
Neurobehavioral Effects of Fermented Rice Bran Extract in Zebrafish Larvae Model
 , , , and
, , , and 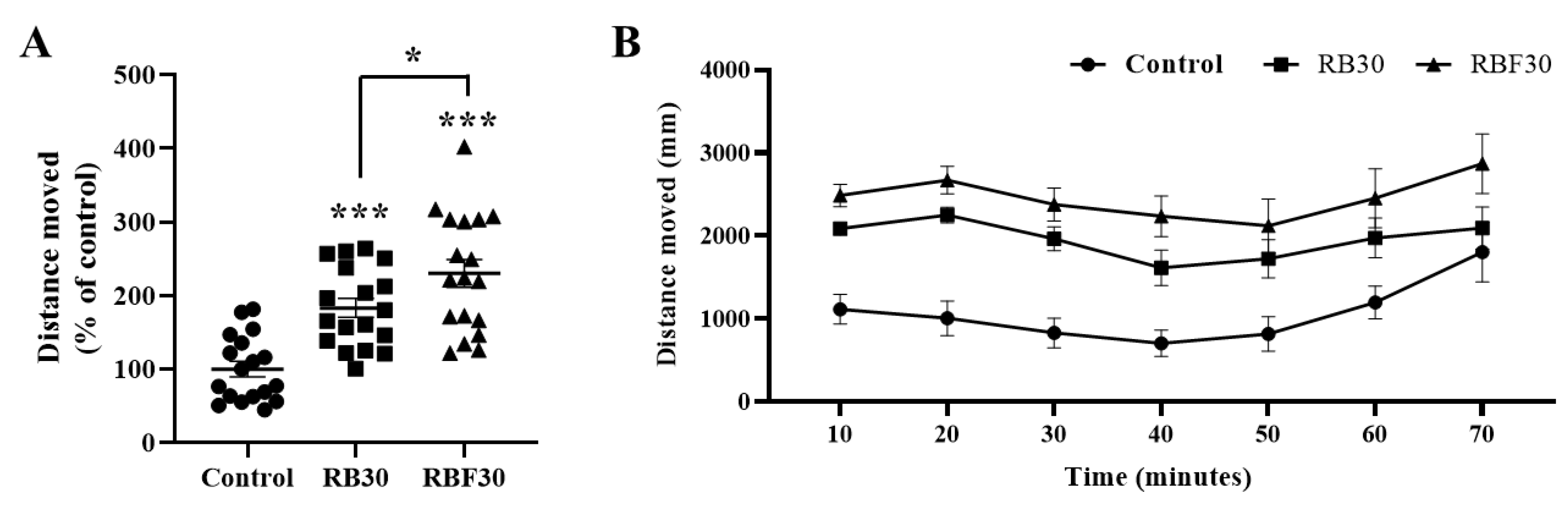{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fermentation and Purification of RBF30
2.2. Ultra-Performance Liquid Chromatography (UPLC) Analysis
2.3. Maintenance of Zebrafish
2.4. Behavioral Analysis of Zebrafish Larvae
2.5. Assessments of Neural Activity in Zebrafish Larvae
2.6. Measurement of Endogenous Neurochemicals in Zebrafish Larvae
2.7. Statistical Analysis
3. Results
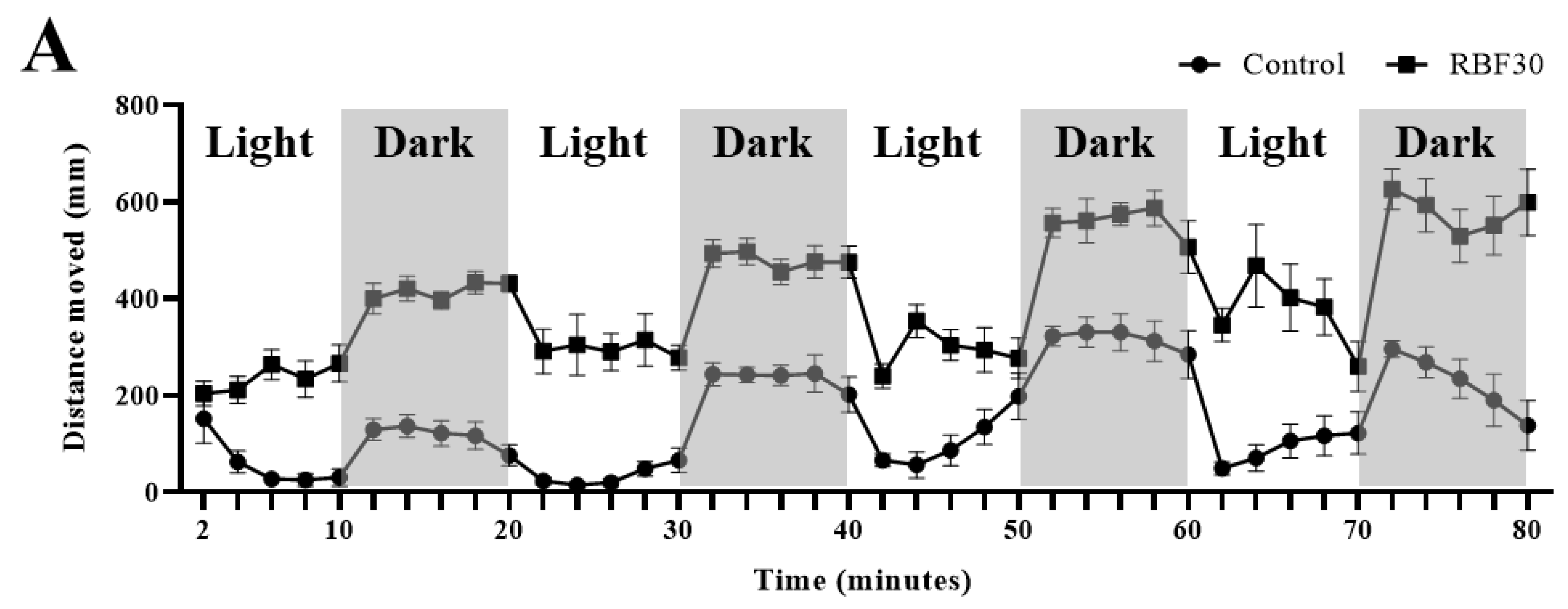
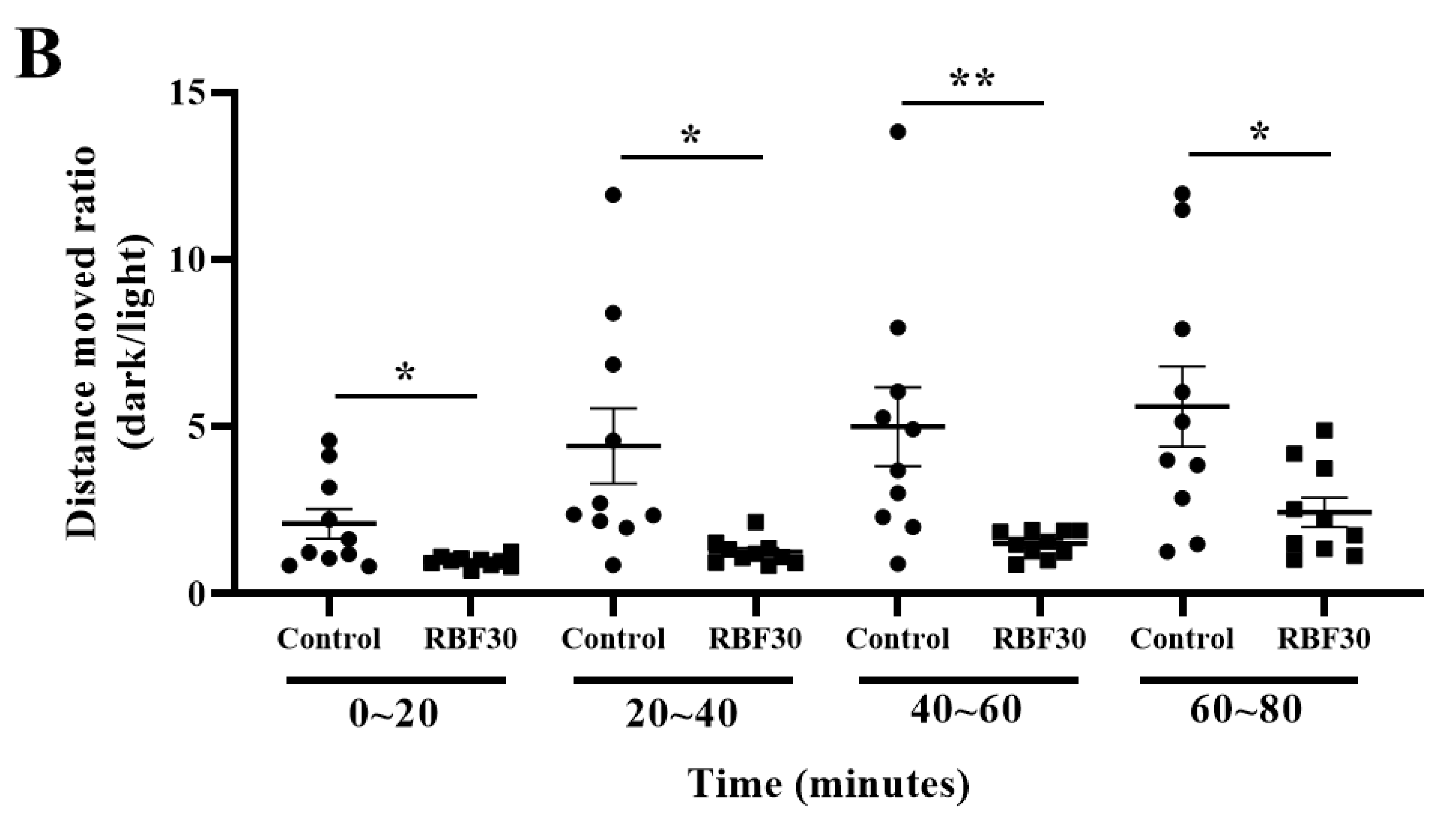
3.1. RBF30 Increased Locomotor Activity and Alleviated Anxiety-Like Behavior
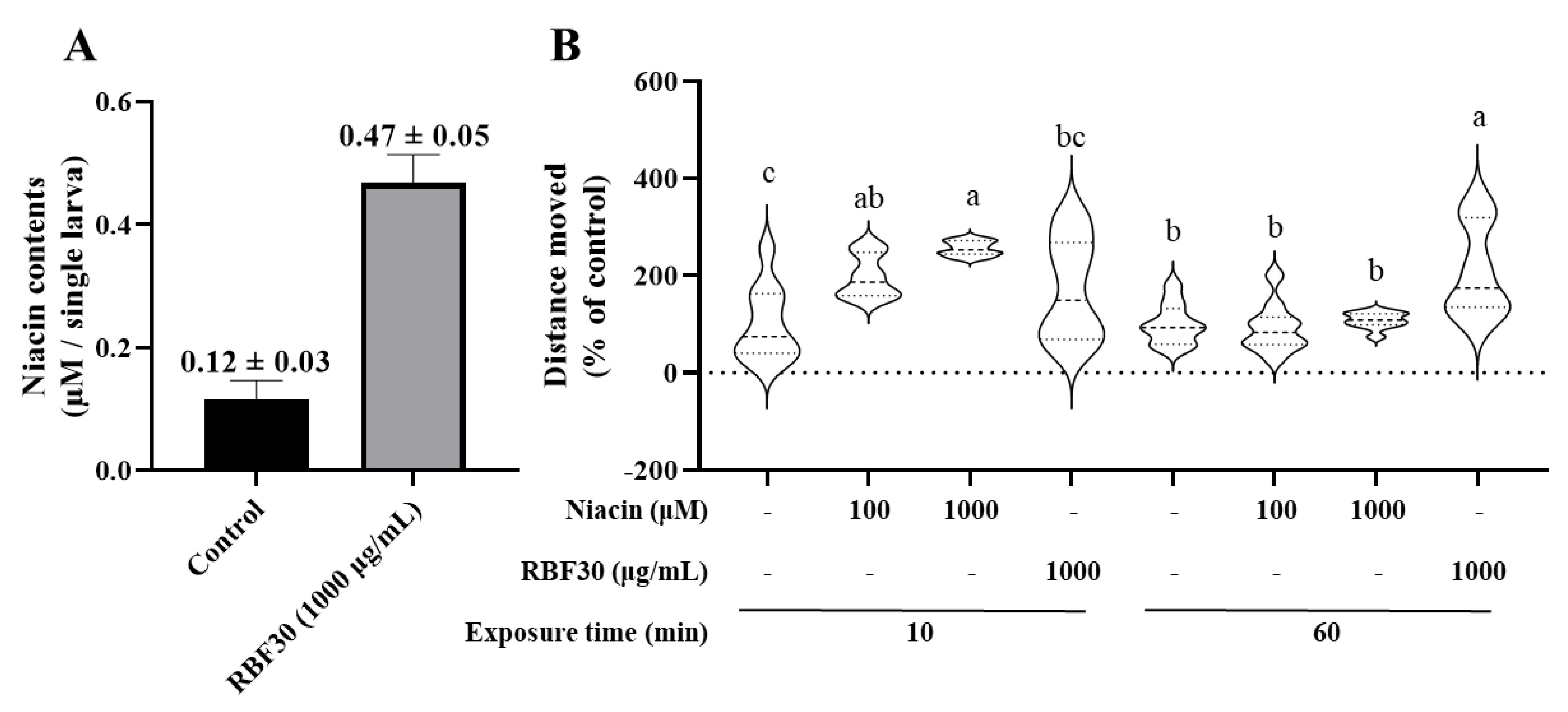
3.2. RBF30 Contains More Niacin Than RB30
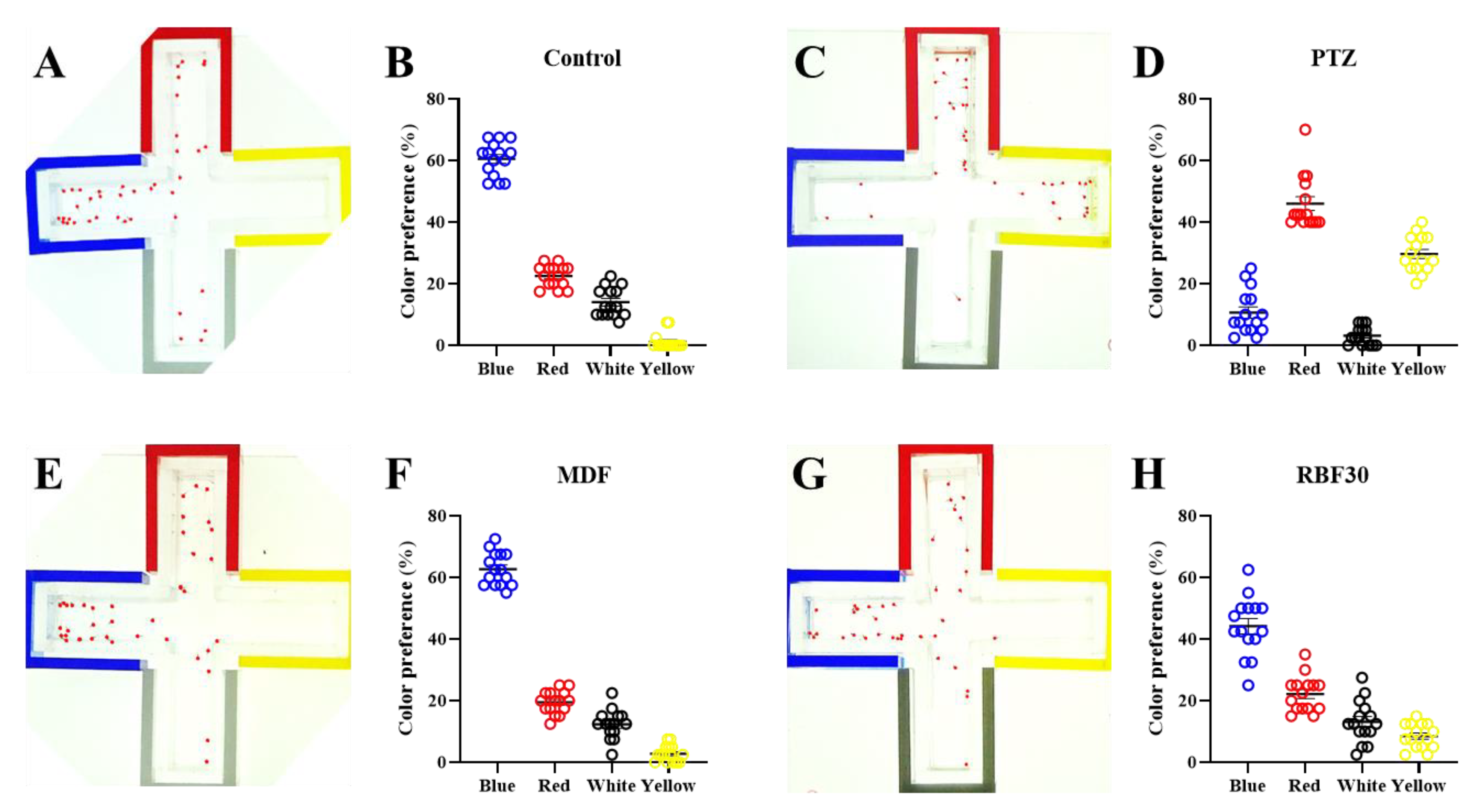
3.3. RBF30 Did Not Affect Innate Color Preference
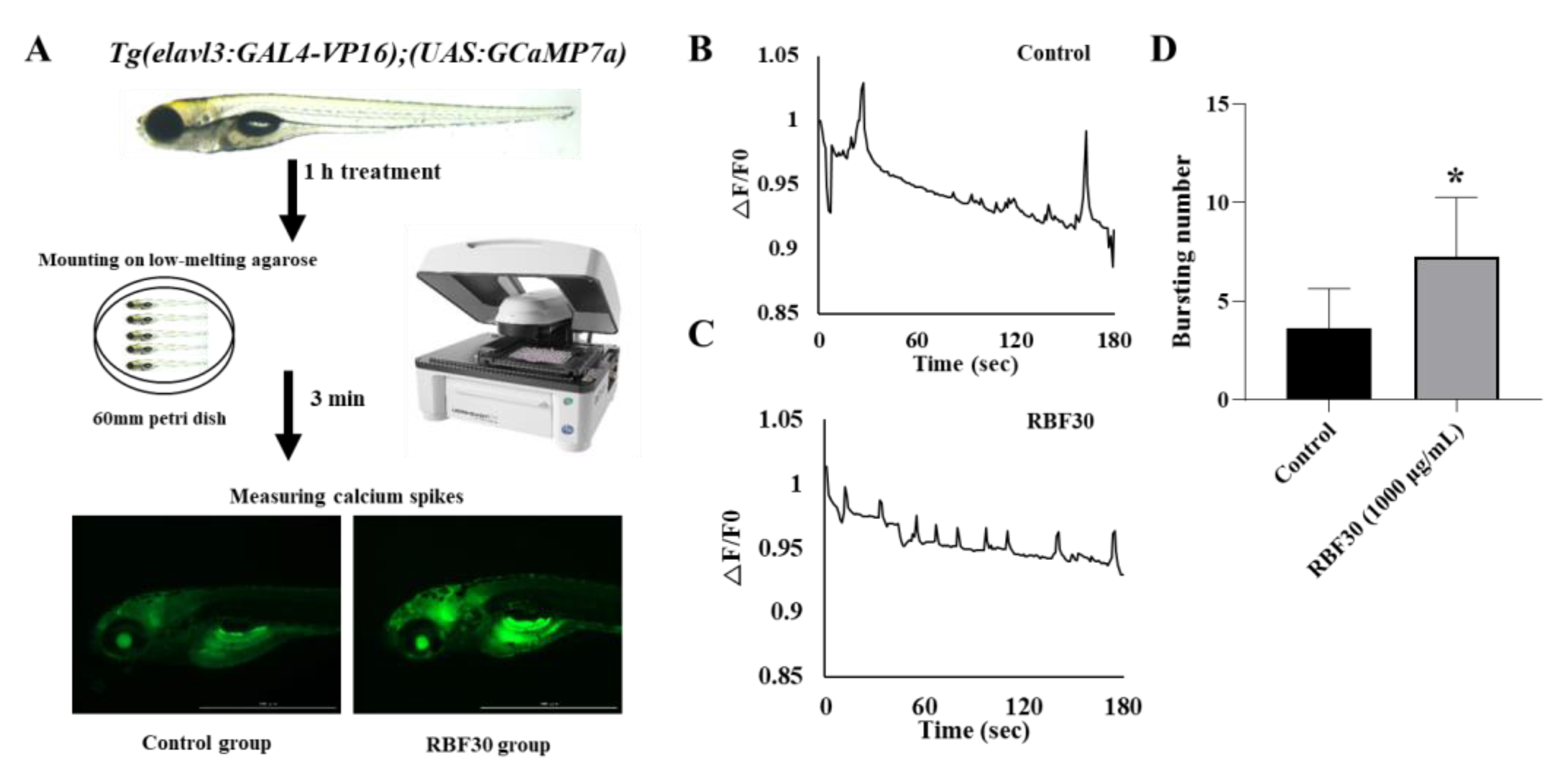
3.4. RBF30 Stimulate the Neural Activity
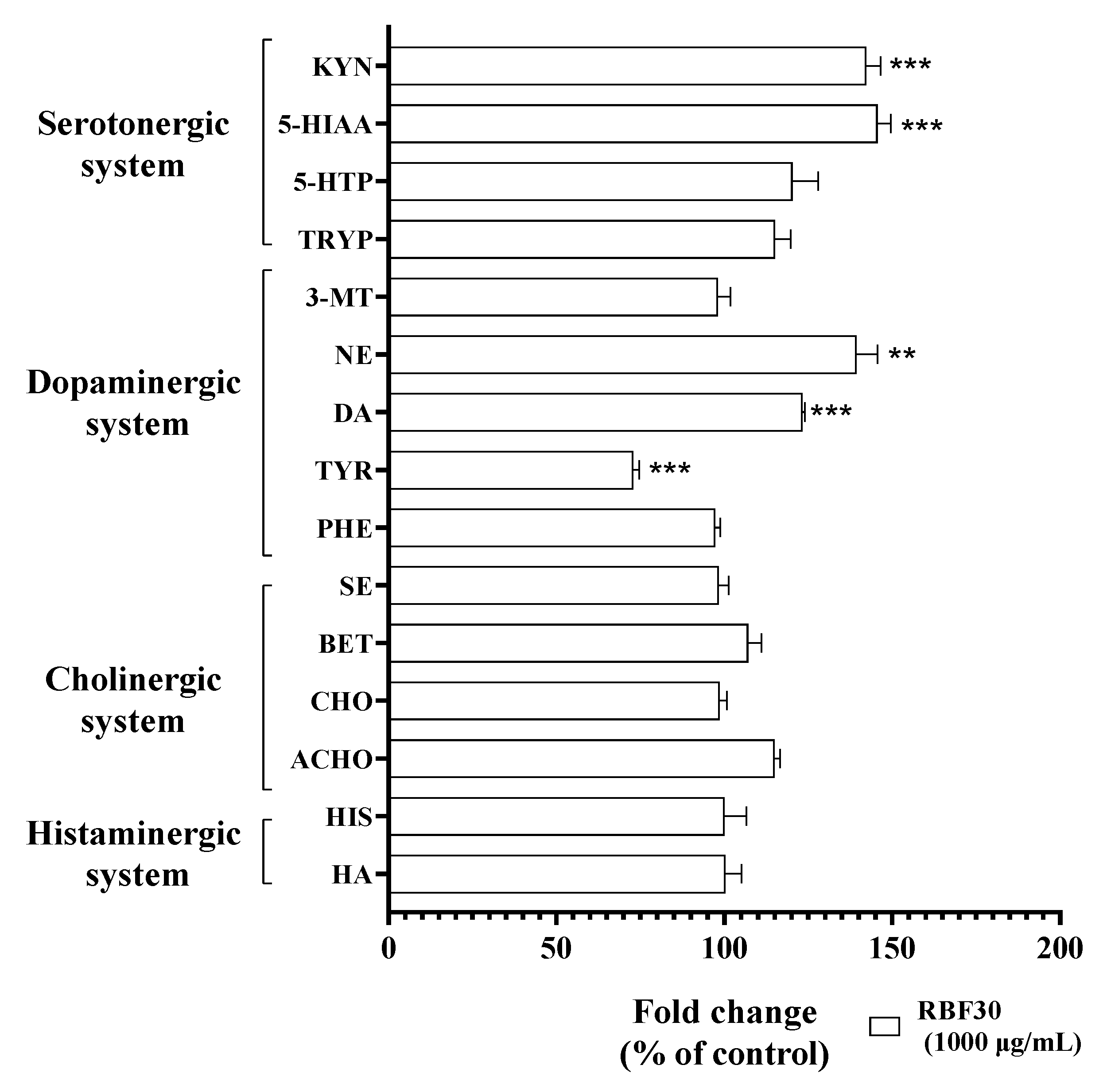
3.5. RBF30-Regulated Neurochemistry
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Boonloh, K.; Kukongviriyapan, V.; Kongyingyoes, B.; Kukongviriyapan, U.; Thawornchinsombut, S.; Pannangpetch, P. Rice bran protein hydrolysates improve insulin resistance and decrease pro-inflammatory cytokine gene expression in rats fed a high carbohydrate-high fat diet. Nutrients 2015, 7, 6313–6329. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.; Bhanger, M.; Anwar, F. Antioxidant properties and components of some commercially available varieties of rice bran in Pakistan. Food Chem. 2005, 93, 265–272. [Google Scholar] [CrossRef]
- Kim, H.-Y.; Kim, J.-H.; Yang, S.-B.; Hong, S.-G.; Lee, S.-A.; Hwang, S.-J.; Shin, K.-S.; Suh, H.-J.; Park, M.-H. A polysaccharide extracted from rice bran fermented with Lentinus edodes enhances natural killer cell activity and exhibits anticancer effects. J. Med. Food 2007, 10, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Shibata, A.; Kawakami, Y.; Kimura, T.; Miyazawa, T.; Nakagawa, K. α-tocopherol attenuates the triglyceride-and cholesterol-lowering effects of rice bran tocotrienol in rats fed a western diet. J. Agric. Food Chem. 2016, 64, 5361–5366. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, N.; Rodriguez-Alegria, M.; Gonzalez, F.; Lopez-Munguia, A. Enzymatic treatment of rice bran to improve processing. J. Am. Oil Chem. Soc. 2000, 77, 177–180. [Google Scholar] [CrossRef]
- Gopinger, E.; Ziegler, V.; da Silva Catalan, A.A.; Krabbe, E.L.; Elias, M.C.; Xavier, E.G. Whole rice bran stabilization using a short chain organic acid mixture. J. Stored Prod. Res. 2015, 61, 108–113. [Google Scholar] [CrossRef]
- Alauddina, M.; Islama, J.; Shirakawaa, H.; Kosekib, T.; Ardiansyahc, K.M. Rice bran as a functional food: An overview of the conversion of rice bran into a superfood/functional food. In Superfood and Functional Food-An Overview of Their Processing and Utilization; InTech: London, UK, 2017. [Google Scholar] [CrossRef]
- Hong, J.H.; Kim, J.Y.; Baek, S.E.; Ingkasupart, P.; Park, H.J.; Kang, S.G. Effects of rice bran extracts fermented with Lactobacillus plantarum on neuroprotection and cognitive improvement in a rat model of ischemic brain Injury. Biomed. Sci. Lett. 2015, 21, 92–102. [Google Scholar] [CrossRef]
- Kupski, L.; Cipolatti, E.; Rocha, M.D.; Oliveira, M.d.S.; Souza-Soares, L.d.A.; Badiale-Furlong, E. Solid-state fermentation for the enrichment and extraction of proteins and antioxidant compounds in rice bran by Rhizopus oryzae. Braz. Arch. Biol. Technol. 2012, 55, 937–942. [Google Scholar] [CrossRef]
- Lee, S.H.; Yeo, D.; Hong, J.H. Effect of dihydroferulic acid obtained from fermented rice bran extract on neuroprotection and behavioral recovery in an ischemic rat model. Food Sci. Technol. 2020, 40, 475–481. [Google Scholar] [CrossRef]
- Hagl, S.; Asseburg, H.; Heinrich, M.; Sus, N.; Blumrich, E.-M.; Dringen, R.; Frank, J.; Eckert, G.P. Effects of long-term rice bran extract supplementation on survival, cognition and brain mitochondrial function in aged NMRI mice. Neuromol. Med. 2016, 18, 347–363. [Google Scholar] [CrossRef]
- Islam, J.; Koseki, T.; Watanabe, K.; Budijanto, S.; Oikawa, A.; Alauddin, M.; Goto, T.; Aso, H.; Komai, M.; Shirakawa, H. Dietary supplementation of fermented rice bran effectively alleviates dextran sodium sulfate-induced colitis in mice. Nutrients 2017, 9, 747. [Google Scholar] [CrossRef] [PubMed]
- Saba, E.; Lee, C.H.; Da Hye Jeong, K.L.; Kim, T.-H.; Roh, S.-S.; Kim, S.-H.; Rhee, M.H. Fermented rice bran prevents atopic dermatitis in DNCB-treated NC/Nga mice. J. Biomed. Res. 2016, 30, 334. [Google Scholar]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.S.; Kan, H.; Hwang, K.-S.; Yang, J.Y.; Son, Y.; Shin, D.-S.; Lee, B.H.; Ahn, S.H.; Ahn, J.H.; Cho, S.-H. Neurochemical Effects of 4-(2Chloro-4-Fluorobenzyl)-3-(2-Thienyl)-1, 2, 4-Oxadiazol-5 (4H)-One in the Pentylenetetrazole (PTZ)-Induced Epileptic Seizure Zebrafish Model. Int. J. Mol. Sci. 2021, 22, 1285. [Google Scholar] [CrossRef] [PubMed]
- Gupta, T.; Mullins, M.C. Dissection of organs from the adult zebrafish. JoVE (J. Vis. Exp.) 2010, 37, e1717. [Google Scholar]
- Kim, S.S.; Im, S.H.; Yang, J.Y.; Lee, Y.-R.; Kim, G.R.; Chae, J.S.; Shin, D.-S.; Song, J.S.; Ahn, S.; Lee, B.H. Zebrafish as a screening model for testing the permeability of blood–brain barrier to small molecules. Zebrafish 2017, 14, 322–330. [Google Scholar] [CrossRef]
- Lawrence, C. The husbandry of zebrafish (Danio rerio): A review. Aquaculture 2007, 269, 1–20. [Google Scholar] [CrossRef]
- Basnet, R.M.; Zizioli, D.; Taweedet, S.; Finazzi, D.; Memo, M. Zebrafish larvae as a behavioral model in neuropharmacology. Biomedicines 2019, 7, 23. [Google Scholar] [CrossRef]
- Tegelenbosch, R.A.; Noldus, L.P.; Richardson, M.K.; Ahmad, F. Zebrafish embryos and larvae in behavioural assays. Behaviour 2012, 149, 1241–1281. [Google Scholar] [CrossRef]
- Park, B.H. Novel Lactobacillus Buchneri and Use Thereof. KR Patent KR101917497B1, 9 November 2018. [Google Scholar]
- Park, B.H. Novel Lactobacillus Harbinensis and Use Thereof. KR Patent KR101889647B1, 20 August 2018. [Google Scholar]
- Park, B.H. Novel Saccharomycopsis Fibuligera and Use Thereof. KR Patent KR101876668B, 19 July 2018. [Google Scholar]
- Westerfield, M. The Zebrafish Book. A Guide for the Laboratory Use of Zebrafish (Danio rerio), 4th ed.; 2000; Available online: https://zfin.org/zf_info/zfbook/zfbk.html (accessed on 12 May 2023).
- Ellis, L.D.; Seibert, J.; Soanes, K.H. Distinct models of induced hyperactivity in zebrafish larvae. Brain Res. 2012, 1449, 46–59. [Google Scholar] [CrossRef]
- Arrant, A.E.; Schramm-Sapyta, N.L.; Kuhn, C.M. Use of the light/dark test for anxiety in adult and adolescent male rats. Behav. Brain Res. 2013, 256, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-S.; Ryu, J.-H.; Choi, T.-I.; Bae, Y.-K.; Lee, S.; Kang, H.J.; Kim, C.-H. Innate color preference of zebrafish and its use in behavioral analyses. Mol. Cells 2016, 39, 750. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Satou, C.; Higashijima, S.-I. V2a and V2b neurons are generated by the final divisions of pair-producing progenitors in the zebrafish spinal cord. Development 2008, 135, 3001–3005. [Google Scholar] [CrossRef] [PubMed]
- Muto, A.; Ohkura, M.; Abe, G.; Nakai, J.; Kawakami, K. Real-time visualization of neuronal activity during perception. Curr. Biol. 2013, 23, 307–311. [Google Scholar] [CrossRef]
- Li, F.; Long, Y.; Xie, J.; Ren, J.; Zhou, T.; Song, G.; Li, Q.; Cui, Z. Generation of GCaMP6s-Expressing Zebrafish to Monitor Spatiotemporal Dynamics of Calcium Signaling Elicited by Heat Stress. Int. J. Mol. Sci. 2021, 22, 5551. [Google Scholar] [CrossRef]
- Kim, S.S.; Lee, H.-Y.; Song, J.S.; Bae, M.-A.; Ahn, S. UPLC-MS/MS-based profiling of 31 neurochemicals in the mouse brain after treatment with the antidepressant nefazodone. Microchem. J. 2021, 169, 106580. [Google Scholar] [CrossRef]
- Afrikanova, T.; Serruys, A.-S.K.; Buenafe, O.E.; Clinckers, R.; Smolders, I.; de Witte, P.A.; Crawford, A.D.; Esguerra, C.V. Validation of the zebrafish pentylenetetrazol seizure model: Locomotor versus electrographic responses to antiepileptic drugs. PLoS ONE 2013, 8, e54166. [Google Scholar] [CrossRef]
- Ornell, F.; Valvassori, S.S.; Steckert, A.V.; Deroza, P.F.; Resende, W.R.; Varela, R.B.; Quevedo, J. Modafinil effects on behavior and oxidative damage parameters in brain of Wistar rats. Behav. Neurol. 2014, 2014, 1–7. [Google Scholar] [CrossRef]
- Rosniyana, A.; Hashifah, M.; Norin, S.S. The physico-chemical properties and nutritional composition of rice bran produced at different milling degrees of rice. J. Trop. Agric. Food Sci. 2007, 35, 99. [Google Scholar]
- Tuncel, N.B.; Yılmaz, N.; Kocabıyık, H.; Uygur, A. The effect of infrared stabilized rice bran substitution on B vitamins, minerals and phytic acid content of pan breads: Part II. J. Cereal Sci. 2014, 59, 162–166. [Google Scholar] [CrossRef]
- Kim, S.S.; Hwang, K.-S.; Yang, J.Y.; Chae, J.S.; Kim, G.R.; Kan, H.; Jung, M.H.; Lee, H.-Y.; Song, J.S.; Ahn, S. Neurochemical and behavioral analysis by acute exposure to bisphenol A in zebrafish larvae model. Chemosphere 2020, 239, 124751. [Google Scholar] [CrossRef] [PubMed]
- Akerboom, J.; Chen, T.-W.; Wardill, T.J.; Tian, L.; Marvin, J.S.; Mutlu, S.; Calderón, N.C.; Esposti, F.; Borghuis, B.G.; Sun, X.R. Optimization of a GCaMP calcium indicator for neural activity imaging. J. Neurosci. 2012, 32, 13819–13840. [Google Scholar] [CrossRef] [PubMed]
- Panula, P.; Sallinen, V.; Sundvik, M.; Kolehmainen, J.; Torkko, V.; Tiittula, A.; Moshnyakov, M.; Podlasz, P. Modulatory neurotransmitter systems and behavior: Towards zebrafish models of neurodegenerative diseases. Zebrafish 2006, 3, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Kacprzak, V.; Patel, N.A.; Riley, E.; Yu, L.; Yeh, J.-R.J.; Zhdanova, I.V. Dopaminergic control of anxiety in young and aged zebrafish. Pharmacol. Biochem. Behav. 2017, 157, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tu, X.; Li, Y.-W.; Chen, Q.-L.; Shen, Y.-J.; Liu, Z.-H. Tributyltin enhanced anxiety of adult male zebrafish through elevating cortisol level and disruption in serotonin, dopamine and gamma-aminobutyric acid neurotransmitter pathways. Ecotoxicol. Environ. Saf. 2020, 203, 111014. [Google Scholar] [CrossRef]
- Qiu, X.; Chen, C.; Shi, Y.; Chen, K.; Li, M.; Xu, H.; Wu, X.; Takai, Y.; Shimasaki, Y.; Oshima, Y. Persistent impact of amitriptyline on the behavior, brain neurotransmitter, and transcriptional profile of zebrafish (Danio rerio). Aquat. Toxicol. 2022, 245, 106129. [Google Scholar] [CrossRef]
- Earl, R.; Thomas, P.R. Opportunities in the Nutrition and Food Sciences: Research Challenges and the Next Generation of Investigators; National Academies Press: Washington, DC, USA, 1994. [Google Scholar]
- Alongi, M.; Anese, M. Re-thinking functional food development through a holistic approach. J. Funct. Foods 2021, 81, 104466. [Google Scholar] [CrossRef]
- Pallas, L. Rice processing: Beyond the farm gate. In Encyclopedia of Food Grains, 2nd ed.; Academic Press: Oxford, UK, 2016; pp. 446–452. [Google Scholar]
- Moon, S.-H.; Chang, H.-C. Rice bran fermentation using Lactiplantibacillus plantarum EM as a starter and the potential of the fermented rice bran as a functional food. Foods 2021, 10, 978. [Google Scholar] [CrossRef]
- Ryan, E.P.; Heuberger, A.L.; Weir, T.L.; Barnett, B.; Broeckling, C.D.; Prenni, J.E. Rice bran fermented with Saccharomyces boulardii generates novel metabolite profiles with bioactivity. J. Agric. Food Chem. 2011, 59, 1862–1870. [Google Scholar] [CrossRef]
- Cheigh, H.S.; Park, K.Y.; Lee, C. Biochemical, microbiological, and nutritional aspects of kimchi (Korean fermented vegetable products). Crit. Rev. Food Sci. Nutr. 1994, 34, 175–203. [Google Scholar] [CrossRef]
- Bell, V.; Ferrão, J.; Fernandes, T. Nutritional guidelines and fermented food frameworks. Foods 2017, 6, 65. [Google Scholar] [CrossRef] [PubMed]
- Behera, S.S.; El Sheikha, A.F.; Hammami, R.; Kumar, A. Traditionally fermented pickles: How the microbial diversity associated with their nutritional and health benefits? J. Funct. Foods 2020, 70, 103971. [Google Scholar] [CrossRef]
- Park, B.H. Novel Pichia Kudriavzevii and Use Thereof. KR Patent KR101876669B1, 9 July 2018. [Google Scholar]
- Xiao, W.; Wang, R.-S.; Handy, D.E.; Loscalzo, J. NAD (H) and NADP (H) redox couples and cellular energy metabolism. Antioxid. Redox Signal. 2018, 28, 251–272. [Google Scholar] [CrossRef] [PubMed]
- Gasperi, V.; Sibilano, M.; Savini, I.; Catani, M.V. Niacin in the central nervous system: An update of biological aspects and clinical applications. Int. J. Mol. Sci. 2019, 20, 974. [Google Scholar] [CrossRef]
- Fukushima, T. Niacin metabolism and Parkinson’s disease. Environ. Health Prev. Med. 2005, 10, 3–8. [Google Scholar] [CrossRef]
- Wakade, C.; Chong, R.; Bradley, E.; Morgan, J.C. Low-dose niacin supplementation modulates GPR109A, niacin index and ameliorates Parkinson’s disease symptoms without side effects. Clin. Case Rep. 2015, 3, 635. [Google Scholar] [CrossRef]
- Seamon, M.; Purohit, S.; Giri, B.; Baban, B.; Morgan, J.; Chong, R.; Wakade, C. Niacin for Parkinson’s disease. Clin. Exp. Neuroimmunol. 2020, 11, 47–56. [Google Scholar] [CrossRef]
- Wakade, C.; Chong, R. A novel treatment target for Parkinson’s disease. J. Neurol. Sci. 2014, 347, 34–38. [Google Scholar] [CrossRef]
- Chong, R.; Wakade, C.; Seamon, M.; Giri, B.; Morgan, J.; Purohit, S. Niacin Enhancement for Parkinson’s Disease: An Effectiveness Trial. Front. Aging Neurosci. 2021, 13, 667032. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, W.; Yang, J.; Wang, F.; Sima, Y.; Zhong, Z.-m.; Wang, H.; Hu, L.-F.; Liu, C.-F. Parkinson’s disease-like motor and non-motor symptoms in rotenone-treated zebrafish. Neurotoxicology 2017, 58, 103–109. [Google Scholar] [CrossRef]
- Shontz, E.; Souders, C.; Schmidt, J.; Martyniuk, C. Domperidone upregulates dopamine receptor expression and stimulates locomotor activity in larval zebrafish (Danio rerio). Genes Brain Behav. 2018, 17, e12460. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chae, J.S.; Kim, S.S.; Hwang, K.-S.; Kan, H.; Yang, J.Y.; Lee, B.; Shin, D.-S.; Park, B.; Bae, M.A. Neurobehavioral Effects of Fermented Rice Bran Extract in Zebrafish Larvae Model. Fermentation 2023, 9, 479. https://doi.org/10.3390/fermentation9050479
Chae JS, Kim SS, Hwang K-S, Kan H, Yang JY, Lee B, Shin D-S, Park B, Bae MA. Neurobehavioral Effects of Fermented Rice Bran Extract in Zebrafish Larvae Model. Fermentation. 2023; 9(5):479. https://doi.org/10.3390/fermentation9050479
Chicago/Turabian StyleChae, Jin Sil, Seong Soon Kim, Kyu-Seok Hwang, Hyemin Kan, Jung Yoon Yang, Byunghoi Lee, Dae-Seop Shin, Byounghee Park, and Myung Ae Bae. 2023. "Neurobehavioral Effects of Fermented Rice Bran Extract in Zebrafish Larvae Model" Fermentation 9, no. 5: 479. https://doi.org/10.3390/fermentation9050479