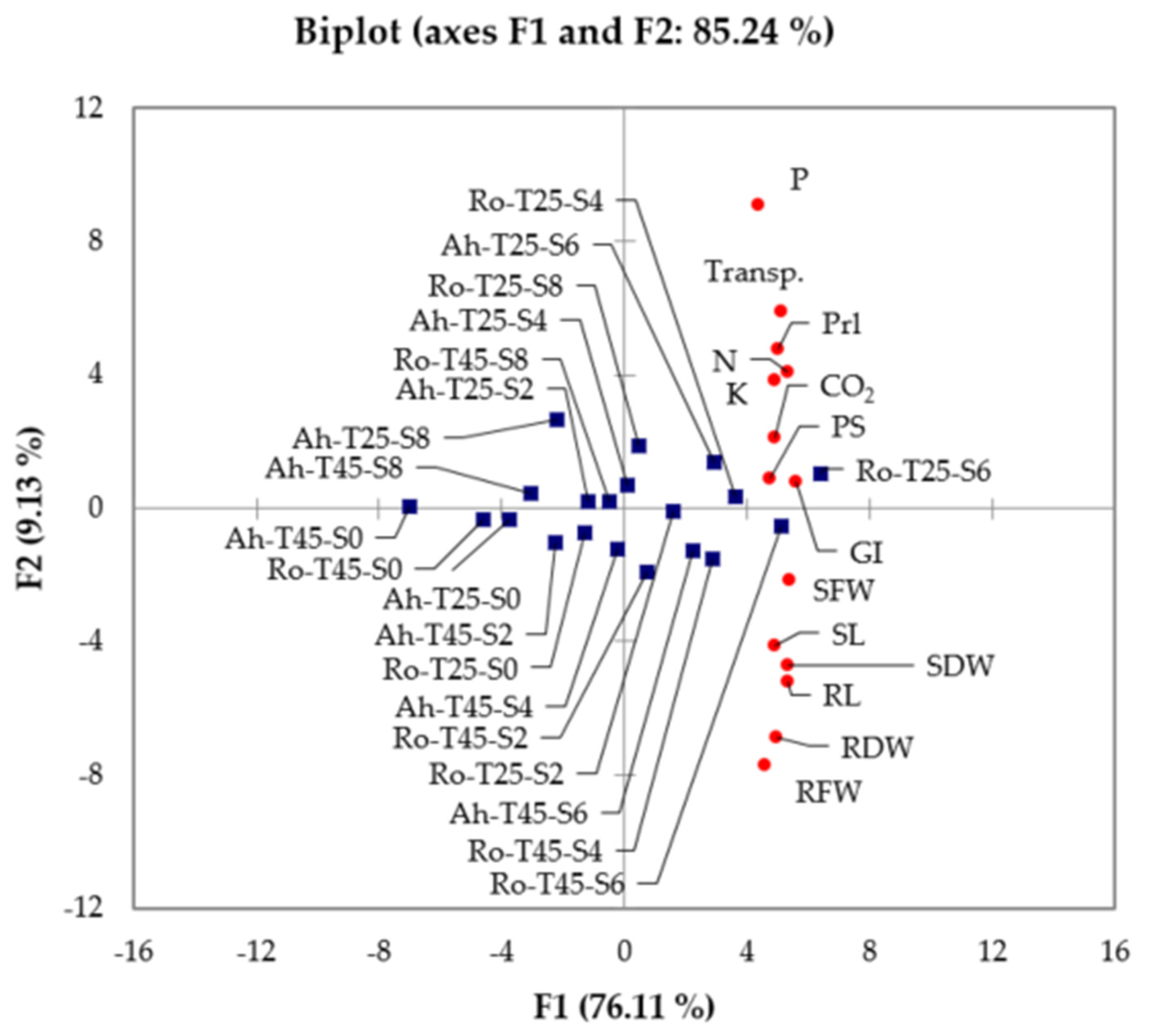
3.3. Principle Component Analysis
4. Discussion
4.1. Roma (Thermotolerant)
4.2. Ahmar (Thermosensitive)
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bai, Y.; Lindhout, P. Domestication and Breeding of Tomatoes: What have We Gained and What Can We Gain in the Future? Ann. Bot. 2007, 100, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Moaaz Ali, M.; Javed, T.; Mauro, R.P.; Shabbir, R.; Afzal, I.; Yousef, A.F. Effect of Seed Priming with Potassium Nitrate on the Performance of Tomato. Agriculture 2020, 10, 498. [Google Scholar] [CrossRef]
- FAOSTAT. World Food and Agriculture 2018: Statistical Pocketbook; FAO: Rome, Italy, 2018; ISBN 9789251310120. [Google Scholar]
- Mauro, R.P.; Agnello, M.; Distefano, M.; Sabatino, L.; San Bautista Primo, A.; Leonardi, C.; Giuffrida, F. Chlorophyll Fluorescence, Photosynthesis and Growth of Tomato Plants as Affected by Long-Term Oxygen Root Zone Deprivation and Grafting. Agronomy 2020, 10, 137. [Google Scholar] [CrossRef] [Green Version]
- Golam, F.; Hossain Prodhan, Z.; Nezhadahmadi, A.; Rahman, M. Heat tolerance in tomato. Life Sci. J. 2012, 9, 1936–1950. [Google Scholar]
- Peet, M.M.; Willits, D.H.; Gardner, R. Response of ovule development and post-pollen production processes in male-sterile tomatoes to chronic, sub-acute high temperature stress. J. Exp. Bot. 1997, 48, 101–111. [Google Scholar] [CrossRef]
- Firon, N.; Pressman, E.; Meir, S.; Khoury, R.; Altahan, L. Ethylene is involved in maintaining tomato (Solanum lycopersicum) pollen quality under heat-stress conditions. AoB Plants 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Baki, A.A. Tolerance of Tomato Cultivars and Selected Germplasm to Heat Stress. J. Am. Soc. Hortic. Sci. 1991, 116, 1113–1116. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.; Roychowdhury, R.; Fujita, M. Physiological, Biochemical, and Molecular Mechanisms of Heat Stress Tolerance in Plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef]
- Wise, R.R.; Olson, A.J.; Schrader, S.M.; Sharkey, T.D. Electron transport is the functional limitation of photosynthesis in field-grown Pima cotton plants at high temperature. Plant Cell Environ. 2004, 27, 717–724. [Google Scholar] [CrossRef]
- Ribeiro, R.V.; Machado, E.C.; Oliveira, R.F. Temperature response of photosynthesis and its interaction with light intensity in sweet orange leaf discs under non-photorespiratory condition. Ciênc. e Agrotecnol. 2006, 30, 670–678. [Google Scholar] [CrossRef]
- Rout, G.R.; Das, A.B. (Eds.) Molecular Stress Physiology of Plants; Springer: New Delhi, India, 2013; ISBN 978-81-322-0806-8. [Google Scholar]
- Ahanger, M.A.; Akram, N.A.; Ashraf, M.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P. Plant responses to environmental stresses—from gene to biotechnology. AoB Plants 2017, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Kaur, R.; Kaur, N.; Bhandhari, K.; Kaushal, N.; Gupta, K.; Bains, T.S.; Nayyar, H. Heat-stress induced inhibition in growth and chlorosis in mungbean (Phaseolus aureus Roxb.) is partly mitigated by ascorbic acid application and is related to reduction in oxidative stress. Acta Physiol. Plant. 2011, 33, 2091–2101. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.S.; Bhuyan, M.H.M.B.; Al Mahmud, J.; Nahar, K.; Fujita, M. The Role of Sulfur in Plant Abiotic Stress Tolerance: Molecular Interactions and Defense Mechanisms. In Plant Nutrients and Abiotic Stress Tolerance; Springer: Singapore, 2018; pp. 221–252. ISBN 9789811090448. [Google Scholar]
- Skudra, I.; Ruza, A. Effect of Nitrogen and Sulphur Fertilization on Chlorophyll Content in Winter Wheat. Rural Sustain. Res. 2017, 37, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Mahmud, J.A.; Nahar, K.; Mohsin, S.M.; Parvin, K.; Fujita, M. Interaction of sulfur with phytohormones and signaling molecules in conferring abiotic stress tolerance to plants. Plant Signal. Behav. 2018, 13, e1477905. [Google Scholar] [CrossRef] [PubMed]
- Bashir, H.; Ibrahim, M.M.; Bagheri, R.; Ahmad, J.; Arif, I.A.; Baig, M.A.; Qureshi, M.I. Influence of sulfur and cadmium on antioxidants, phytochelatins and growth in Indian mustard. AoB Plants 2015, 7, plv001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehar, F.; Masood, A.; Khan, N. Coordinate changes in assimilatory sulfate reduction are correlated to salt tolerance: Involvement of phytohormones. Annu. Rev. Res. Biol. 2013, 3, 267–295. [Google Scholar]
- Biswal, B.; Raval, M.K.; Biswal, U.C.; Joshi, P. Response of Photosynthetic Organelles to Abiotic Stress: Modulation by Sulfur Metabolism. In Sulfur Assimilation and Abiotic Stress in Plants; Springer: Berlin/Heidelberg, Germany, 2008; pp. 167–191. [Google Scholar]
- Ihsan, M.Z.; Daur, I.; Alghabari, F.; Alzamanan, S.; Rizwan, S.; Ahmad, M.; Waqas, M.; Shafqat, W. Heat stress and plant development: Role of sulphur metabolites and management strategies. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2019, 69, 332–342. [Google Scholar] [CrossRef]
- Khokhar, K.M.; HRI, N. Present status and prospects of tomatoes in Pakistan. Agric. Corner-Farmers Glob. Mark. Rep. 2013. [Google Scholar] [CrossRef]
- Alsamir, M.; Ahmad, N.; Arief, V.; Mahmood, T.; Trethowan, R. Phenotypic diversity and marker-trait association studies under heat stress in tomato (Solanum lycopersicum L.). Aust. J. Crop Sci. 2019, 13, 578–587. [Google Scholar] [CrossRef]
- Alsamir, M. Genetic and Physiological Analysis of Tomato (Solanum lycopersicum L.) Adaption under Heat and Disease Stress. Ph.D. Thesis, School of Life and Environmental Sciences, The University of Sydney, Sydney, Australia, 2019. [Google Scholar]
- Sorgonà, A.; Abenavoli, M.R.; Cacco, G.; Gelsomino, A. Growth of Tomato and Zucchini Seedlings in Orange Waste Compost Media: pH and Implication of Dosage. Compost Sci. Util. 2011, 19, 189–196. [Google Scholar] [CrossRef]
- Estefan, G.; Sommer, R.; Ryan, J. Methods of Soil, Plant, and Water Analysis: A manual for the West Asia and North. Int. Cent. Agric. Res. Dry Areas 2013. Available online: https://hdl.handle.net/20.500.11766/7512 (accessed on 20 January 2021).
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Wang, C.; Jing, R.; Mao, X.; Chang, X.; Li, A. TaABC1, a member of the activity of bc1 complex protein kinase family from common wheat, confers enhanced tolerance to abiotic stresses in Arabidopsis. J. Exp. Bot. 2011, 62, 1299–1311. [Google Scholar] [CrossRef] [PubMed]
- Orman, S.; Kaplan, M. Effects of elemental sulphur and farmyard manure on pH and salinity of calcareous sandy loam soil and some nutrient elements in tomato plant. J. Agric. Sci. Technol. 2011, 5, 20–26. [Google Scholar]
- Ayenan, M.A.T.; Danquah, A.; Hanson, P.; Ampomah-Dwamena, C.; Sodedji, F.A.K.; Asante, I.K.; Danquah, E.Y. Accelerating Breeding for Heat Tolerance in Tomato (Solanum lycopersicum L.): An Integrated Approach. Agronomy 2019, 9, 720. [Google Scholar] [CrossRef] [Green Version]
- Bukhov, N.G.; Wiese, C.; Neimanis, S.; Heber, U. Heat sensitivity of chloroplasts and leaves: Leakage of protons from thylakoids and reversible activation of cyclic electron transport. Photosynth. Res. 1999, 59, 81–93. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Machado, S.; Paulsen, G.M. Combined effects of drought and high temperature on water relations of wheat and sorghum. Plant Soil 2001, 233, 179–187. [Google Scholar] [CrossRef]
- Huve, K.; Bichele, I.; Tobias, M.; Niinemets, U. Heat sensitivity of photosynthetic electron transport varies during the day due to changes in sugars and osmotic potential. Plant Cell Environ. 2006, 29, 212–228. [Google Scholar] [CrossRef]
- Gururani, M.; Mohanta, T.; Bae, H. Current Understanding of the Interplay between Phytohormones and Photosynthesis under Environmental Stress. Int. J. Mol. Sci. 2015, 16, 19055–19085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camejo, D.; Rodríguez, P.; Angeles Morales, M.; Miguel Dell’Amico, J.; Torrecillas, A.; Alarcón, J.J. High temperature effects on photosynthetic activity of two tomato cultivars with different heat susceptibility. J. Plant Physiol. 2005, 162, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Warman, P.R.; Sampson, H.G. Effect of sulfur additions on the yield and elemental composition of canola and spring wheat 1. J. Plant Nutr. 1994, 17, 1817–1825. [Google Scholar] [CrossRef]
- Giaveno, C.; Ferrero, J. Introduction of tropical maize genotypes to increase silage production in the central area of Santa Fe, Argentina. Crop. Breed. Appl. Biotechnol. 2003, 3, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Hlaváčová, M.; Klem, K.; Rapantová, B.; Novotná, K.; Urban, O.; Hlavinka, P.; Smutná, P.; Horáková, V.; Škarpa, P.; Pohanková, E.; et al. Interactive effects of high temperature and drought stress during stem elongation, anthesis and early grain filling on the yield formation and photosynthesis of winter wheat. Field Crop. Res. 2018, 221, 182–195. [Google Scholar] [CrossRef]
- Moreira, A.; de Carvalho, J.G.; Evangelista, A.R. Effect of sulfur doses on the production and mineral composition of alfalfa. Pesqui. Agropecuária Bras. 1997, 32, 533–538. [Google Scholar]
- De Souza Silva, M.L.; Trevizam, A.R.; de Cássia Piccolo, M.; Furlan, G. Tomato production in function of sulfur doses application. Rev. Bras. Tecnol. Apl. Nas Ciênc. Agrár. 2014, 7. [Google Scholar] [CrossRef]
- Zhang, J.-H.; Huang, W.-D.; Liu, Y.-P.; Pan, Q.-H. Effects of Temperature Acclimation Pretreatment on the Ultrastructure of Mesophyll Cells in Young Grape Plants (Vitis vinifera L. cv. Jingxiu) Under Cross-Temperature Stresses. J. Integr. Plant Biol. 2005, 47, 959–970. [Google Scholar] [CrossRef]
- Vollenweider, P.; Günthardt-Goerg, M.S. Diagnosis of abiotic and biotic stress factors using the visible symptoms in foliage. Environ. Pollut. 2005, 137, 455–465. [Google Scholar] [CrossRef]
- Bita, C.E.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Ruban, A.V. Nonphotochemical Chlorophyll Fluorescence Quenching: Mechanism and Effectiveness in Protecting Plants from Photodamage. Plant Physiol. 2016, 170, 1903–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherer, H.W.; Lange, A. N2 fixation and growth of legumes as affected by sulphur fertilization. Biol. Fertil. Soils 1996, 23, 449–453. [Google Scholar] [CrossRef]
- Giri, A.; Heckathorn, S.; Mishra, S.; Krause, C. Heat Stress Decreases Levels of Nutrient-Uptake and -Assimilation Proteins in Tomato Roots. Plants 2017, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Habtegebrial, K.; Singh, B.R. Effects of timing of nitrogen and sulphur fertilizers on yield, nitrogen, and sulphur contents of Tef (Eragrostis tef (Zucc.) Trotter). Nutr. Cycl. Agroecosyst. 2006, 75, 213–222. [Google Scholar] [CrossRef]
- Carciochi, W.D.; Divito, G.A.; Fernández, L.A.; Echeverría, H.E. Sulfur affects root growth and improves nitrogen recovery and internal efficiency in wheat. J. Plant Nutr. 2017, 40, 1231–1242. [Google Scholar] [CrossRef]
- Salvagiotti, F.; Castellarín, J.M.; Miralles, D.J.; Pedrol, H.M. Sulfur fertilization improves nitrogen use efficiency in wheat by increasing nitrogen uptake. Field Crop. Res. 2009, 113, 170–177. [Google Scholar] [CrossRef]
- Pużyńska, K.; Kulig, B.; Halecki, W.; Lepiarczyk, A.; Pużyński, S. Response of oilseed rape leaves to sulfur and boron foliar application. Acta Physiol. Plant. 2018, 40, 169. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, M.M.; Waleed Shafique, M.; Gull, S.; Afzal Naveed, W.; Javed, T.; Yousef, A.F.; Mauro, R.P. Alleviation of Heat Stress in Tomato by Exogenous Application of Sulfur. Horticulturae 2021, 7, 21. https://doi.org/10.3390/horticulturae7020021
Ali MM, Waleed Shafique M, Gull S, Afzal Naveed W, Javed T, Yousef AF, Mauro RP. Alleviation of Heat Stress in Tomato by Exogenous Application of Sulfur. Horticulturae. 2021; 7(2):21. https://doi.org/10.3390/horticulturae7020021
Chicago/Turabian StyleAli, Muhammad Moaaz, Muhammad Waleed Shafique, Shaista Gull, Waleed Afzal Naveed, Talha Javed, Ahmed Fathy Yousef, and Rosario Paolo Mauro. 2021. "Alleviation of Heat Stress in Tomato by Exogenous Application of Sulfur" Horticulturae 7, no. 2: 21. https://doi.org/10.3390/horticulturae7020021