
Roles of Si and SiNPs in Improving Thermotolerance of Wheat Photosynthetic Machinery via Upregulation of PsbH, PsbB and PsbD Genes Encoding PSII Core Proteins

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Growth Conditions
2.2. Imposition of Treatments
2.3. Methods
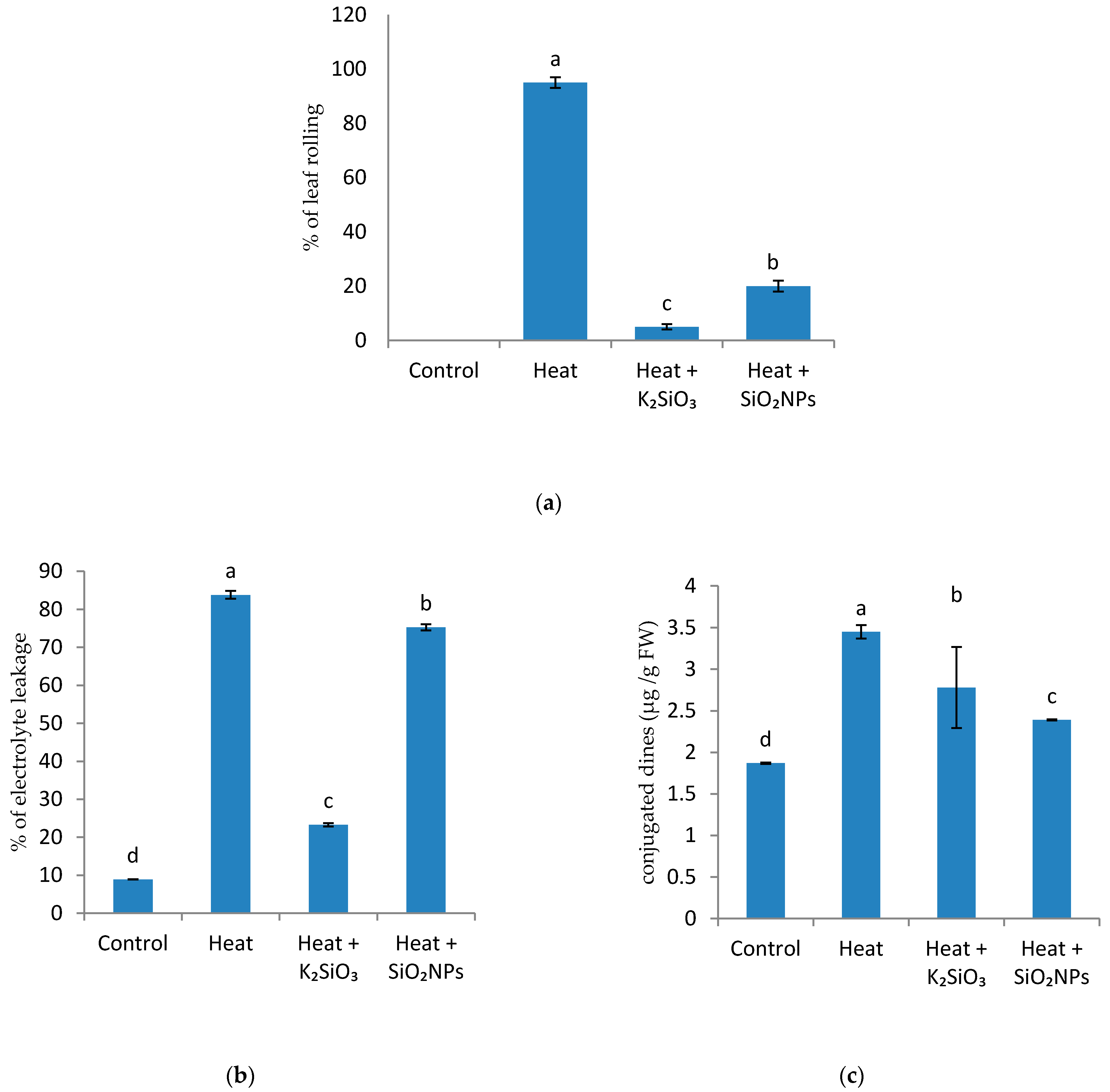
2.3.1. Leaf Rolling Score
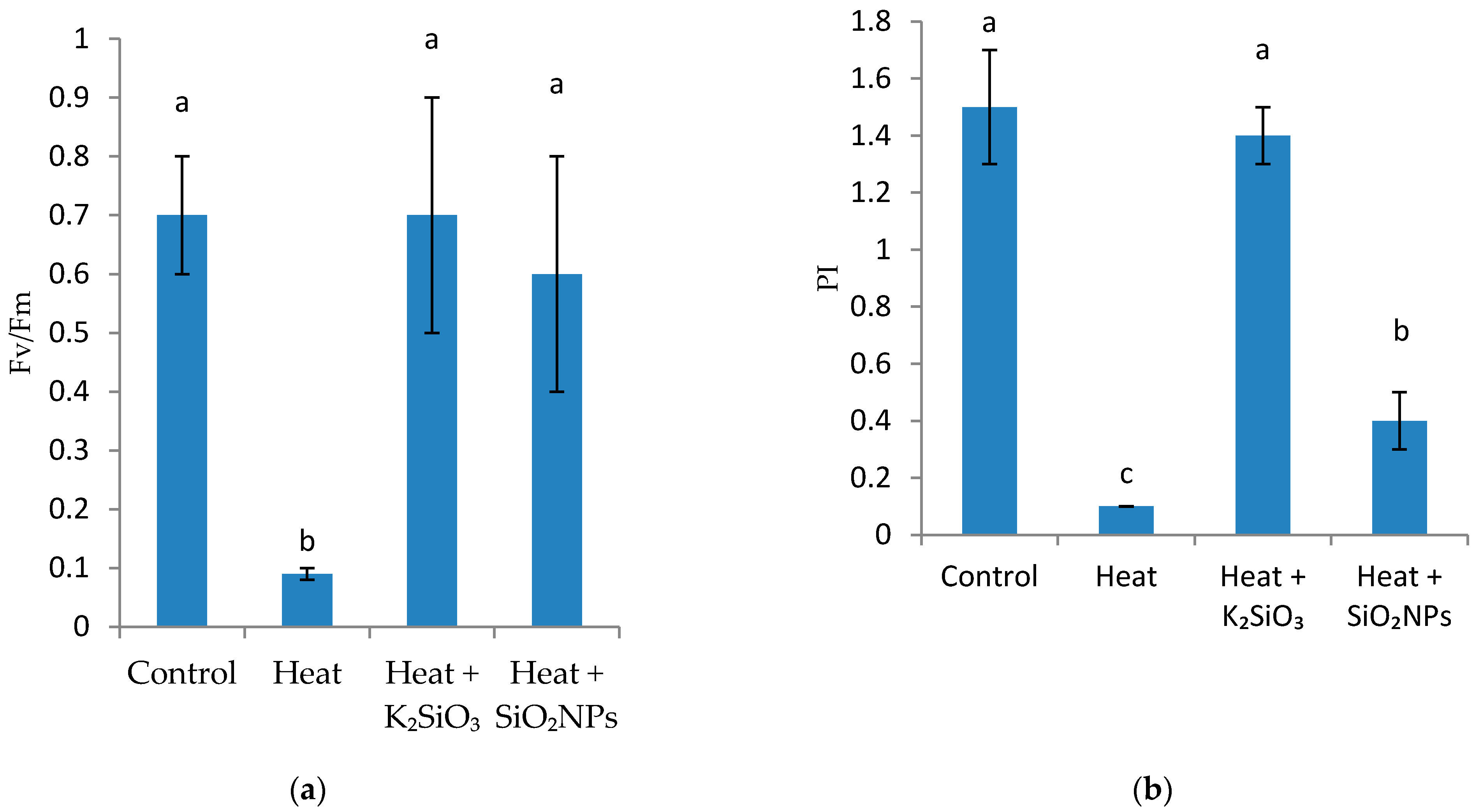
2.3.2. Chlorophyll Fluorescence Measurements
2.3.3. Estimation of Photosynthetic Pigments
2.3.4. Estimation of Carbohydrates
2.3.5. Determination of Proline
2.3.6. Determination of Electrolyte Leakage (EL)
2.3.7. Estimation of Lipid Peroxidation Products (Conjugated Dienes, CD)
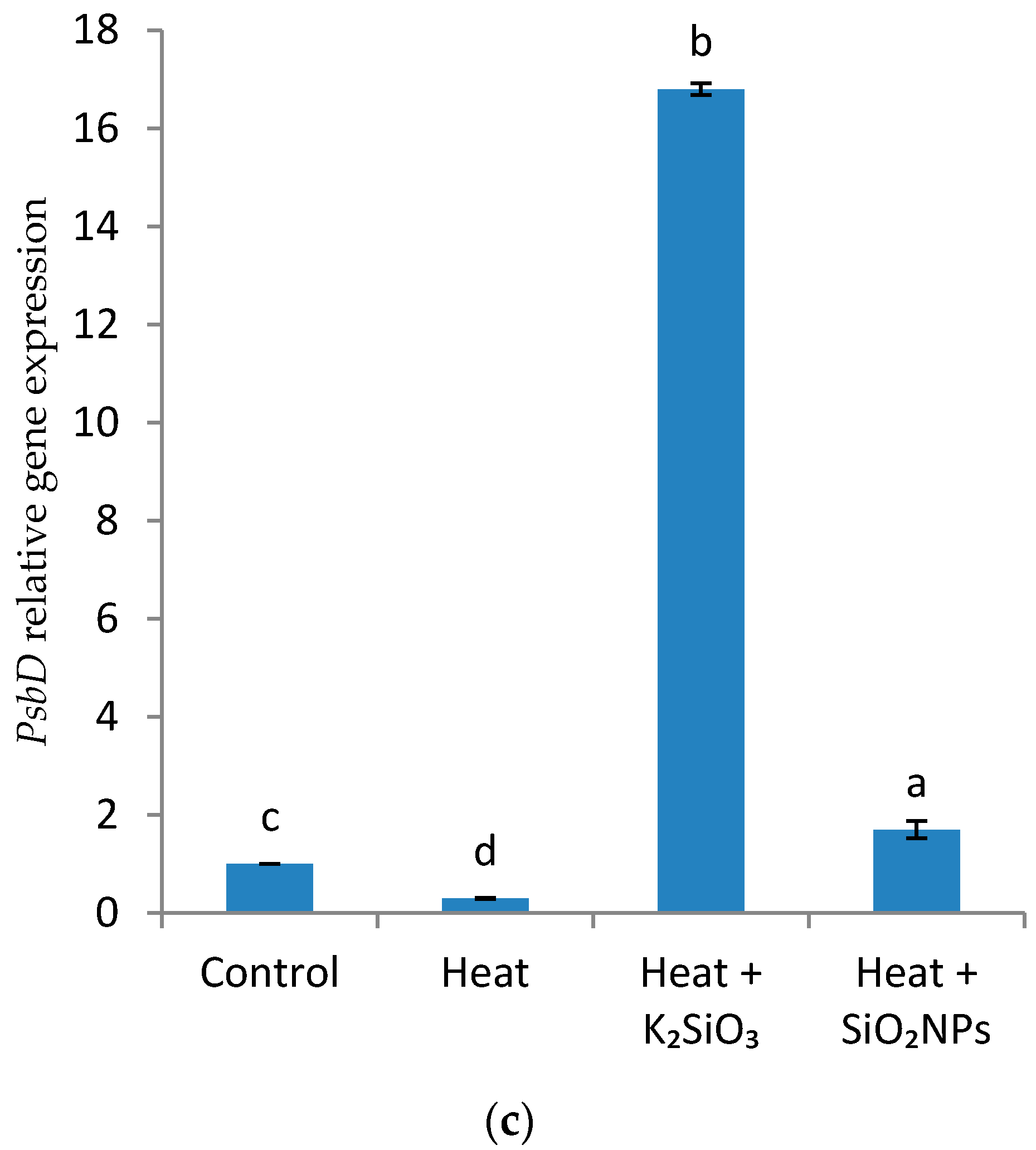
2.3.8. Quantitative Real-Time PCR (qRT-PCR) Analysis
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Chl a | chlorophyll a |
| Chl b | chlorophyll b |
| EL | electrolyte leakage |
| Fv/Fm | maximum quantum yield of photosystem II |
| PIabs | performance index |
| PsbH, PsbB, and PsbD | photosystem II reaction center protein H, B, and D |
| PSII | photosystem II |
| Si | silicon |
| SiNPs | silicon nanoparticles |
| WHC | water holding capacity |
References
- Tan, W.; Meng, Q.W.; Brestic, M.; Olsovska, K.; Yang, X. Photosynthesis is improved by exogenous calcium in heat-stressed tobacco plants. J. Plant Physiol. 2011, 168, 2063–2071. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.L.; Chen, J.H.; He, N.Y.; Guo, F.Q. Metabolic reprogramming in chloroplasts under heat stress in plants. Int. J. Mol. Sci. 2018, 19, 849. [Google Scholar] [CrossRef] [PubMed]
- Paul, P.; Mesihovic, A.; Chaturvedi, P.; Ghatak, A.; Weckwerth, W.; Böhmer, M.; Schleiff, E. Structural and Functional Heat Stress Responses of Chloroplasts of Arabidopsis thaliana. Genes 2020, 11, 650. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, T.D. Effects of moderate heat stress on photosynthesis: Importance of thylakoid reactions, rubisco deactivation, reactive oxygen species, and thermotolerance provided by isoprene. Plant Cell Environ. 2005, 28, 269–277. [Google Scholar] [CrossRef]
- Taylor, N.L.; Tan, Y.F.; Jacoby, R.P.; Millar, A.H. Abiotic environmental stress induced changes in the Arabidopsis thaliana chloroplast, mitochondria and peroxisome proteomes. J. Proteom. 2009, 72, 367–378. [Google Scholar] [CrossRef]
- Allakhverdiev, S.I.; Kreslavski, V.D.; Klimov, V.V.; Los, D.A.; Carpentier, R.; Mohanty, P. Heat stress: An overview of molecular responses in photosynthesis. Photosynth. Res. 2008, 98, 541. [Google Scholar] [CrossRef]
- Law, R.D.; Crafts-Brandner, S.J. Inhibition and acclimation of photosynthesis to heat stress is closely correlated with activation of ribulose-1, 5-bisphosphate carboxylase/oxygenase. Plant Physiol. 1999, 120, 173–182. [Google Scholar] [CrossRef]
- Nelson, N.; Junge, W. Structure and energy transfer in photosystems of oxygenic photosynthesis Effect of silicon on drought tolerance of upland rice. Annu. Rev. Biochem. 2015, 84, 659–683. [Google Scholar] [CrossRef]
- Lu, C.M.; Zhang, J.H. Heat-Induced multiple effects on PSII in wheat plants. J. Plant Physiol. 2000, 156, 259–265. [Google Scholar] [CrossRef]
- Dankov, K.; Rashkov, G.; Misra, A.; Apostolova, E.L. Temperature sensitivity of photosystem II in isolated thylakoid membranes from fluridone-treated pea leaves. Turk. J. Bot. 2015, 39, 420–428. [Google Scholar] [CrossRef]
- Li, H.; Xu, H.; Zhang, P.; Gao, M.; Wang, D.; Zhao, H. High temperature effects on D1 protein turnover in three wheat varieties with different heat susceptibility. Plant Growth Regul. 2017, 81, 1–9. [Google Scholar] [CrossRef]
- Li, L.; Yang, H.; Liu, P.; Ren, W.; Wu, X.; Huang, F. Combined impact of heat stress and phosphate deficiency on growth and photochemical activity of sheep grass (Leymus chinensis). J. Plant Physiol. 2018, 231, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, E.I.; Fischer, G.; Van Velthuizen, H.; Walter, C.; Ewert, F. Global hot-spots of heat stress on agricultural crops due to climate change. Agric. For. Meteorol. 2013, 170, 206–215. [Google Scholar] [CrossRef]
- Porcar-Castell, A.; Tyystjarvi, E.; Atherton, J.; van der Tol, C.; Flexas, J.; Pfundel, E.E.; Moreno, J.; Frankenberg, C.; Berry, J.A. Linking chlorophyll a fluorescence to photosynthesis for remote sensing applications: Mechanisms and challenges. J. Exp. Bot. 2014, 65, 4065–4095. [Google Scholar] [CrossRef]
- Strasser, R.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. In Probing Photosynthesis: Mechanism, Regulation and Adaptation; Yunus, M., Pathre, U., Mohanty, P., Eds.; Taylor and Francis: London, UK, 2000; pp. 445–483. [Google Scholar]
- Brestic, M.; Zivcak, M. PSII fluorescence techniques for measurement of drought and high temperature stress signal in crop plants: Protocols and applications. In Molecular Stress Physiology of Plants; Rout, G., Das, A., Eds.; Springer: New Delhi, India, 2013; pp. 87–131. [Google Scholar]
- Van Heerden, P.D.; Tsimilli-Michael, M.; Krüger, G.H.; Strasser, R.J. Dark chilling effects on soybean genotypes during vegetative development: Parallel studies of CO2 assimilation, chlorophyll a fluorescence kinetics O-J-I-P and nitrogen fixation. Physiol. Plant. 2003, 117, 476–491. [Google Scholar] [CrossRef]
- Ripley, B.S.; Redfern, S.P.; Dames, J.F. Quantification of the photosynthetic performance of phosphorus-deficient Sorghum by means of chlorophyll-a fluorescence kinetics. S. Afr. J. Sci. South 2004, 100, 615–618. [Google Scholar]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the fluorescence transient. In Chlorophyll fluorescence: A signature of photosynthesis; Advances in Photosynthesis and Respiration Series; George, C., Papageorgiou, C., Govindjee, G., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 321–362. [Google Scholar]
- Mohanty, P.; Kreslavski, V.D.; Klimov, V.V.; Los, D.A.; Mimuro, M.; Carpentier, R.; Allakhverdiev, S.I. Heat stress: Susceptibility, recovery and regulation. In Photosynthesis: Plastid Biology, Energy Conservation and Carbon Assimilation; Eaton-Rye, J.J., Tripathy, B.C., Sharkey, T.D., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 251–274. [Google Scholar]
- Yan, K.; Chen, P.; Shao, H.; Zhao, S. Characterization of photosynthetic electron transport chain in bioenergy crop Jerusalem artichoke (Helianthus tuberosus L.) under heat stress for sustainable cultivation. Ind. Crop. Prod. 2013, 50, 809–815. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, A.; Li, X.; Lu, C. The Role of Chloroplast Gene Expression in Plant Responses to Environmental Stress. Int. J. Mol. Sci. 2020, 21, 6082. [Google Scholar] [CrossRef]
- Vener, A.V. Environmentally modulated phosphorylation and dynamics of proteins photosynthetic membranes. Biochim. Biophys. Acta 2007, 1767, 449–457. [Google Scholar] [CrossRef]
- Yao, W.B.; Meng, B.Y.; Tanaka, M.; Sugiura, M. An additional promoter within the protein-coding region of the psb D-psb C gene cluster in tobacco chloroplast DNA. Nucleic Acids Res. 1989, 17, 9583–9591. [Google Scholar] [CrossRef]
- Barber, J.; Nield, J.; Morris, E.P.; Zheleva, D.; Hankamer, B. The structure, function and dynamics of photosystem two. Physiol. Plant. 1997, 100, 817–827. [Google Scholar] [CrossRef]
- Vener, A.V.; Harms, A.; Sussman, M.R.; Vierstra, R.D. Mass spectrometric resolution of reversible protein phosphorylation in photosynthetic membranes of Arabidopsis thaliana. J. Biol. Chem. 2001, 276, 6959–6966. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J. Phosphorylation of chloroplast membrane polypeptides. Nature 1977, 269, 344–346. [Google Scholar] [CrossRef]
- Shi, L.X.; Schröder, W.P. The low molecular mass subunits of the photosynthetic supracomplex, photosystem II. Biochim. Biophys. Acta Bioenerg. 2004, 1608, 75–96. [Google Scholar] [CrossRef]
- Huang, W.; Yang, Y.J.; Hu, H.; Zhang, S.B.; Cao, K.F. Evidence for the role of cyclic electron flow in photoprotection for oxygen-evolving complex. J. Plant Physiol. 2016, 194, 54–60. [Google Scholar] [CrossRef]
- Janská, A.; Maršík, P.; Zelenková, S.; Ovesná, J. Cold stress and acclimation–what is important for metabolic adjustment? Plant Biol. 2010, 12, 395–405. [Google Scholar] [CrossRef]
- Palma, F.; López-Gómez, M.; Tejera, N.A.; Lluch, C. Salicylic acid improves the salinity tolerance of Medicago sativa in symbiosis with Sinorhizobium meliloti by preventing nitrogen fixation inhibition. Plant Sci. 2013, 208, 75–82. [Google Scholar] [CrossRef]
- Couée, I.; Sulmon, C.; Gouesbet, G.; El Amrani, A. Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants. J. Exp. Bot. 2006, 57, 449–459. [Google Scholar] [CrossRef]
- Signorelli, S.; Coitiño, E.L.; Borsani, O.; Monza, J. Molecular mechanisms for the reaction between •OH radicals and proline: Insights on the role as reactive oxygen species scavenger in plant stress. J. Phys. Chem. 2013, 118, 37–47. [Google Scholar] [CrossRef]
- Curtis, T.; Halford, N.G. Food security: The challenge of increasing wheat yield and the importance of not compromising food safety. Ann. Appl. Biol. 2014, 164, 354–372. [Google Scholar] [CrossRef]
- Chakrabarti, B.; Singh, S.D.; Kumar, V.; Harit, R.C.; Misra, S. Growth and yield response of wheat and chickpea crops under high temperature. Indian J. Plant Physiol. 2013, 18, 7–14. [Google Scholar] [CrossRef]
- Cossani, C.M.; Reynolds, M.P. Physiological traits for improving heat tolerance in wheat. Plant Physiol. 2012, 160, 1710–1718. [Google Scholar] [CrossRef]
- Narayanan, S. Effects of high temperature stress and traits associated with tolerance in wheat. Open Access. J. Sci. 2018, 2, 177–186. [Google Scholar]
- Agarie, S.; Hanaoka, N.; Ueno, O.; Miyazaki, A.; Kubota, F.; Agata, W.; Kaufman, P.B. Effects of silicon on tolerance to water deficit and heat stress in rice plants (Oryza sativa L.), monitored by electrolyte leakage. Plant Prod. Sci. 1998, 1, 96–103. [Google Scholar] [CrossRef]
- Sivanesan, I.; Son, M.; Soundararajan, P.; Jeong, B. Effect of silicon on growth and temperature stress tolerance of Nephrolepis exaltata “Corditas”. Korean J. Horticult. Sci. Technol. 2014, 32, 142–148. [Google Scholar] [CrossRef]
- Rios, J.J.; Martínez-Ballesta, M.C.; Ruiz, J.M.; Blasco, B.; Carvajal, M. Silicon-Mediated improvement in plant salinity tolerance: The role of aquaporins. Front. Plant Sci. 2017, 8, 948. [Google Scholar] [CrossRef]
- Younis, A.A.; Khattab, H.; Emam, M.M. Impacts of silicon and silicon nanoparticles on leaf ultrastructure and TaPIP1 and TaNIP2 gene expressions in heat stressed wheat seedlings. Biol. Plant. 2020, 64, 343–352. [Google Scholar] [CrossRef]
- Ma, D.; Sun, D.; Wang, C.; Qin, H.; Ding, H.; Li, Y.; Guo, T. Silicon application alleviates drought stress in wheat through transcriptional regulation of multiple antioxidant defense pathways. J. Plant Growth Regul. 2016, 35, 1–10. [Google Scholar] [CrossRef]
- Wang, X.; Gao, F.; Bing, J.; Sun, W.; Feng, X.; Ma, X.; Zhou, Y.; Zhang, G. Overexpression of the jojoba aquaporin gene, scpip1, enhances drought and salt tolerance in transgenic Arabidopsis. Int. J. Mol. Sci. 2019, 20, 153. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.; Tran, L.S.P. Impacts of priming with silicon on the growth and tolerance of maize plants to alkaline stress. Front. Plant Sci. 2016, 7, 243. [Google Scholar] [CrossRef]
- Alsaeedia, A.; El-Ramadyb, H.; Alshaa, T.; El-Garawanic, M.; Elhawat, N.; Al-Otaibi, A. Exogenous nanosilica improves germination and growth of cucumber by maintaining K+/Na+ ratio under elevated Na+ stress. Plant Physiol. Biochem. 2018, 125, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Rizwan, M.; Ali, Q.; Ali, S. Seed priming with silicon nanoparticles improved the biomass and yield while reduced the oxidative stress and cadmium concentration in wheat grains. Environ. Sci. Pollut. Res. 2019, 26, 7579–7588. [Google Scholar] [CrossRef] [PubMed]
- Bajwa, A.A.; Chauhan, B.S.; Adkins, S. Morphological, physiological and biochemical responses of two Australian biotypes of Parthenium hysterophorus to different soil moisture regimes. Environ. Sci. Pollut. Res. 2017, 24, 16186–16194. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, J.C.; Moya, T.B. Genotypic variation in maintenance of leaf water potential in rice. Crop Sci. 1978, 18, 873–876. [Google Scholar] [CrossRef]
- Stirbet, A.; Govindjee, G. On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and photosystem II: Basics and applications of the OJIP fluorescence transient. J. Photochem. Photobiol. B Biol. 2011, 104, 236–257. [Google Scholar] [CrossRef]
- Metzner, H.; Rau, H.; Senger, H. Untersuchungen zur synchronisierbarkeit einzelner pigmentmangel-mutanten von Chlorella. Planta 1965, 65, 186–194. [Google Scholar] [CrossRef]
- Fairbairn, N.J. A modified anthrone reagent. Chem. Ind. 1953, 4, 285–313. [Google Scholar]
- Hubbard, N.L.; Pharr, D.M.; Huber, S.C. Role of sucrose phosphate synthase in sucrose biosynthesis in ripening bananas and its relationship to the respiratory climacteric. Plant Physiol. 1990, 94, 201–208. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free prolin for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Valentovič, P.; Luxová, M.; Kolarovič, L.; Gašparíková, O. Effect of osmotic stress on compatible solutes content, membrane stability and water relations in maize cultivars. Plant Soil Environ. 2006, 52, 186–191. [Google Scholar] [CrossRef]
- Angelo, A.J.S.; Ory, R.L.; Brown, L.E. Comparison of methods for determining peroxidation in processed whole peanut products. J. Am. Oil Chem. Soc. 1975, 52, 34–35. [Google Scholar] [CrossRef]
- Fishwick, M.J.; Swoboda, P.A. Measurement of oxidation of polyunsaturated fatty acids by spectrophotometric assay of conjugated derivatives. J. Sci. Food Agric. 1977, 28, 387–393. [Google Scholar] [CrossRef]
- Tenea, G.N.; Bota, A.P.; Raposo, F.C.; Maquet, A. Reference genes for gene expression studies in wheat flag leaves grown under different farming conditions. BMC Res. Notes 2011, 4, 373. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef]
- Nolla, A.; de Faria, R.J.; Korndörfer, G.H.; da Silva, T.R.B. Effect of silicon on drought tolerance of upland rice. J. Food Agric. Environ. 2012, 10, 269–272. [Google Scholar]
- Ashkavand, P.; Tabari, M.; Zarafshar, M.; Tomášková, I.; Struve, D. Effect of SiO2 nanoparticles on drought resistance in hawthorn seedlings. For. Res. Pap. 2015, 76, 350–359. [Google Scholar] [CrossRef]
- Emam, M.M.; Khattab, H.I.; Helal, N.M. Effects of silicon or selenium on photosynthetic apparatus and antioxidant capacity of rice grown under drought condition. Egypt. J. Exp. Biol. 2012, 8, 271–283. [Google Scholar]
- Saud, S.; Li, X.; Chen, Y.; Zhang, L.; Fahad, S.; Hussain, S.; Sadiq, A.; Chen, Y. Silicon application increases drought tolerance of Kentucky blue grass by improving plant water relations and morphophysiological functions. Sci. World J. 2014, 2014. [Google Scholar] [CrossRef]
- Avestan, S.; Ghasemnezhad, M.; Esfahani, M.; Byrt, C.S. Application of Nano-Silicon Dioxide Improves Salt Stress Tolerance in Strawberry. Plant Agron. 2019, 9, 246. [Google Scholar] [CrossRef]
- Sun, D.; Hussain, H.I.; Yi, Z.; Siegele, R.; Cresswell, T.; Kong, L.; Cahill, D.M. Uptake and cellular distribution, in four plant species of fluorescently labeled mesoporous silica nanoparticles. Plant Cell Rep. 2014, 33, 1389–1402. [Google Scholar] [CrossRef]
- El Basyoni, I.; Saadalla, M.; Baenziger, S.; Bockelman, H.; Morsy, S. Cellmembrane stability and association mapping for drought and heat tolerance in a worldwide wheat collection. Sustainability 2017, 9, 1606. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Rehman, M.Z.; Malik, S.; Adrees, M.; Qayyum, M.F.; Alamri, S.A.; Alyemeni, M.N.; Ahmad, P. Effect of foliar applications of silicon and titanium dioxide nanoparticles on growth, oxidative stress, and cadmium accumulation by rice (Oryza sativa). Acta Physiol. Plant. 2019, 41, 35. [Google Scholar] [CrossRef]
- Nazaralian, S.; Majd, A.; Irian, S.; Najafi, F.; Ghahremaninejad, F.; Landberg, T.; Greger, M. Comparison of silicon nanoparticles and silicate treatments in fenugreek. Plant Physiol. Biochem. 2017, 115, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Merwad, A.R.M.; Desoky, E.S.M.; Rady, M.M. Response of water deficit-stressed Vigna unguiculata performances to silicon, proline or methionine foliar application. Sci. Hortic. 2018, 228, 132–144. [Google Scholar] [CrossRef]
- Li-Feng, W.; Hao, F.; Yun-He, J. Photosynthetic characterization of a rolled leaf mutant of rice (Oryza sativa L.). Afr. J. Biotechnol. 2012, 11, 6839–6846. [Google Scholar] [CrossRef]
- Chen, Q.; Zhao, X.; Lei, D.; Hu, S.; Shen, Z.; Shen, W.; Xu, X. Hydrogen-Rich water pretreatment alters photosynthetic gas exchange, chlorophyll fluorescence, and antioxidant activities in heat-stressed cucumber leaves. Plant Growth Regul. 2017, 83, 69–82. [Google Scholar] [CrossRef]
- Jumrani, K.; Bhatia, V.S.; Pandey, G.P. Impact of elevated temperatures on specific leaf weight, stomatal density, photosynthesis and chlorophyll fluorescence in soybean. Photosynth. Res. 2017, 131, 333–350. [Google Scholar] [CrossRef]
- Gosavi, G.U.; Jadhav, A.S.; Kale, A.A.; Gadakh, S.R.; Pawar, B.D.; Chimote, V.P. Effect of heat stress on proline, chlorophyll content, heat shock proteins and antioxidant enzyme activity in sorghum (Sorghum bicolor) at seedlings stage. Indian J. Biotechnol. 2014, 13, 356–363. [Google Scholar]
- Yüzbaşıoğlu, E.; Dalyan, E.; Akpınar, I. Changes in photosynthetic pigments, anthocyanin content and antioxidant enzyme activities of maize (Zea mays L.) seedlings under high temperature stress conditions. Trakya Univ. J. Nat. Sci. 2017, 18, 97–104. [Google Scholar]
- Djanaguiraman, M.; Boyle, D.L.; Welti, R.; Jagadish, S.V.K.; Prasad, P.V.V. Decreased photosynthetic rate under high temperature in wheat is due to lipid desaturation, oxidation, acylation, and damage of organelles. BMC Plant Biol. 2018, 18, 55. [Google Scholar] [CrossRef] [PubMed]
- Teramura, A.H.; Sullivan, J.H. Effects of UV-B radiation on photosynthesis and growth of terrestrial plants. Photosynth. Res. 1994, 39, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Cicek, N.; Fedina, I.; Çakirlar, H.; Velitchkova, M.; Georgieva, K. The role of short-term high temperature pretreatment on the UV-B tolerance of barley cultivars. Turk. J. Agric. For. 2012, 36, 153–165. [Google Scholar]
- Tyutereva, E.V.; Evkaikina, A.I.; Ivanova, A.N.; Voitsekhovskaja, O.V. The absence of chlorophyll b affects lateral mobility of photosynthetic complexes and lipids in grana membranes of Arabidopsis and barley chlorina mutants. Photosynth. Res. 2017, 133, 357–370. [Google Scholar] [CrossRef]
- Donegá, M.A. Ratio K: Ca and Application of Silicon in the Nutrient Solution for the Hydroponic Cultivation of Coriander. Master’s Thesis, ESALQ, Piracicaba, Brazil, 2009; pp. 1–62. [Google Scholar]
- Silva, E.N.; Vieira, S.A.; Ribeiro, R.V.; Ponte, L.F.; Ferreira-Silva, S.L.; Silveira, J.A. Contrasting physiological responses of Jatropha curcas plants to single and combined stresses of salinity and heat. J. Plant Growth Regul. 2013, 32, 159–169. [Google Scholar] [CrossRef]
- Cao, B.L.; Ma, Q.; Zhao, Q.; Wang, L.; Xu, K. Effects of silicon on absorbed light allocation, antioxidant enzymes and ultrastructure of chloroplasts in tomato leaves under simulated drought stress. Sci. Hortic. 2015, 194, 53–62. [Google Scholar] [CrossRef]
- Ju, S.M.; Wang, L.P.; Chen, J.Y. Effects of silicon on the growth, photosynthesis and chloroplast ultrastructure of Oryza sativa L. seedlings under acid rain stress. Silicon 2020, 12, 655–664. [Google Scholar] [CrossRef]
- Haghighi, M.; Pessarakli, M. Influence of silicon and nano-silicon on salinity tolerance of cherry tomatoes (Solanum lycopersicum L.) at early growth stage. Sci. Hortic. 2013, 61, 111–117. [Google Scholar] [CrossRef]
- Noji, T.; Kamidaki, C.; Kawakami, K.; Shen, J.R.; Kajino, T.; Fukushima, Y.; Sekitoh, T.; Itoh, S. Photosynthetic oxygen evolution in mesoporous silica material: Adsorption of photosystem II reaction center complex into 23 nm nanopores in SBA. Langmuir 2010, 27, 705–713. [Google Scholar] [CrossRef]
- Colom, M.R.; Vazzana, C. Photosynthesis and PSII functionality of drought-resistant and drought-sensitive weeping love grass plants. Environ. Exp. Bot. 2003, 49, 135–144. [Google Scholar] [CrossRef]
- Yoshioka, M.; Uchida, S.; Mori, H.; Komayama, K.; Ohira, S.; Morita, N.; Nakanishi, T.; Yamamoto, Y. Quality control of photosystem II cleavage of reaction center D1 protein in spinach thylakoids by FtsH protease under moderate heat stress. J. Biol. Chem. 2006, 281, 21660–21669. [Google Scholar] [CrossRef] [PubMed]
- Komayama, K.; Khatoon, M.; Takenaka, D.; Horie, J.; Yamashita, A.; Yoshioka, M.; Nakayama, Y.; Yoshida, M.; Ohira, S.; Morita, N.; et al. Quality control of photosystem II: Cleavage and aggregation of heat-damaged D1 protein in spinach thylakoids. Biochim. Biophys. Acta Bioenerg. 2007, 1767, 838–846. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Hu, Z.; Xie, Y.; Chan, Z.; Chen, K.; Amombo, E.; Chen, L.; Fu, J. Alleviation of cold damage to photosystem II and metabolisms by melatonin in Bermuda grass. Front. Plant Sci. 2015, 6, 925. [Google Scholar] [CrossRef] [PubMed]
- Ghassemi-Golezani, K.; Lotfi, R. The impact of salicylic acid and silicon on chlorophyll a fluorescence in mung bean under salt stress. Russ. J. Plant Physiol. 2015, 62, 611–616. [Google Scholar] [CrossRef]
- Maghsoudi, K.; Emam, Y.; Ashraf, M. Influence of foliar application of silicon on chlorophyll fluorescence, photosynthetic pigments, and growth in water-stressed wheat cultivars differing in drought tolerance. Turk. J. Bot. 2015, 39, 625–634. [Google Scholar] [CrossRef]
- Hussain, M.; Khan, T.A.; Yusuf, M.; Fariduddin, Q. Silicon-Mediated role of 24-epibrassinolide in wheat under high-temperature stress. Environ. Sci. Pollut. Res. 2019, 26, 17163–17172. [Google Scholar] [CrossRef]
- Han, Y.; Fan, S.; Zhang, Q.; Wang, Y. Effect of heat stress on the MDA, proline and soluble sugar content in leaf lettuce seedlings. Agric. Sci. 2013, 4, 112. [Google Scholar] [CrossRef]
- Mohamed, H.I.; Abdel-Hamid, A.M.E. Molecular and biochemical studies for heat tolerance on four cotton genotypes. Rom. Biotechnol. Lett. 2013, 18, 8823–8831. [Google Scholar]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef]
- Kumar, S.; Gupta, D.; Nayyar, H. Comparative response of maize and rice genotypes to heat stress: Status of oxidative stress and antioxidants. Acta Physiol. Plant. 2012, 34, 75–86. [Google Scholar] [CrossRef]
- Kohila, S.; Gomathi, R. Adaptive physiological and biochemical response of sugarcane genotypes to high-temperature stress. Indian J. Plant Physiol. 2018, 23, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Hameed, A.; Goher, M.; Iqbal, N. Heat stress-induced cell death, changes in antioxidants, lipid peroxidation, and protease activity in wheat leaves. J. Plant Growth Regul. 2012, 31, 283–291. [Google Scholar] [CrossRef]
- Yin, L.; Wang, S.; Li, J.; Tanaka, K.; Oka, M. Application of silicon improves salt tolerance through ameliorating osmotic and ionic stresses in the seedling of Sorghum bicolor. Acta Physiol. Plant. 2013, 35, 3099–3107. [Google Scholar] [CrossRef]
- Lana, R.M.Q.; Korndorfer, G.; Zanao-Junior, L.; Silva, A.; Lana, A.M.Q. Effect of calcium silicate on the productivity and silicon accumulation in the tomato plant. Biosci. J. 2003, 19, 15–20. [Google Scholar]
- Meena, V.D.; Dotaniya, M.L.; Coumar, V.; Rajendiran, S.; Kundu, S.; Rao, A.S. A case for silicon fertilization to improve crop yields in tropical soils. Proc. Natl. Acad. Sci. India Sect. B J. Biol. Sci. 2014, 84, 505–518. [Google Scholar] [CrossRef]
- Kalteh, M.; Alipour, Z.T.; Ashraf, S.; Aliabadi, M.M.; Nosratabadi, A.F. Effect of silica nanoparticles on Basil (Ocimum basilicum) under salinity stress. J. Chem. Health Saf. 2014, 4, 49–55. [Google Scholar]
- Abdel-Haliem, M.E.F.; Hegazy, H.S.; Hassan, N.S.; Naguib, D.M. Effect of silica ions and nano silica on rice plants under salinity stress. Ecol. Eng. 2017, 99, 282–289. [Google Scholar] [CrossRef]
- Lípová, L.; Krchňák, P.; Komenda, J.; Ilík, P. Heat-Induced disassembly and degradation of chlorophyll-containing protein complexes in vivo. Biochim. Biophys. Acta 2010, 1797, 63–70. [Google Scholar] [CrossRef]
- Ifuku, K.; Endo, T.; Shikanai, T.; Aro, E. Structure of the Chloroplast NADH Dehydrogenase-Like Complex: Nomenclature for Nuclear-Encoded Subunits. Plant Cell Physiol. 2011, 52, 1560–1568. [Google Scholar] [CrossRef]
- Fristedt, R.; Willig, A.; Granath, P.; Crèvecoeur, M.; Rochaix, J.D.; Vener, A.V. Phosphorylation of photosystem II controls functional macroscopic folding of photosynthetic membranes in Arabidopsis. Plant Cell 2009, 21, 3950–3964. [Google Scholar] [CrossRef]
- Vener, A.V.; Rokka, A.; Fulgosi, H.; Andersson, B.; Herrmann, R.G. A cyclophilin-regulated PP2A-like protein phosphatase in thylakoid membranes of plant chloroplasts. Biochemistry 1999, 38, 14955–14965. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Chen, Y.E.; Tian, W.J.; Du, J.B.; Zhang, Z.W.; Xu, F.; Zhang, F.; Yuan, S.; Lin, H.H. Dephosphorylation of photosystem II proteins and phosphorylation of CP29 in barley photosynthetic membranes as a response to water stress. Biochim. Biophys. Acta Bioenerg. 2009, 1787, 1238–1245. [Google Scholar] [CrossRef] [PubMed]
- Varghese, N.; Alyammahi, O.; Nasreddine, S.; Alhassani, A.; Gururani, M.A. Melatonin positively influences the photosynthetic machinery and antioxidant system of Avena sativa during salinity stress. Plants 2019, 8, 610. [Google Scholar] [CrossRef] [PubMed]
- Rantala, M.; Rantala, S.; Aro, E.M. Composition, phosphorylation and dynamic organization of photosynthetic protein complexes in plant thylakoid membrane. Photochem. Photobiol. Sci. 2020, 19, 604–619. [Google Scholar] [CrossRef]
- Gururani, M.A.; Venkatesh, J.; Tran, L.S.P. Regulation of photosynthesis during abiotic stress-induced photoinhibition. Mol. Plant 2015, 8, 1304–1320. [Google Scholar] [CrossRef]
- Sasi, S.; Venkatesh, J.; Daneshi, R.F.; Gururani, M.A. Photosystem II extrinsic proteins and their putative role in abiotic stress tolerance in higher plants. Plants 2018, 7, 100. [Google Scholar] [CrossRef]
- Wei, X.; Su, X.; Cao, P.; Liu, X.; Chang, W.; Li, M.; Zhang, X.; Liu, Z. Structure of spinach photosystem II–LHCII super complex at 3.2 Å resolution. Nature 2016, 534, 69. [Google Scholar] [CrossRef]
- Takahashi, S.; Nakamura, T.; Sakamizu, M.; Woesik, R.V.; Yamasaki, H. Repair machinery of symbiotic photosynthesis as the primary target of heat stress for reef-building corals. Plant Cell Physiol. 2004, 45, 251–255. [Google Scholar] [CrossRef]
- Kamiya, N.; Shen, J.R. Crystal structure of oxygen-evolving photosystem II from Thermosynechococcus vulcanus at 3.7-Å resolution. Proc. Natl. Acad. Sci. USA 2003, 100, 98–103. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
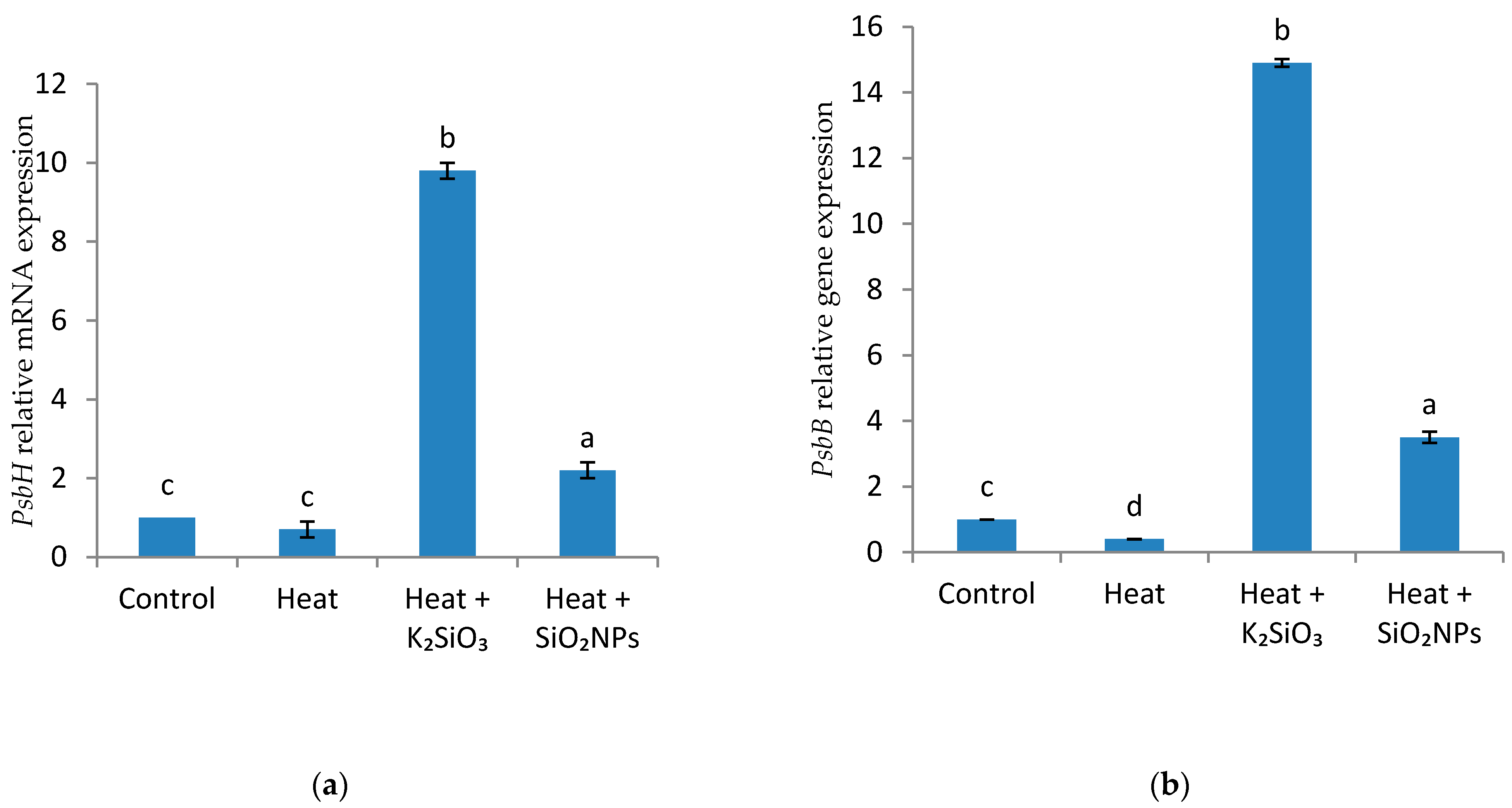{kind=link}
{kind=link}
| Primer Name | Primer Sequence 5′-3′ | Accession Number | Gene Name |
|---|---|---|---|
| PsbHF | TGGCTACACAAACCGTTGAA | NC_002762.1 (70762..70983) NC_002762.1 (68672..70198) NC_002762.1 (8995..10056) | Photosystem II Reaction center protein H |
| PsbHR | CCGTCCAGTAAAACGGAAGA | ||
| PsbBF | GGTTTGCCTTGGTATCGTGT | ||
| PsbBR | TCCACATTGGATCCAGAACA | ||
| PsbDF | CGCTTTAGGGGGTGTGTTTA | ||
| PsbDR | GCCCCCATAGTAGCAACAAA | ||
| TaActinF | TGCTATCCTTCGTTTGGACCTT | AB181991.1 | |
| TaActin R | AGCGGTTGTTGTGAGGGAGT |
| Treatments | Chl (a) | Chl (b) | Chl (a + b) | Carotenoids | Chlorophyll a/b Ratio |
|---|---|---|---|---|---|
| Control | 11.3 ± 0.17 a | 5.5 ± 0.17 a | 16.8 ± 0.12 a | 2.5 ± 0.06 a | 2.05 ± 0.08 b |
| Heat | 6.5 ± 0.12 c | 2.1 ± 0.06 c | 8.6 ± 0.12 d | 1.6 ± 0.05 c | 3.0 ± 0.15 a |
| Heat+ K2SiO3 | 10.8 ± 0.17 a | 5.3 ± 0.13 a | 16.1 ± 0.12 b | 2.2 ± 0.52 b | 2.3 ± 0.17 b |
| Heat+ SiO2NPs | 9.8 ± 0.23 b | 4.3 ± 0.17 b | 14.1 ± 0.04 c | 2 ± 0.11 b | 2.03 ± 0.09 b |
| Treatments | Total Soluble Sugars | Sucrose | Proline |
|---|---|---|---|
| Control | 113.6 ± 0.036 d | 2.29 ± 0.030 d | 26.8 ± 0.69 d |
| Heat | 223.5 ± 0.034 c | 3.06 ± 0.040 c | 46 ± 0.28 c |
| Heat+ K2SiO3 | 330.0 ± 0.06 a | 5.54 ± 0.037 a | 86 ± 0.90 a |
| Heat+ SiO2NPs | 328.6 ±0.04 b | 5.28 ± 0.028 b | 57 ± 0.40 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, H.; Alatawi, A.; Abdulmajeed, A.; Emam, M.; Khattab, H. Roles of Si and SiNPs in Improving Thermotolerance of Wheat Photosynthetic Machinery via Upregulation of PsbH, PsbB and PsbD Genes Encoding PSII Core Proteins. Horticulturae 2021, 7, 16. https://doi.org/10.3390/horticulturae7020016
Hassan H, Alatawi A, Abdulmajeed A, Emam M, Khattab H. Roles of Si and SiNPs in Improving Thermotolerance of Wheat Photosynthetic Machinery via Upregulation of PsbH, PsbB and PsbD Genes Encoding PSII Core Proteins. Horticulturae. 2021; 7(2):16. https://doi.org/10.3390/horticulturae7020016
Chicago/Turabian StyleHassan, Heba, Aishah Alatawi, Awatif Abdulmajeed, Manal Emam, and Hemmat Khattab. 2021. "Roles of Si and SiNPs in Improving Thermotolerance of Wheat Photosynthetic Machinery via Upregulation of PsbH, PsbB and PsbD Genes Encoding PSII Core Proteins" Horticulturae 7, no. 2: 16. https://doi.org/10.3390/horticulturae7020016