
In Vitro Characterization of Limosilactobacillus reuteri Lac Ib01 (OL468126.1) Isolated from Traditional Sheep Dry Sausage and Evaluation of the Activity of Arthrospira platensis or Phycocyanin on Its Growth-Promoting Ability
 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Isolation and Identification of Limosilactobacillus reuteri
2.2. Characteristic Features of Isolated Probiotic
Acid and Bile Tolerance Tests
2.3. Antimicrobial Assay
2.3.1. Agar Well Diffusion Method
2.3.2. Antimicrobial Agent Characterization
2.4. Antibacterial Activity of Reuterin Extracts
2.5. Antiproliferative Assay
2.5.1. Cell Lines and Culture Medium
2.5.2. Cell Viability Screening Assays
2.6. Limosilactobacillus reuteri Adhesion Assay
2.7. Growth Kinetics of L. reuteri Supplemented with Arthrospira platensis or Phycocyanin
2.8. Statistical Analysis
3. Results
3.1. Metabolic Profile and Molecular Characterization of Limosilactobacillus reuteri (OL468126.1)
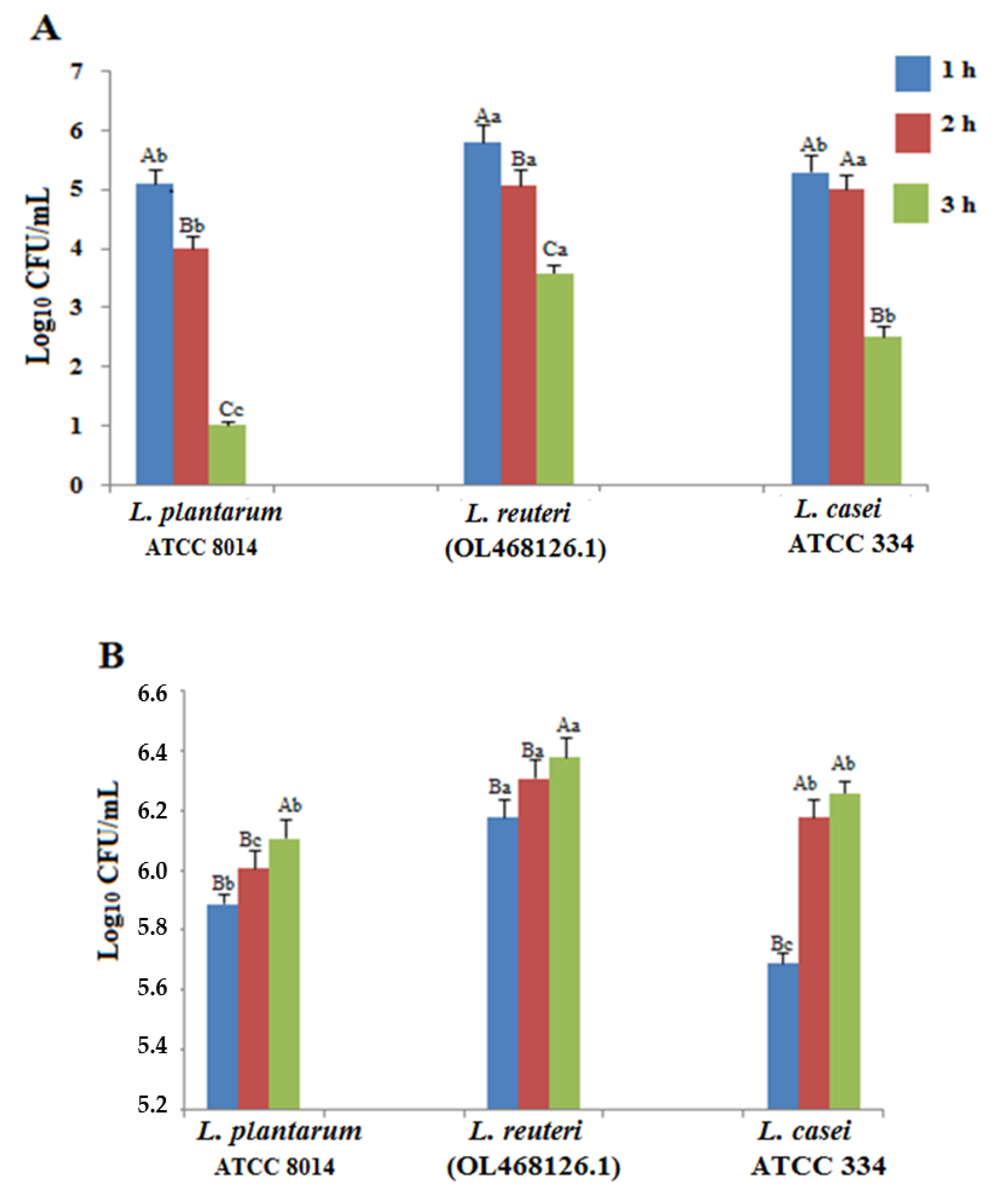
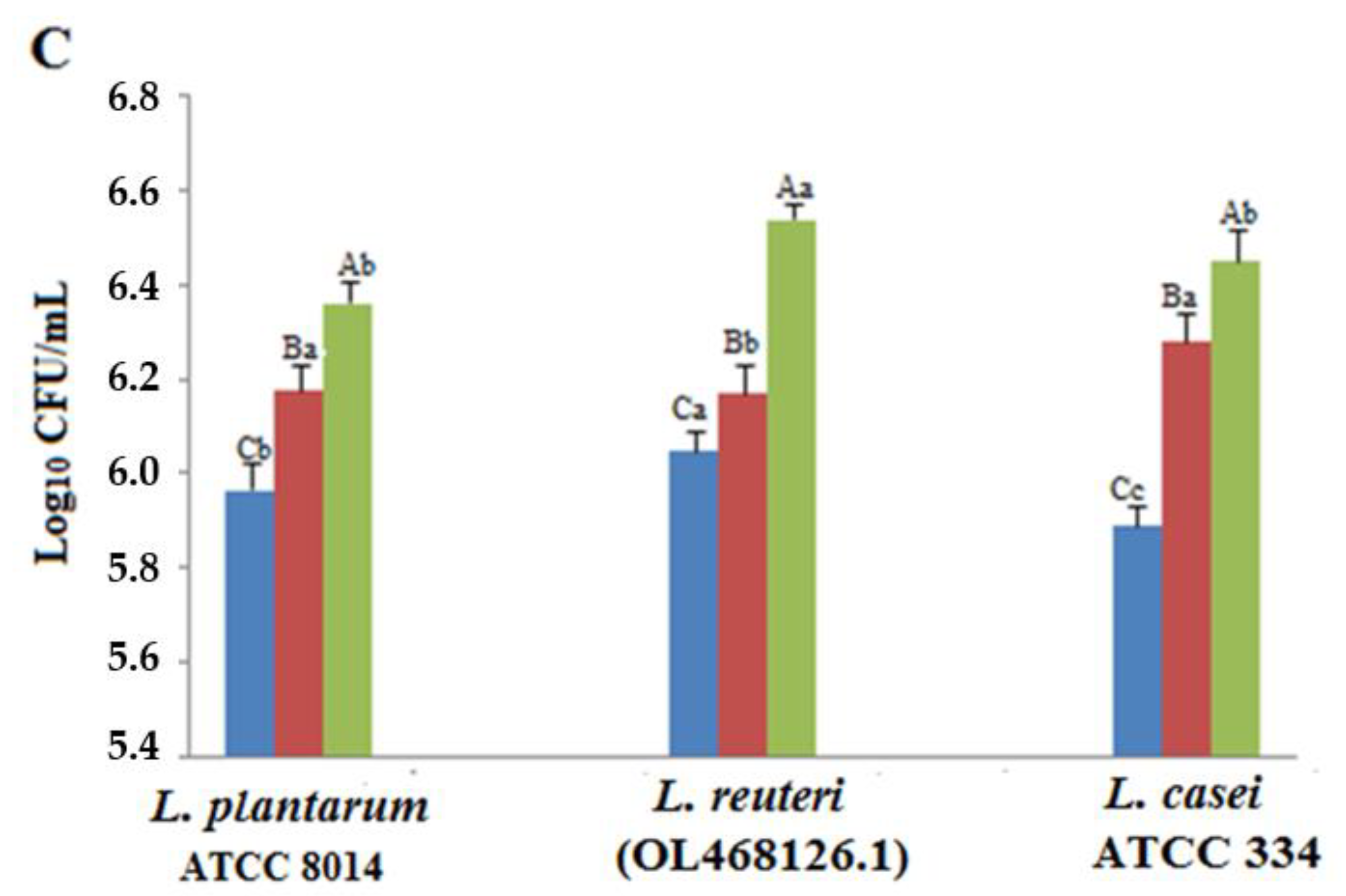
3.2. Acid and Bile Tolerances
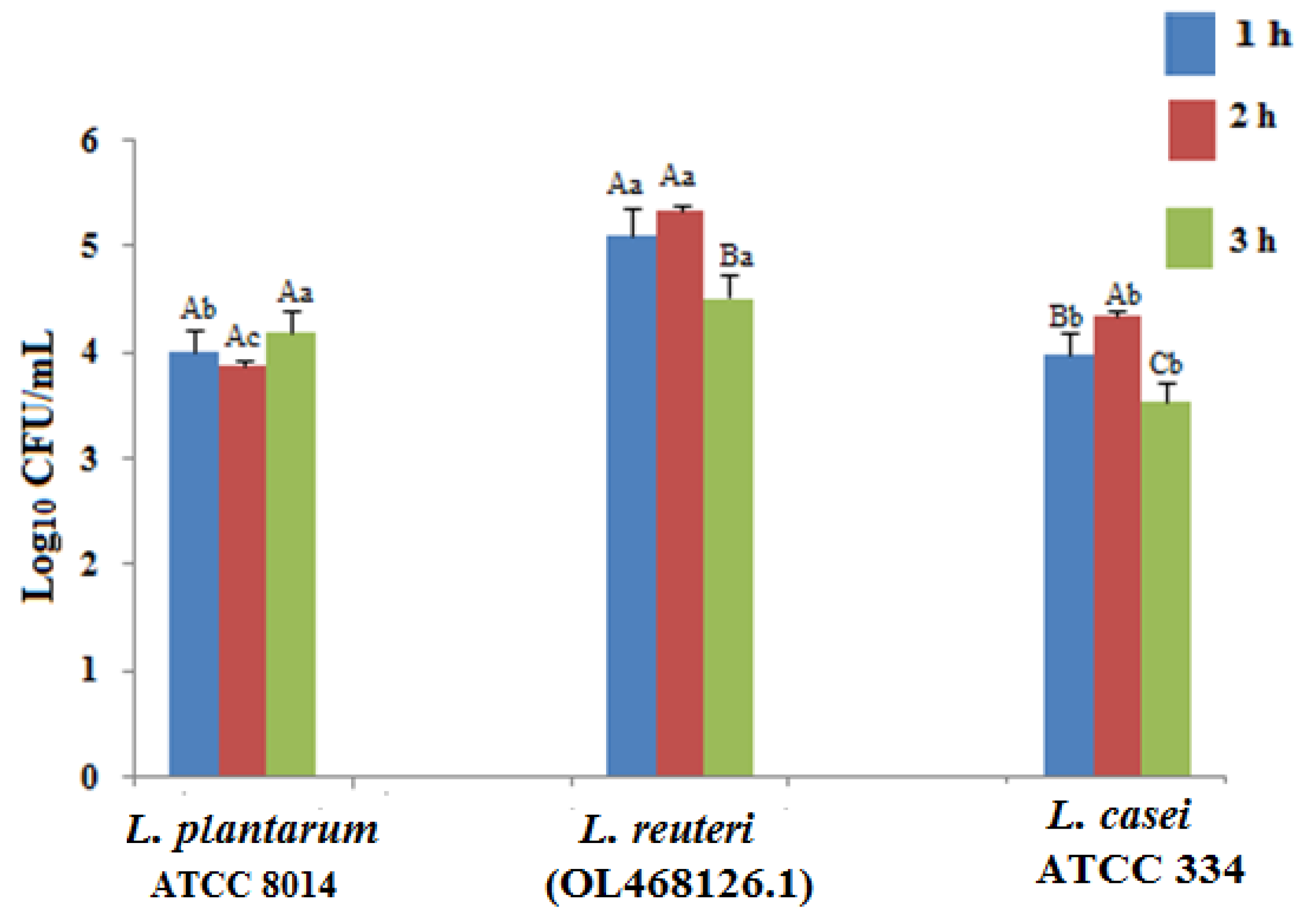
3.3. Antimicrobial Characterization of Limosilactobacillus reuteri (OL468126.1)
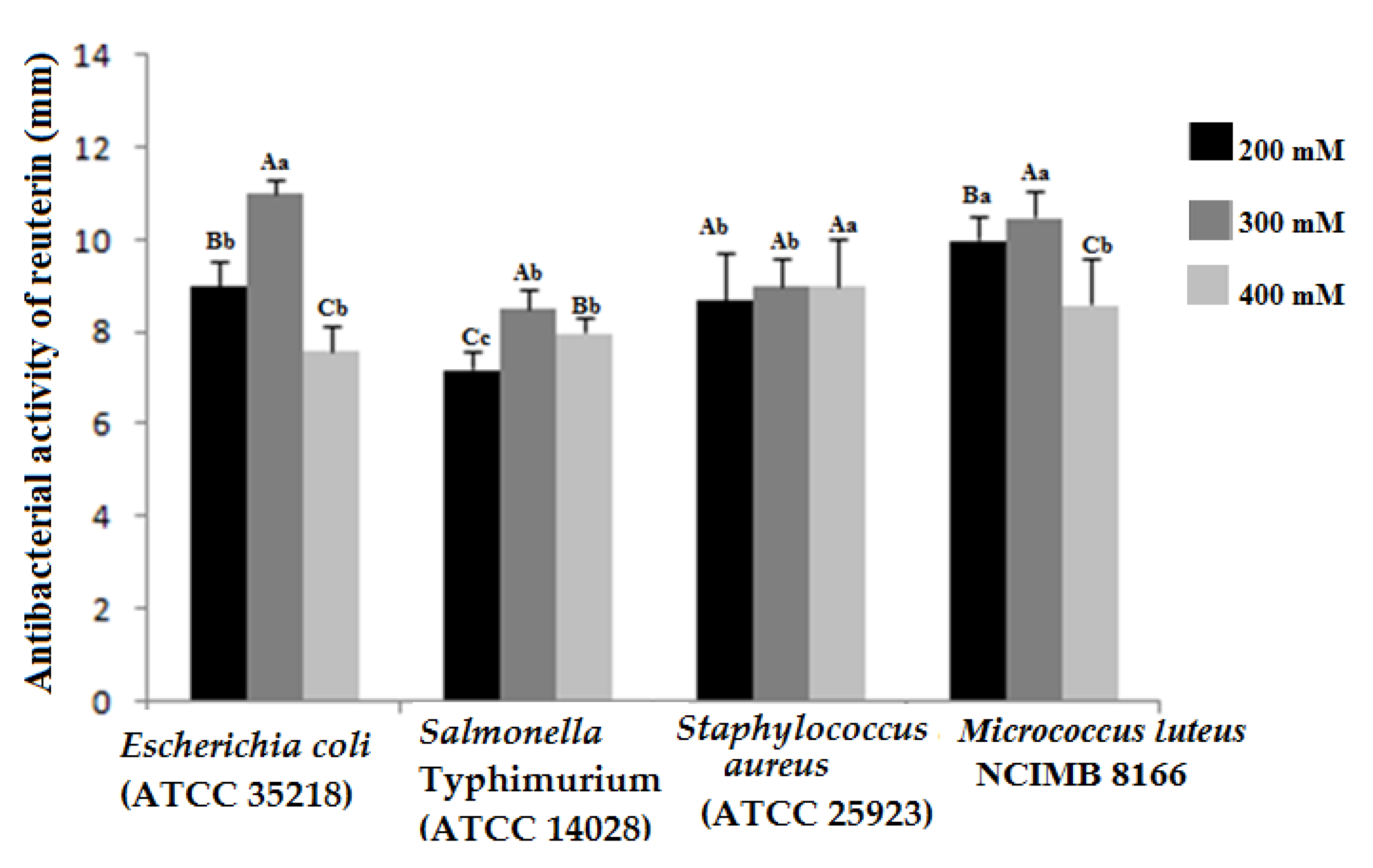
3.4. Antibacterial Activity of Reuterin Extracts
3.5. Adhesion Ability and Antiproliferative Activity
3.6. Growth Promotion of Isolated Limosilactobacillus reuteri with A. platensis or Phycocyanin
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Topolska, K.; Florkiewicz, A.; Filipiak-Florkiewicz, A. Functional Food—Consumer Motivations and Expectations. Int. J. Environ. Res. Public Health 2021, 18, 5327. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Rodrigues, C.F.; Stojanović-Radić, Z.; Dimitrijević, M.; Aleksić, A.; Neffe-Skocińska, K.; Zielińska, D.; Kołożyn-Krajewska, D.; Salehi, B.; Milton Prabu, S.; et al. Probiotics: Versatile Bioactive Components in Promoting Human Health. Medicina 2020, 56, 433. [Google Scholar] [CrossRef]
- Torres-Maravilla, E.; Reyes-Pavón, D.; Benítez-Cabello, A.; González-Vázquez, R.; Ramírez-Chamorro, L.M.; Langella, P.; Bermúdez-Humarán, L.G. Strategies for the Identification and Assessment of Bacterial Strains with Specific Probiotic Traits. Microorganisms 2022, 10, 1389. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh Behbahani, B.; Noshad, M.; Falah, F. Inhibition of Escherichia coli Adhesion to Human Intestinal Caco-2 Cells by Probiotic Candidate Lactobacillus Plantarum Strain L15. Microb. Pathog. 2019, 136, 103677. [Google Scholar] [CrossRef] [PubMed]
- Holzapfel, W.H.; Haberer, P.; Geisen, R.; Björkroth, J.; Schillinger, U. Taxonomy and Important Features of Probiotic Microorganisms in Food and Nutrition. Am. J. Clin. Nutr. 2001, 73, 365s–373s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darmastuti, A.; Hasan, P.N.; Wikandari, R.; Utami, T.; Rahayu, E.S.; Suroto, D.A. Adhesion Properties of Lactobacillus Plantarum Dad-13 and Lactobacillus Plantarum Mut-7 on Sprague Dawley Rat Intestine. Microorganisms 2021, 9, 2336. [Google Scholar] [CrossRef]
- Reid, G.; Gadir, A.A.; Dhir, R. Probiotics: Reiterating What They Are and What They Are Not. Front. Microbiol. 2019, 10, 424. [Google Scholar] [CrossRef] [Green Version]
- Gibson, G.R.; Roberfroid, M.B. Dietary Modulation of the Human Colonic Microbiota: Introducing the Concept of Prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef]
- Beheshtipour, H.; Mortazavian, A.M.; Mohammadi, R.; Sohrabvandi, S.; Khosravi-Darani, K. Supplementation of Spirulina Platensis and Chlorella Vulgaris Algae into Probiotic Fermented Milks: Algae Addition into Probiotic Fermented Milks…. Compr. Rev. Food Sci. Food Saf. 2013, 12, 144–154. [Google Scholar] [CrossRef]
- Molnár, N.; Gyenis, B.; Varga, L. Influence of a Powdered Spirulina Platensis Biomass on Acid Production of Lactococci in Milk. Milchwissenschaft 2005, 60, 380–382. [Google Scholar]
- Sommella, E.; Conte, G.; Salviati, E.; Pepe, G.; Bertamino, A.; Ostacolo, C.; Sansone, F.; Prete, F.; Aquino, R.; Campiglia, P. Fast Profiling of Natural Pigments in Different Spirulina (Arthrospira Platensis) Dietary Supplements by DI-FT-ICR and Evaluation of Their Antioxidant Potential by Pre-Column DPPH-UHPLC Assay. Molecules 2018, 23, 1152. [Google Scholar] [CrossRef] [Green Version]
- Safari, R.; Raftani Amiri, Z.; Esmaeilzadeh Kenari, R. Antioxidant and Antibacterial Activities of C-Phycocyanin from Common Name Spirulina Platensis. Iran. J. Fish. Sci. 2019, 19, 1911–1927. [Google Scholar] [CrossRef]
- Wang, X.-Q.; Li, L.-N.; Chang, W.-R.; Zhang, J.-P.; Gui, L.-L.; Guo, B.-J.; Liang, D.-C. Structure of C-Phycocyanin from Spirulina Platensis at 2.2 AÊ Resolution: A Novel Monoclinic Crystal Form for Phycobiliproteins in Phycobilisomes. Acta Crystallogr. Sect. D: Biol. Crystallogr. 2001, 57, 784–792. [Google Scholar] [CrossRef]
- Abdel-Moneim, A.-M.E.; El-Saadony, M.T.; Shehata, A.M.; Saad, A.M.; Aldhumri, S.A.; Ouda, S.M.; Mesalam, N.M. Antioxidant and Antimicrobial Activities of Spirulina Platensis Extracts and Biogenic Selenium Nanoparticles against Selected Pathogenic Bacteria and Fungi. Saudi J. Biol. Sci. 2021, 29, 1197–1209. [Google Scholar] [CrossRef] [PubMed]
- Barros de Medeiros, V.P.; Salgaço, M.K.; Pimentel, T.C.; Rodrigues da Silva, T.C.; Sartoratto, A.; dos Lima, M.S.; da Sassi, C.F.C.; Mesa, V.; Magnani, M.; Sivieri, K. Spirulina Platensis Biomass Enhances the Proliferation Rate of Lactobacillus Acidophilus 5 (La-5) and Combined with La-5 Impact the Gut Microbiota of Medium-Age Healthy Individuals through an in Vitro Gut Microbiome Model. Food Res. Int. 2022, 154, 110880. [Google Scholar] [CrossRef] [PubMed]
- Shimamatsu, H. Mass Production of Spirulina, an Edible Microalga. Hydrobiologia 2004, 512, 39–44. [Google Scholar] [CrossRef]
- Çelekli, A.; Alslibi, Z.A.; Bozkurt, H. Boosting Effects of Spirulina Platensis, Whey Protein, and Probiotics on the Growth of Microflora and the Nutritional Value of Ayran. Eng. Rep. 2020, 2, e12235. [Google Scholar] [CrossRef]
- Barkallah, M.; Dammak, M.; Louati, I.; Hentati, F.; Hadrich, B.; Mechichi, T.; Ayadi, M.A.; Fendri, I.; Attia, H.; Abdelkafi, S. Effect of Spirulina Platensis Fortification on Physicochemical, Textural, Antioxidant and Sensory Properties of Yogurt during Fermentation and Storage. LWT 2017, 84, 323–330. [Google Scholar] [CrossRef]
- Çelekli, A.; Alslibi, Z.A.; Bozkurt, H. Influence of Incorporated Spirulina Platensis on the Growth of Microflora and Physicochemical Properties of Ayran as a Functional Food. Algal Res. 2019, 44, 101710. [Google Scholar] [CrossRef]
- Patel, P.; Jethani, H.; Radha, C.; Vijayendra, S.V.N.; Mudliar, S.N.; Sarada, R.; Chauhan, V.S. Development of a Carotenoid Enriched Probiotic Yogurt from Fresh Biomass of Spirulina and Its Characterization. J. Food Sci. Technol. 2019, 56, 3721–3731. [Google Scholar] [CrossRef]
- Martelli, F.; Alinovi, M.; Bernini, V.; Gatti, M.; Bancalari, E. Arthrospira Platensis as Natural Fermentation Booster for Milk and Soy Fermented Beverages. Foods 2020, 9, 350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plaza, M.; Herrero, M.; Cifuentes, A.; Ibáñez, E. Innovative Natural Functional Ingredients from Microalgae. J. Agric. Food Chem. 2009, 57, 7159–7170. [Google Scholar] [CrossRef] [PubMed]
- Chiong, T.; Acquah, C.; Lau, S.Y.; Khor, E.H.; Danquah, M.K. Microalgal-Based Protein By-Products: Extraction, Purification, and Applications. In Protein Byproducts; Elsevier: Amsterdam, The Netherlands, 2016; pp. 213–234. [Google Scholar] [CrossRef]
- Xie, Y.; Li, W.; Zhu, L.; Zhai, S.; Qin, S.; Du, Z. Effects of Phycocyanin in Modulating the Intestinal Microbiota of Mice. Microbiol. Open 2019, 8, e00825. [Google Scholar] [CrossRef] [Green Version]
- The Human Microbiome Project Consortium. Structure, Function and Diversity of the Healthy Human Microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heilig, H.G.H.J.; Zoetendal, E.G.; Vaughan, E.E.; Marteau, P.; Akkermans, A.D.L.; de Vos, W.M. Molecular Diversity of Lactobacillus Spp. and Other Lactic Acid Bacteria in the Human Intestine as Determined by Specific Amplification of 16S Ribosomal DNA. Appl. Environ. Microbiol. 2002, 68, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Schumann, P. E. Stackebrandt and M. Goodfellow (Editors), Nucleic Acid Techniques in Bacterial Systematics (Modern Microbiological Methods). XXIX + 329 S., 46 Abb., 28 Tab. Chichester—New York—Brisbane—Toronto—Singapore 1991. John Wiley & Sons. $ 55.00. ISBN: 0-471-92906-9. J. Basic Microbiol. 1991, 31, 479–480. [Google Scholar] [CrossRef]
- Kaboré, D.; Sawadogo-Lingani, H.; Dicko, M.H.; Diawara, B.; Jakobsen, M. Acid Resistance, Bile Tolerance and Antimicrobial Properties of Dominant Lactic Acid Bacteria Isolated from Traditional Maari Baobab Seeds Fermented Condiment. Afr. J. Biotechnol. 2012, 11, 1197–1205. [Google Scholar] [CrossRef]
- Klingberg, T.D.; Axelsson, L.; Naterstad, K.; Elsser, D.; Budde, B.B. Identification of Potential Probiotic Starter Cultures for Scandinavian-Type Fermented Sausages. Int. J. Food Microbiol. 2005, 105, 419–431. [Google Scholar] [CrossRef]
- Lamari, F.; Mahdhi, A.; Chakroun, I.; Esteban, M.A.; Mazurais, D.; Amina, B.; Gatesoupe, F.-J. Interactions between Candidate Probiotics and the Immune and Antioxidative Responses of European Sea Bass (Dicentrarchus Labrax) Larvae. J. Fish Dis. 2016, 39, 1421–1432. [Google Scholar] [CrossRef]
- Shokryazdan, P.; Sieo, C.C.; Kalavathy, R.; Liang, J.B.; Alitheen, N.B.; Faseleh Jahromi, M.; Ho, Y.W. Probiotic Potential of Lactobacillus Strains with Antimicrobial Activity against Some Human Pathogenic Strains. BioMed Res. Int. 2014, 2014, 927268. [Google Scholar] [CrossRef] [Green Version]
- Doleyres, Y.; Beck, P.; Vollenweider, S.; Lacroix, C. Production of 3-Hydroxypropionaldehyde Using a Two-Step Process with Lactobacillus Reuteri. Appl. Microbiol. Biotechnol. 2005, 68, 467–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Verdenelli, M.C.; Ghelfi, F.; Silvi, S.; Orpianesi, C.; Cecchini, C.; Cresci, A. Probiotic Properties of Lactobacillus Rhamnosus and Lactobacillus Paracasei Isolated from Human Faeces. Eur. J. Nutr. 2009, 48, 355–363. [Google Scholar] [CrossRef]
- Bhowmik, D.; Dubey, J.; Mehra, S. Probiotic Efficiency of Spirulina Platensis—Stimulating Growth of Lactic Acid Bacteria. World J. Dairy Food Sci. 2009, 4, 160–163. [Google Scholar]
- De Filippis, F.; Pasolli, E.; Ercolini, D. The Food-Gut Axis: Lactic Acid Bacteria and Their Link to Food, the Gut Microbiome and Human Health. FEMS Microbiol. Rev. 2020, 44, 454–489. [Google Scholar] [CrossRef]
- Zommiti, M.; Feuilloley, M.G.J.; Connil, N. Update of Probiotics in Human World: A Nonstop Source of Benefactions till the End of Time. Microorganisms 2020, 8, 1907. [Google Scholar] [CrossRef] [PubMed]
- Mu, Q.; Tavella, V.J.; Luo, X.M. Role of Lactobacillus Reuteri in Human Health and Diseases. Front. Microbiol. 2018, 9, 757. [Google Scholar] [CrossRef] [Green Version]
- Abuqwider, J.; Altamimi, M.; Mauriello, G. Limosilactobacillus Reuteri in Health and Disease. Microorganisms 2022, 10, 522. [Google Scholar] [CrossRef] [PubMed]
- Sulemankhil, I.; Parent, M.; Jones, M.L.; Feng, Z.; Labbé, A.; Prakash, S. In Vitro and in Vivo Characterization and Strain Safety of Lactobacillus Reuteri NCIMB 30253 for Probiotic Applications. Can. J. Microbiol. 2012, 58, 776–787. [Google Scholar] [CrossRef]
- Hou, C.; Zeng, X.; Yang, F.; Liu, H.; Qiao, S. Study and Use of the Probiotic Lactobacillus Reuteri in Pigs: A Review. J. Anim. Sci. Biotechnol. 2015, 6, 14. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Ali Esmail, G.; Fahad Alzeer, A.; Valan Arasu, M.; Vijayaraghavan, P.; Choon Choi, K.; Abdullah Al-Dhabi, N. Probiotic Characteristics of Lactobacillus Strains Isolated from Cheese and Their Antibacterial Properties against Gastrointestinal Tract Pathogens. Saudi J. Biol. Sci. 2020, 27, 3505–3513. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Puniya, A.K.; Singh, K. Effect of Alginate Concentrations on Survival of Microencapsulated Lactobacillus Casei NCDC-298. Int. Dairy J. 2006, 16, 1190–1195. [Google Scholar] [CrossRef]
- Hofmann, A.F. Chemistry and Enterohepatic Circulation of Bile Acids. Hepatology 1984, 4 (Suppl. S2), 4S–14S. [Google Scholar] [CrossRef]
- Whitehead, K.; Versalovic, J.; Roos, S.; Britton, R.A. Genomic and Genetic Characterization of the Bile Stress Response of Probiotic Lactobacillus Reuteri ATCC 55730. Appl. Environ. Microbiol. 2008, 74, 1812–1819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunne, C.; O’Mahony, L.; Murphy, L.; Thornton, G.; Morrissey, D.; O’Halloran, S.; Feeney, M.; Flynn, S.; Fitzgerald, G.; Daly, C.; et al. In Vitro Selection Criteria for Probiotic Bacteria of Human Origin: Correlation with in Vivo Findings. Am. J. Clin. Nutr. 2001, 73, 386s–392s. [Google Scholar] [CrossRef] [Green Version]
- Agriopoulou, S.; Stamatelopoulou, E.; Sachadyn-Król, M.; Varzakas, T. Lactic Acid Bacteria as Antibacterial Agents to Extend the Shelf Life of Fresh and Minimally Processed Fruits and Vegetables: Quality and Safety Aspects. Microorganisms 2020, 8, 952. [Google Scholar] [CrossRef]
- Singh, T.P.; Kaur, G.; Kapila, S.; Malik, R.K. Antagonistic Activity of Lactobacillus Reuteri Strains on the Adhesion Characteristics of Selected Pathogens. Front. Microbiol. 2017, 8, 486. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.M.; Kim, J.-S.; Kim, H.-S.; Kim, Y.-Y.; Oh, J.-K.; Jung, H.-W.; Park, D.-S.; Bae, K.-H. Lactobacillus Reuteri AN417 Cell-Free Culture Supernatant as a Novel Antibacterial Agent Targeting Oral Pathogenic Bacteria. Sci. Rep. 2021, 11, 1631. [Google Scholar] [CrossRef]
- Makras, L.; De Vuyst, L. The in Vitro Inhibition of Gram-Negative Pathogenic Bacteria by Bifidobacteria Is Caused by the Production of Organic Acids. Int. Dairy J. 2006, 16, 1049–1057. [Google Scholar] [CrossRef]
- Ilavenil, S.; Vijayakumar, M.; Kim, D.H.; Valan Arasu, M.; Park, H.S.; Ravikumar, S.; Choi, K.C. Assessment of Probiotic, Antifungal and Cholesterol Lowering Properties of Pediococcus Pentosaceus KCC-23 Isolated from Italian Ryegrass: Properties of P. Pentosaceus from Ryegrass. J. Sci. Food Agric. 2016, 96, 593–601. [Google Scholar] [CrossRef]
- Kang, M.-S.; Oh, J.-S.; Lee, H.-C.; Lim, H.-S.; Lee, S.-W.; Yang, K.-H.; Choi, N.-K.; Kim, S.-M. Inhibitory Effect of Lactobacillus Reuteri on Periodontopathic and Cariogenic Bacteria. J. Microbiol. 2011, 49, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Cleusix, V.; Lacroix, C.; Vollenweider, S.; Duboux, M.; Le Blay, G. Inhibitory Activity Spectrum of Reuterin Produced by Lactobacillus Reuteri against Intestinal Bacteria. BMC Microbiol. 2007, 7, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asare, P.T.; Zurfluh, K.; Greppi, A.; Lynch, D.; Schwab, C.; Stephan, R.; Lacroix, C. Reuterin Demonstrates Potent Antimicrobial Activity against a Broad Panel of Human and Poultry Meat Campylobacter Spp. Isolates. Microorganisms 2020, 8, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Q.; Maddox, I.S.; Mutukumira, A.; Lee, S.J.; Shu, Q. The Effect of Cell Immobilization on the Antibacterial Activity of Lactobacillus Reuteri DPC16 Cells during Passage through a Simulated Gastrointestinal Tract System. World J. Microbiol. Biotechnol. 2012, 28, 3025–3037. [Google Scholar] [CrossRef]
- Schaefer, L.; Auchtung, T.A.; Hermans, K.E.; Whitehead, D.; Borhan, B.; Britton, R.A. The Antimicrobial Compound Reuterin (3-Hydroxypropionaldehyde) Induces Oxidative Stress via Interaction with Thiol Groups. Microbiology 2010, 156, 1589–1599. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Lu, Y.; Xie, J.; Fei, Y.; Zheng, G.; Wang, Z.; Liu, J.; Lv, L.; Ling, Z.; Berglund, B.; et al. Probiotic Gastrointestinal Transit and Colonization After Oral Administration: A Long Journey. Front. Cell. Infect. Microbiol. 2021, 11, 609722. [Google Scholar] [CrossRef]
- Fradinho, P.; Niccolai, A.; Soares, R.; Rodolfi, L.; Biondi, N.; Tredici, M.R.; Sousa, I.; Raymundo, A. Effect of Arthrospira Platensis (Spirulina) Incorporation on the Rheological and Bioactive Properties of Gluten-Free Fresh Pasta. Algal Res. 2020, 45, 101743. [Google Scholar] [CrossRef]
- Wang, Y.; Tibbetts, S.; McGinn, P. Microalgae as Sources of High-Quality Protein for Human Food and Protein Supplements. Foods 2021, 10, 3002. [Google Scholar] [CrossRef]
- Varga, L.; Szigeti, J.; Kovács, R.; Földes, T.; Buti, S. Influence of a Spirulina Platensis Biomass on the Microflora of Fermented ABT Milks During Storage (R1). J. Dairy Sci. 2002, 85, 1031–1038. [Google Scholar] [CrossRef]
- Perković, L.; Djedović, E.; Vujović, T.; Baković, M.; Paradžik, T.; Čož-Rakovac, R. Biotechnological Enhancement of Probiotics through Co-Cultivation with Algae: Future or a Trend? Mar. Drugs 2022, 20, 142. [Google Scholar] [CrossRef]
- Bhalamurugan, G.L.; Valerie, O.; Mark, L. Valuable Bioproducts Obtained from Microalgal Biomass and Their Commercial Applications: A Review. Environ. Eng. Res. 2018, 23, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Niccolai, A.; Shannon, E.; Abu-Ghannam, N.; Biondi, N.; Rodolfi, L.; Tredici, M.R. Lactic Acid Fermentation of Arthrospira Platensis (Spirulina) Biomass for Probiotic-Based Products. J. Appl. Phycol. 2019, 31, 1077–1083. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Lu, L.; Liu, B.; Qin, S. Effects of Phycocyanin on Pulmonary and Gut Microbiota in a Radiation-Induced Pulmonary Fibrosis Model. Biomed. Pharmacother. 2020, 132, 110826. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tube | Biochemical Tests | Results |
|---|---|---|
| 0 | Temoin | - |
| 1 | Glycerol | - |
| 2 | Erythritol | - |
| 3 | D-Arabinose | - |
| 4 | L-Arabinose | + |
| 5 | D-Ribose | + |
| 6 | D-Xylose | + |
| 7 | L-Xylose | - |
| 8 | D-Adonitol | - |
| 9 | Methyl-b-D-xylopyranoside | - |
| 10 | D-Galactose | + |
| 11 | D-Glucose | + |
| 12 | D-Fructose | - |
| 13 | D-Mannose | - |
| 14 | L-Sorbose | - |
| 15 | L-Rhamnose | - |
| 16 | Dulcitol | - |
| 17 | Inositol | - |
| 18 | D-Mannitol | - |
| 19 | D-Sorbitol | - |
| 20 | Methyl-a-D-mannopyroside | - |
| 21 | Methyl-a-D-glucopyranoside | - |
| 22 | N-Acetylglucosamine | - |
| 23 | Amygdalin | - |
| 24 | Arbutin | - |
| 25 | Esculinferriccitrate | - |
| 26 | Salicin | - |
| 27 | D-Cellobiose | - |
| 28 | D-Maltose | + |
| 29 | D-Lactose | + |
| 30 | D-Melibiose | + |
| 31 | D-Saccharose | + |
| 32 | D-Trehalose | - |
| 33 | Inulin | - |
| 34 | D-Melezitose | - |
| 35 | D-Raffinose | + |
| 36 | Amidon | - |
| 37 | Glycogen | - |
| 38 | Xylitol | - |
| 39 | Gentibiose | - |
| 40 | D-Turanose | - |
| 41 | D-Lyxose | - |
| 42 | D-Tagatose | - |
| 43 | D-Fucose | - |
| 44 | L-Fucose | - |
| 45 | D-Arabitol | - |
| 46 | L-Arabitol | - |
| 47 | Potassiumgluconate | ± |
| 48 | Potassium2-ketogluconate | - |
| 49 | Potassium5-ketogluconate | - |
| Strains | Escherichia coli (ATCC 35218) | Salmonella Typhimurium (ATCC 14028) | Pseudomonas aeruginosa (ATCC 27853) | Staphylococcus aureus (ATCC 25923) | Micrococcus luteus NCIMB 8166 | |
|---|---|---|---|---|---|---|
| Treatment | ||||||
| CFCS: control | 12.3 ± 0.6 Ba | 11.6 ± 1.0 Ca | - | 10.76 ± 1.2 Ca | 13.3 ± 0.6 Aa | |
| Trypsin | 10.6 ± 0.5 Ab | 8.3 ± 0.6 Cb | - | 9 ± 1 Cb | 10.6 ± 1 Bb | |
| Catalase | 9.5 ± 1 Ab | 8 ± 1.02 Bb | - | 9 ± 0.6 Ab | 8 ± 1.2 Bc | |
| pH = 6.5 | 8 ± 0.0 Ac | 7 ± 0.9 Bc | - | 8.3 ± 1.3 Ab | 7.2 ± 1.1 Bc | |
| Strain | Antiproliferative Activity (%) | Adhesion (%) | ||
|---|---|---|---|---|
| HCT-116 | Hep-2 | HCT-116 | Hep-2 | |
| Lac Ib01 | 60.3 ± 2.1 Ab | 57.06 ± 2.8 Aa | 73.12 ± 3.02 Ab | 60.7 ± 3.02 Aa |
| L. casei ATCC 334 | 76.25 ± 0.9 Aa | 40.18 ± 1.4 Bb | 78.08 ± 1.8 Ab | 53.64 ± 2.1 Bb |
| L. plantarum ATCC 8014 | 71.3 ± 1.1 Aa | 50.2 ± 2.2 Ba | 80.1 ± 2.5 Aa | 42.15 ± 2.8 Bc |
| L. reuteri OL468126.1 | L. casei ATCC 334 | L. plantarum ATCC 8014 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 h | 24 h | 48 h | 72 h | 0 h | 24 h | 48 h | 72 h | 0 h | 24 h | 48 h | 72 h | ||
| Control | 3.51 ± 0.25 Ca | 6.42 ± 0.3 Bc | 8.41± 1.5 Ac | 7.05 ± 1.12 Bc | 3.6 ± 0.52 Ca | 7.02 ± 0.2Bc | 9.1 ± 0.37 Ac | 8.79 ± 0.5 Ac | 4.2 ± 0.6 Ca | 7.02 ± 0.73 Bc | 9.4 ± 0.25 Ab | 8.15 ± 0.5 Ac | |
| Concentration of A. platensis (mg/mL) | 01 | 3.58 ± 0.58 Ca | 7.18 ± 0.63 Bc | 8.7 ± 1.1 Ac | 8.21 ± 1.03 Ac | 3.82 ± 0.9 Ca | 7.53 ± 0.5 Bc | 8.6 ± 0.21 Ac | 8.01 ± 1.05 Ac | 3.9 ± 0.83 Ca | 7.6 ± 0.78 Bc | 9.12 ± 0.97 Ab | 9.05 ± 0.62 Ab |
| 04 | 3.7 ± 0.12 Ca | 7.9 ± 0.7 Bb | 9.92 ± 0.73 Ab | 11.0 ± 0.94 Aa | 3.7 ± 0.6 Ca | 8.23 ± 1.1 Bb | 10.33 ± 0.5 Aa | 10.8 ± 0.9 Aa | 4 ± 0.2 Ca | 8.04 ± 0.35Bb | 10.58 ± 1 Aa | 10.6 ± 0.4 Aa | |
| 08 | 3.52 ± 0.26 Ca | 9.17 ± 0.41 Ba | 12.0 ± 0.6 Aa | 8.12 ± 0.92 Bb | 3.47 ± 0.3 Ca | 11.0 ± 0.91 Ba | 13.1 ± 0.84 Aa | 10.21 ± 0.9 Aa | 3.8 ± 0.15 Ca | 10.5 ± 0.5 Ba | 12.6 ± 0.2 Aa | 10.12 ± 0.7 Ba | |
| Concentration of phycocyanin extract (mg/mL) | 01 | 3.81 ± 0.95 Ca | 6.78 ± 0.5 Bc | 8.1 ± 0.2 Ac | 7.1 ± 1.2 Ac | 3.65 ± 0.47 Ca | 7.3 ± 0.83 Bc | 9.51 ± 0.9 Ab | 8.32 ± 0.7 Ac | 3.6 ± 0.5 Ca | 6.58 ± 1.02 Bc | 8.67 ± 1.Ab | 7.9 ± 0.81 Ac |
| 04 | 3.57 ± 0.2 Ca | 7.5 ± 0.9 Bc | 8.62 ± 0.15 Ac | 8.54 ± 1.0Ac | 3.75 ± 0.9 Ca | 8.02 ± 0.1 Bb | 10 ± 1.06 Ab | 10.6 ± 0.2Aa | 4.1 ± 0.7 Ca | 7.05 ± 0.3 Bc | 9.1 ± 0.Ab | 9.37 ± 0.61 Ab | |
| 08 | 3.64 ± 0.7 Ca | 8.03 ± 0.21 Bb | 9.51 ± 0.33 Ab | 9.05 ± 0.6Ab | 3.56 ± 1.02 Ca | 9.7 ± 0.5 Bb | 11.06 ± 0.8 Aa | 9.5 ± 0.34 Bb | 3.81 ± 0.92 Ca | 8.9 ± 0.57 Bb | 10.84 ± 0.92 Aa | 9.25 ± 0.7 Ab | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chakroun, I.; Haddaji, N.; Fedhila, K.; Maatallah, M.; Mzoughi, R.; Chaabouni, Y.; Krichen, Y.; Bakhrouf, A. In Vitro Characterization of Limosilactobacillus reuteri Lac Ib01 (OL468126.1) Isolated from Traditional Sheep Dry Sausage and Evaluation of the Activity of Arthrospira platensis or Phycocyanin on Its Growth-Promoting Ability. Fermentation 2023, 9, 248. https://doi.org/10.3390/fermentation9030248
Chakroun I, Haddaji N, Fedhila K, Maatallah M, Mzoughi R, Chaabouni Y, Krichen Y, Bakhrouf A. In Vitro Characterization of Limosilactobacillus reuteri Lac Ib01 (OL468126.1) Isolated from Traditional Sheep Dry Sausage and Evaluation of the Activity of Arthrospira platensis or Phycocyanin on Its Growth-Promoting Ability. Fermentation. 2023; 9(3):248. https://doi.org/10.3390/fermentation9030248
Chicago/Turabian StyleChakroun, Ibtissem, Najla Haddaji, Kais Fedhila, Makaoui Maatallah, Ridha Mzoughi, Yassine Chaabouni, Youssef Krichen, and Amina Bakhrouf. 2023. "In Vitro Characterization of Limosilactobacillus reuteri Lac Ib01 (OL468126.1) Isolated from Traditional Sheep Dry Sausage and Evaluation of the Activity of Arthrospira platensis or Phycocyanin on Its Growth-Promoting Ability" Fermentation 9, no. 3: 248. https://doi.org/10.3390/fermentation9030248