
Brackish and Hypersaline Lakes as Potential Reservoir for Enzymes Involved in Decomposition of Organic Materials on Frescoes
 , , , , , and
, , , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Salt Requirements of Fungal Isolates and Qualitative Evaluation of Hydrolytic Enzymes
2.2. Taxonomic Identification
2.3. Establishing the Optimal Methodology for Selecting Lipase-Producing Strains
2.4. Evaluation of the Hydrolytic Decomposition of the Deposits on the Surface of the Painted Laboratory Models and Fragments of Frescoes
3. Results and Discussion
3.1. Taxonomy of the Fungal Strains
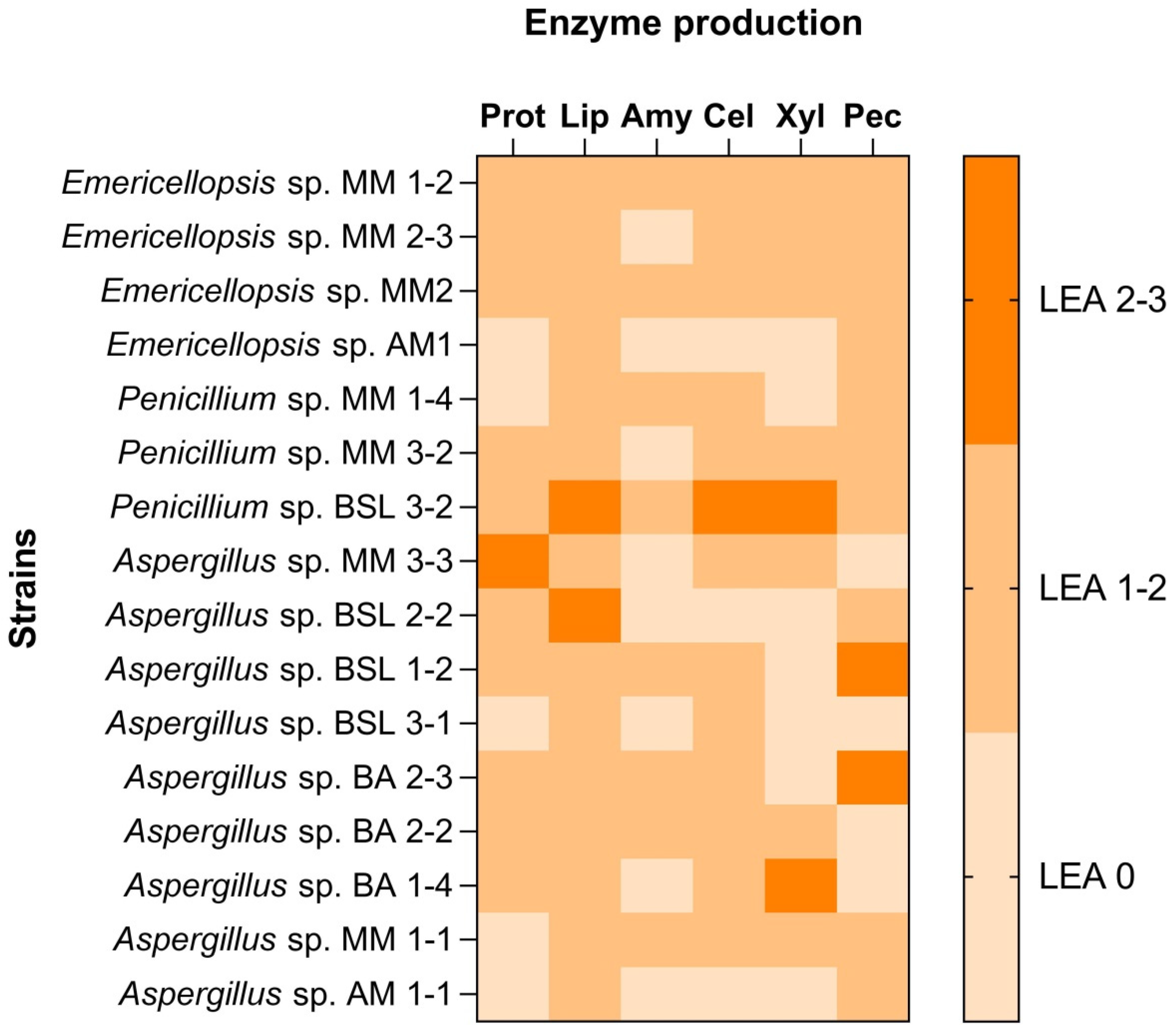
3.2. Salt Requirements of Fungal Isolates and Qualitative Evaluation of Hydrolytic Enzymes
3.3. Screening of Lipase-Producing Strains
3.4. Testing of the Best Lipase-Producing Strains for the Decomposition of Deposits on the Painted Laboratory Models
3.5. Spectroscopic and Imaging Methods also Revealed Changes on the Surface of Deposits
3.6. Testing of the Best Lipase-Producing Strains for the Decomposition of Deposits on the Fresco Fragments
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fabian, J.; Zlatanovic, S.; Mutz, M.; Premke, K. Fungal–Bacterial Dynamics and Their Contribution to Terrigenous Carbon Turnover in Relation to Organic Matter Quality. ISME J. 2017, 11, 415–425. [Google Scholar] [CrossRef]
- Zanne, A.E.; Abarenkov, K.; Afkhami, M.E.; Aguilar-Trigueros, C.A.; Bates, S.; Bhatnagar, J.M.; Busby, P.E.; Christian, N.; Cornwell, W.K.; Crowther, T.W.; et al. Fungal Functional Ecology: Bringing a Trait-based Approach to Plant-associated Fungi. Biol. Rev. 2020, 95, 409–433. [Google Scholar] [CrossRef]
- Geethanjali, P.A.; Jayashankar, M. A Review on Litter Decomposition by Soil Fungal Community. IOSR 2016, 11, 1–3. [Google Scholar] [CrossRef]
- Likar, M.; Grašič, M.; Stres, B.; Regvar, M.; Gaberščik, A. Original Leaf Colonisers Shape Fungal Decomposer Communities of Phragmites australis in Intermittent Habitats. JoF 2022, 8, 284. [Google Scholar] [CrossRef]
- Berbee, M.L.; James, T.Y.; Strullu-Derrien, C. Early Diverging Fungi: Diversity and Impact at the Dawn of Terrestrial Life. Annu. Rev. Microbiol. 2017, 71, 41–60. [Google Scholar] [CrossRef]
- Gadd, G.M. Fungal Biomineralization. Curr. Biol. 2021, 31, R1557–R1563. [Google Scholar] [CrossRef]
- Saranraj, P.; Stella, D. Fungal Amylase—A Review. Int. J. Microbiol. Res. 2013, 4, 203–211. [Google Scholar] [CrossRef]
- Ayansina, A.D.V.; Adelaja, A.O.; Mohammed, S.S.D. Characterization of Amylase from Some Aspergillus and Bacillus Species Associated with Cassava Waste Peels. AiM 2017, 7, 280–292. [Google Scholar] [CrossRef]
- Li, J.-X.; Zhang, F.; Jiang, D.-D.; Li, J.; Wang, F.-L.; Zhang, Z.; Wang, W.; Zhao, X.-Q. Diversity of Cellulase-Producing Filamentous Fungi from Tibet and Transcriptomic Analysis of a Superior Cellulase Producer Trichoderma harzianum LZ117. Front. Microbiol. 2020, 11, 1617. [Google Scholar] [CrossRef]
- Li, X.; Dilokpimol, A.; Kabel, M.A.; de Vries, R.P. Fungal Xylanolytic Enzymes: Diversity and Applications. Bioresour. Technol. 2022, 344, 126290. [Google Scholar] [CrossRef]
- Souza, P.M.; Bittencourt, M.L.A.; Caprara, C.C.; Freitas, M.; Almeida, R.P.C.; Silveira, D.; Fonseca, Y.M.; Ferreira Filho, E.X.; Pessoa Junior, A.; Magalhães, P.O. A Biotechnology Perspective of Fungal Proteases. Braz. J. Microbiol. 2015, 46, 337–346. [Google Scholar] [CrossRef]
- Haile, S.; Ayele, A. Pectinase from Microorganisms and Its Industrial Applications. Sci. World J. 2022, 2022, 1881305. [Google Scholar] [CrossRef]
- Kovacic, F.; Babic, N.; Krauss, U.; Jaeger, K.E. Classification of Lipolytic Enzymes from Bacteria. In Aerobic Utilization of Hydrocarbons, Oils, and Lipids; Rojo, F., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 255–289. ISBN 978-3-319-50417-9. [Google Scholar]
- Bracco, P.; van Midden, N.; Arango, E.; Torrelo, G.; Ferrario, V.; Gardossi, L.; Hanefeld, U. Bacillus subtilis Lipase A—Lipase or Esterase? Catalysts 2020, 10, 308. [Google Scholar] [CrossRef]
- Sayali, K.; Sadichha, P.; Surekha, S. Microbial Esterases: An Overview. Int. J. Curr. Microbiol. App. Sci. 2013, 2, 135–146. [Google Scholar]
- Faiz, O.; Colak, A.; Saglam, N.; Canakçi, S.; Beldüz, A.O. Determination and Characterization of Thermostable Esterolytic Activity from a Novel Thermophilic Bacterium Anoxybacillus gonensis A4. J. Biochem. Mol. Biol. 2007, 40, 588–594. [Google Scholar] [CrossRef]
- Dilokpimol, A.; Mäkelä, M.R.; Varriale, S.; Zhou, M.; Cerullo, G.; Gidijala, L.; Hinkka, H.; Brás, J.L.A.; Jütten, P.; Piechot, A.; et al. Fungal Feruloyl Esterases: Functional Validation of Genome Mining Based Enzyme Discovery Including Uncharacterized Subfamilies. New Biotechnol. 2018, 41, 9–14. [Google Scholar] [CrossRef]
- Lin, M.I.; Hiyama, A.; Kondo, K.; Nagata, T.; Katahira, M. Classification of Fungal Glucuronoyl Esterases (FGEs) and Characterization of Two New FGEs from Ceriporiopsis subvermispora and Pleurotus eryngii. Appl. Microbiol. Biotechnol. 2018, 102, 9635–9645. [Google Scholar] [CrossRef]
- Yamamoto-Tamura, K.; Hiradate, S.; Watanabe, T.; Koitabashi, M.; Sameshima-Yamashita, Y.; Yarimizu, T.; Kitamoto, H. Contribution of Soil Esterase to Biodegradation of Aliphatic Polyester Agricultural Mulch Film in Cultivated Soils. AMB Expr. 2015, 5, 10. [Google Scholar] [CrossRef]
- Ruginescu, R.; Gomoiu, I.; Popescu, O.; Cojoc, R.; Neagu, S.; Lucaci, I.; Batrinescu-Moteau, C.; Enache, M. Bioprospecting for Novel Halophilic and Halotolerant Sources of Hydrolytic Enzymes in Brackish, Saline and Hypersaline Lakes of Romania. Microorganisms 2020, 8, 1903. [Google Scholar] [CrossRef]
- Trovão, J.; Portugal, A. Current Knowledge on the Fungal Degradation Abilities Profiled through Biodeteriorative Plate Essays. Appl. Sci. 2021, 11, 4196. [Google Scholar] [CrossRef]
- Minotti, D.; Vergari, L.; Proto, M.R.; Barbanti, L.; Garzoli, S.; Bugli, F.; Sanguinetti, M.; Sabatini, L.; Peduzzi, A.; Rosato, R.; et al. Il Silenzio: The First Renaissance Oil Painting on Canvas from the Uffizi Museum Restored with a Safe, Green Antimicrobial Emulsion Based on Citrus aurantium var. amara Hydrolate and Cinnamomum zeylanicum Essential Oil. JoF 2022, 8, 140. [Google Scholar] [CrossRef] [PubMed]
- Paolo Cremonesi L’ambiente Acquoso per Il Trattamento Di Opere Policrome. In Metodologie, Tecniche e Formazione Nel Mondo Del Restauro; Il Prato Publishing House: Villatora, Italy, 2012; ISBN 978-88-6336-156-8.
- Borgioli, L.; de Comelli, A.; Pressi, G. Indagini Microbiologiche per La Verifica Dell’efficacia Di Alcuni Biocidi Esenti Da Metalli Pesanti. Progett. Restauro 2006, 11, 24–29. [Google Scholar]
- Ortega-Morales, B.O.; Gaylarde, C.C. Bioconservation of Historic Stone Buildings—An Updated Review. Appl. Sci. 2021, 11, 5695. [Google Scholar] [CrossRef]
- da Silva, M.T.C. Caldeira Novel Biocides for Cultural Heritage; Universidade de Évora: Évora, Portugal, 2017. [Google Scholar]
- Masi, M.; Petraretti, M.; De Natale, A.; Pollio, A.; Evidente, A. Fungal Metabolites with Antagonistic Activity against Fungi of Lithic Substrata. Biomolecules 2021, 11, 295. [Google Scholar] [CrossRef] [PubMed]
- Gomoiu, I.; Radvan, R.; Ghervase, L.; Mohanu, I.; Enache, M.; Neagu, S.; Ruginescu, R.; Cojoc, R. Cleaning of Mural Paintings and Mortars: Review. Rev. Rom. Mater. 2020, 50, 485–492. [Google Scholar]
- Gomoiu, I.; Enache, M.; Neagu, S.; Ruginescu, R.; Dumbrăvician, M.; Radvan, R.; Ghervase, L.; Mohanu, I.; Cojoc, R. Green Biotechnologies Used in the Restoration of Mural Painting and Lithic Support: Review. Rev. Rom. Mater. 2021, 51, 495–504. [Google Scholar]
- Marvasi, M.; Mastromei, G.; Perito, B. Bacterial Calcium Carbonate Mineralization in Situ Strategies for Conservation of Stone Artworks: From Cell Components to Microbial Community. Front. Microbiol. 2020, 11, 1386. [Google Scholar] [CrossRef]
- Castro-Alonso, M.J.; Montañez-Hernandez, L.E.; Sanchez-Muñoz, M.A.; Macias Franco, M.R.; Narayanasamy, R.; Balagurusamy, N. Microbially Induced Calcium Carbonate Precipitation (MICP) and Its Potential in Bioconcrete: Microbiological and Molecular Concepts. Front. Mater. 2019, 6, 126. [Google Scholar] [CrossRef]
- Nazel, T. Bioconsolidation of Stone Monuments. An Overview. Restor. Build. Monum. 2016, 22, 37–45. [Google Scholar] [CrossRef]
- Balloi, A.; Palla, F. Biocleaning. In Biotechnology and Conservation of Cultural Heritage; Palla, F., Barresi, G., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 67–84. ISBN 978-3-319-46166-3. [Google Scholar]
- Bosch-Roig, P.; Ranalli, G. The Safety of Biocleaning Technologies for Cultural Heritage. Front. Microbiol. 2014, 5, 155. [Google Scholar] [CrossRef]
- Ruginescu, R.; Enache, M.; Popescu, O.; Gomoiu, I.; Cojoc, R.; Batrinescu-Moteau, C.; Maria, G.; Dumbravician, M.; Neagu, S. Characterization of Some Salt-Tolerant Bacterial Hydrolases with Potential Utility in Cultural Heritage Bio-Cleaning. Microorganisms 2022, 10, 644. [Google Scholar] [CrossRef] [PubMed]
- Shirai, K.; Jackson, R.L. Lipoprotein Lipase-Catalyzed Hydrolysis of p-Nitrophenyl Butyrate. Interfacial Activation by Phospholipid Vesicles. J. Biol. Chem. 1982, 257, 1253–1258. [Google Scholar] [CrossRef]
- de Lourdes Moreno, M.; García, M.T.; Ventosa, A.; Mellado, E. Characterization of Salicola Sp. IC10, a Lipase- and Protease-Producing Extreme Halophile. FEMS Microbiol. Ecol. 2009, 68, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Bevilacqua, N. CTS Pure Artis’s Pigments. 2010. Available online: file:///D:/Downloads/CTS%20PURE%20ARTISTS%E2%80%99%20PIGMENTS.pdf (accessed on 31 July 2022).
- Cojoc, L.R.; Enache, M.I.; Neagu, S.E.; Lungulescu, M.; Setnescu, R.; Ruginescu, R.; Gomoiu, I. Carotenoids Produced by Halophilic Bacterial Strains on Mural Paintings and Laboratory Conditions. FEMS Microbiol. Lett. 2019, 366, fnz243. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, H.E.; Kolisis, F.N. An Investigation into the Removal of Starch Paste Adhesives from Historical Textiles by Using the Enzyme α-Amylase. J. Cult. Herit. 2011, 12, 169–179. [Google Scholar] [CrossRef]
- Gioventù, E.; Lorenzi, P. Bio-Removal of Black Crust from Marble Surface: Comparison with Traditional Methodologies and Application on a Sculpture from the Florence’s English Cemetery. Procedia Chem. 2013, 8, 123–129. [Google Scholar] [CrossRef]
- Bosch-Roig, P.; Pozo-Antonio, J.S.; Sanmartín, P. Identification of the Best-Performing Novel Microbial Strains from Naturally-Aged Graffiti for Biocleaning Research. Int. Biodeterior. Biodegrad. 2021, 159, 105206. [Google Scholar] [CrossRef]
- Derrick, M.R.; Stulik, D.; Landry, J.M. Infrared Spectroscopy in Conservation Science; Scientific Tools for Conservation; Getty Conservation Institute: Los Angeles, CA, USA, 1999; ISBN 978-0-89236-469-5. [Google Scholar]
- Minaei, F.; Ravandi, S.A.H.; Hejazi, S.M.; Alihosseini, F. The Fabrication and Characterization of Casein/PEO Nanofibrous Yarn via Electrospinning. e-Polymers 2019, 19, 154–167. [Google Scholar] [CrossRef]
- Cortea, I.M.; Ratoiu, L.; Ghervase, L.; Țentea, O.; Dinu, M. Investigation of Ancient Wall Painting Fragments Discovered in the Roman Baths from Alburnus maior by Complementary Non-Destructive Techniques. Appl. Sci. 2021, 11, 10049. [Google Scholar] [CrossRef]
- Genestar, C.; Pons, C. Earth Pigments in Painting: Characterisation and Differentiation by Means of FTIR Spectroscopy and SEM-EDS Microanalysis. Anal. Bioanal. Chem. 2005, 382, 269–274. [Google Scholar] [CrossRef]
- Naumann, A. Fourier Transform Infrared (FTIR) Microscopy and Imaging of Fungi. In Advanced Microscopy in Mycology; Dahms, T.E.S., Czymmek, K.J., Eds.; Fungal Biology; Springer International Publishing: Cham, Switzerland, 2015; pp. 61–88. ISBN 978-3-319-22436-7. [Google Scholar]
- Alves, P.D.D.; Siqueira, F.d.F.; Facchin, S.; Horta, C.C.R.; Victória, J.M.N.; Kalapothakis, E. Survey of Microbial Enzymes in Soil, Water, and Plant Microenvironments. TOMICROJ 2014, 8, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Souza, R.C.; Cantão, M.E.; Nogueira, M.A.; Vasconcelos, A.T.R.; Hungria, M. Outstanding Impact of Soil Tillage on the Abundance of Soil Hydrolases Revealed by a Metagenomic Approach. Braz. J. Microbiol. 2018, 49, 723–730. [Google Scholar] [CrossRef] [PubMed]
- El-Gendi, H.; Saleh, A.K.; Badierah, R.; Redwan, E.M.; El-Maradny, Y.A.; El-Fakharany, E.M. A Comprehensive Insight into Fungal Enzymes: Structure, Classification, and Their Role in Mankind’s Challenges. J. Fungi 2021, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Obahiagbon, K.O.; Owabor, C.N. Bio-Treatment of Crude Oil Polluted Water Using Mixed Microbial Populations of P. aureginosa, Penicillium notatum, E. coli and Aspergillus niger. AMR 2009, 62–64, 802–807. [Google Scholar] [CrossRef]
- Chandra, P.; Enespa; Singh, R.; Arora, P.K. Microbial Lipases and Their Industrial Applications: A Comprehensive Review. Microb. Cell Fact. 2020, 19, 169. [Google Scholar] [CrossRef]
- Lloyd, G.I.; Morris, E.O.; Smith, J.E. A Study of the Esterases and Their Function in Candida lipolytica, Aspergillus niger and a Yeast-like Fungus. J. Gen. Microbiol. 1970, 63, 141–150. [Google Scholar] [CrossRef]
- Abdel-Kareem, O. Microbiological Testing of Polymers and Resins Used in Conservation of Linen Textiles; Associazione Italiana Prove non Distruttive: Roma, Italy, 2000. [Google Scholar]
- Milanesi, C.; Baldi, F.; Borin, S.; Brusetti, L.; Ciampolini, F.; Iacopini, F.; Cresti, M. Deterioration of Medieval Painting in the Chapel of the Holy Nail, Siena (Italy) Partially Treated with Paraloid B72. Int. Biodeterior. Biodegrad. 2009, 63, 844–850. [Google Scholar] [CrossRef]
- Elhagrassy, A.F. Modification of EAPC-XYL by Pseudomonas Lipases Bacteria to Remove Acrylic from the Mural Oil Paintings. Shedet 2019, 6, 189–202. [Google Scholar] [CrossRef]
- Ranalli, G.; Zanardini, E. Biocleaning on Cultural Heritage: New Frontiers of Microbial Biotechnologies. J. Appl. Microbiol. 2021, 131, 583–603. [Google Scholar] [CrossRef]
- Bosch-Roig, P.; Regidor Ros, J.L.; Estellés, R.M. Biocleaning of Nitrate Alterations on Wall Paintings by Pseudomonas stutzeri. Int. Biodeterior. Biodegrad. 2013, 84, 266–274. [Google Scholar] [CrossRef]
- Ranalli, G.; Zanardini, E.; Rampazzi, L.; Corti, C.; Andreotti, A.; Colombini, M.P.; Bosch-Roig, P.; Lustrato, G.; Giantomassi, C.; Zari, D.; et al. Onsite Advanced Biocleaning System for Historical Wall Paintings Using New Agar-gauze Bacteria Gel. J. Appl. Microbiol. 2019, 126, 1785–1796. [Google Scholar] [CrossRef] [PubMed]
- Soffritti, I.; D’Accolti, M.; Lanzoni, L.; Volta, A.; Bisi, M.; Mazzacane, S.; Caselli, E. The Potential Use of Microorganisms as Restorative Agents: An Update. Sustainability 2019, 11, 3853. [Google Scholar] [CrossRef]
- Troiano, F.; Gulotta, D.; Balloi, A.; Polo, A.; Toniolo, L.; Lombardi, E.; Daffonchio, D.; Sorlini, C.; Cappitelli, F. Successful Combination of Chemical and Biological Treatments for the Cleaning of Stone Artworks. Int. Biodeterior. Biodegrad. 2013, 85, 294–304. [Google Scholar] [CrossRef]














Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomoiu, I.; Cojoc, R.; Ruginescu, R.; Neagu, S.; Enache, M.; Maria, G.; Dumbrăvician, M.; Olteanu, I.; Rădvan, R.; Ratoiu, L.-C.; et al. Brackish and Hypersaline Lakes as Potential Reservoir for Enzymes Involved in Decomposition of Organic Materials on Frescoes. Fermentation 2022, 8, 462. https://doi.org/10.3390/fermentation8090462
Gomoiu I, Cojoc R, Ruginescu R, Neagu S, Enache M, Maria G, Dumbrăvician M, Olteanu I, Rădvan R, Ratoiu L-C, et al. Brackish and Hypersaline Lakes as Potential Reservoir for Enzymes Involved in Decomposition of Organic Materials on Frescoes. Fermentation. 2022; 8(9):462. https://doi.org/10.3390/fermentation8090462
Chicago/Turabian StyleGomoiu, Ioana, Roxana Cojoc, Robert Ruginescu, Simona Neagu, Madalin Enache, Gabriel Maria, Maria Dumbrăvician, Ioana Olteanu, Roxana Rădvan, Lucian-Cristian Ratoiu, and et al. 2022. "Brackish and Hypersaline Lakes as Potential Reservoir for Enzymes Involved in Decomposition of Organic Materials on Frescoes" Fermentation 8, no. 9: 462. https://doi.org/10.3390/fermentation8090462