
Identification and Characterization of a Novel Cold-Adapted GH15 Family Trehalase from the Psychrotolerant Microbacterium phyllosphaerae LW106
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical Reagents and Strains
2.2. Bacteria Cultivation
2.3. Sequence, Phylogenetic, and Modeling Analysis
2.4. Cloning, Mutant Plasmid Construction, Expression, and Purification of MpTre15A
2.5. Trehalase Activity and Protein Concentration Determination
2.6. Substrate Specificity Assay
2.7. Optimum pH, Optimum Temperature, and Stability
2.8. Effects of Metal Ions and Other Chemicals
2.9. Kinetic Constants
2.10. Analysis of Trehalose Hydrolysis Products
3. Results
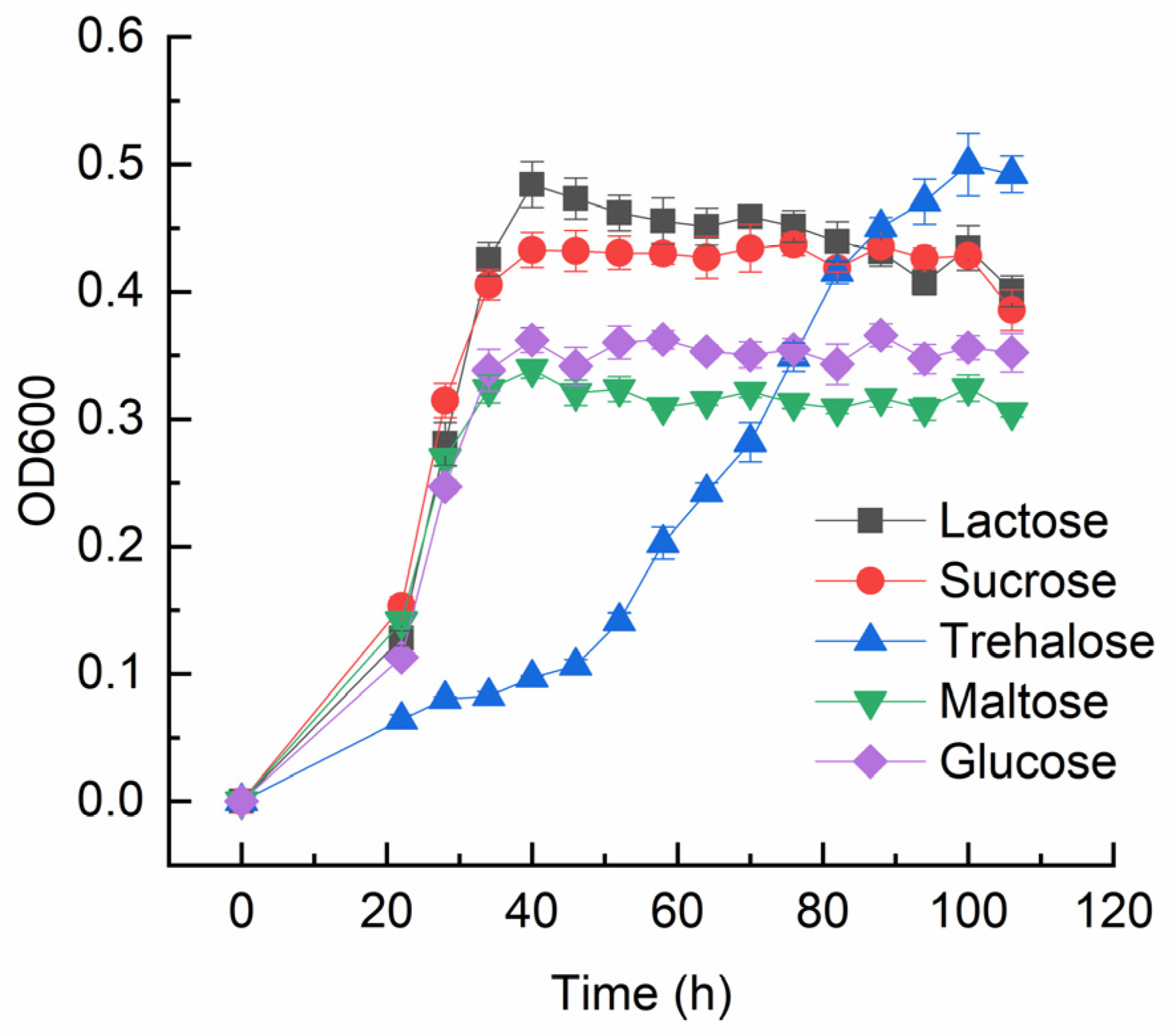
3.1. The Ability of M. phyllosphaerae LW106 to Utilize Trehalose as the Sole Source of Carbon
3.2. Bioinformatics Analysis of MpTre15A
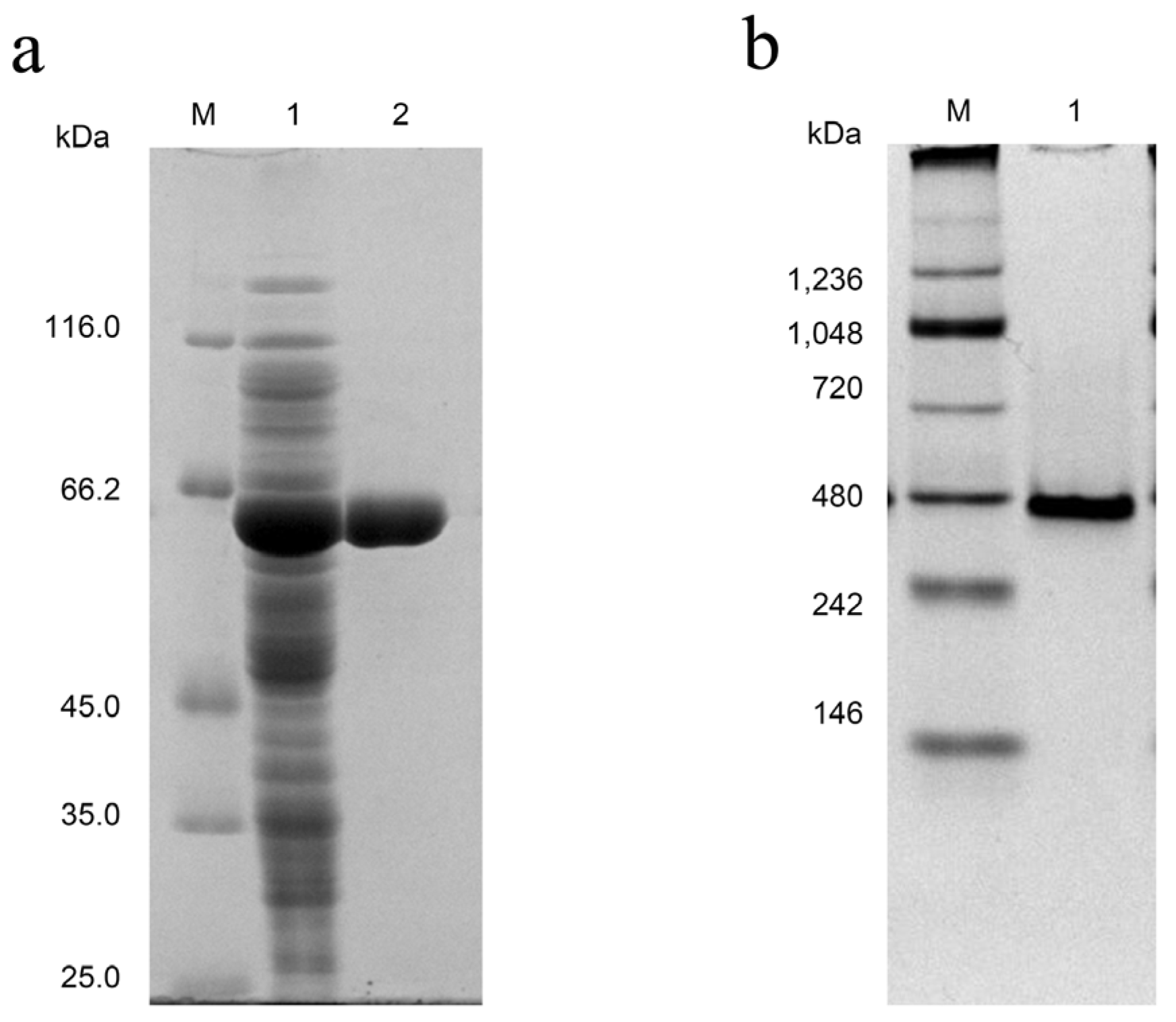
3.3. Expression, Purification, and Activity Assay of GA-Like Trehalase MpTre15A
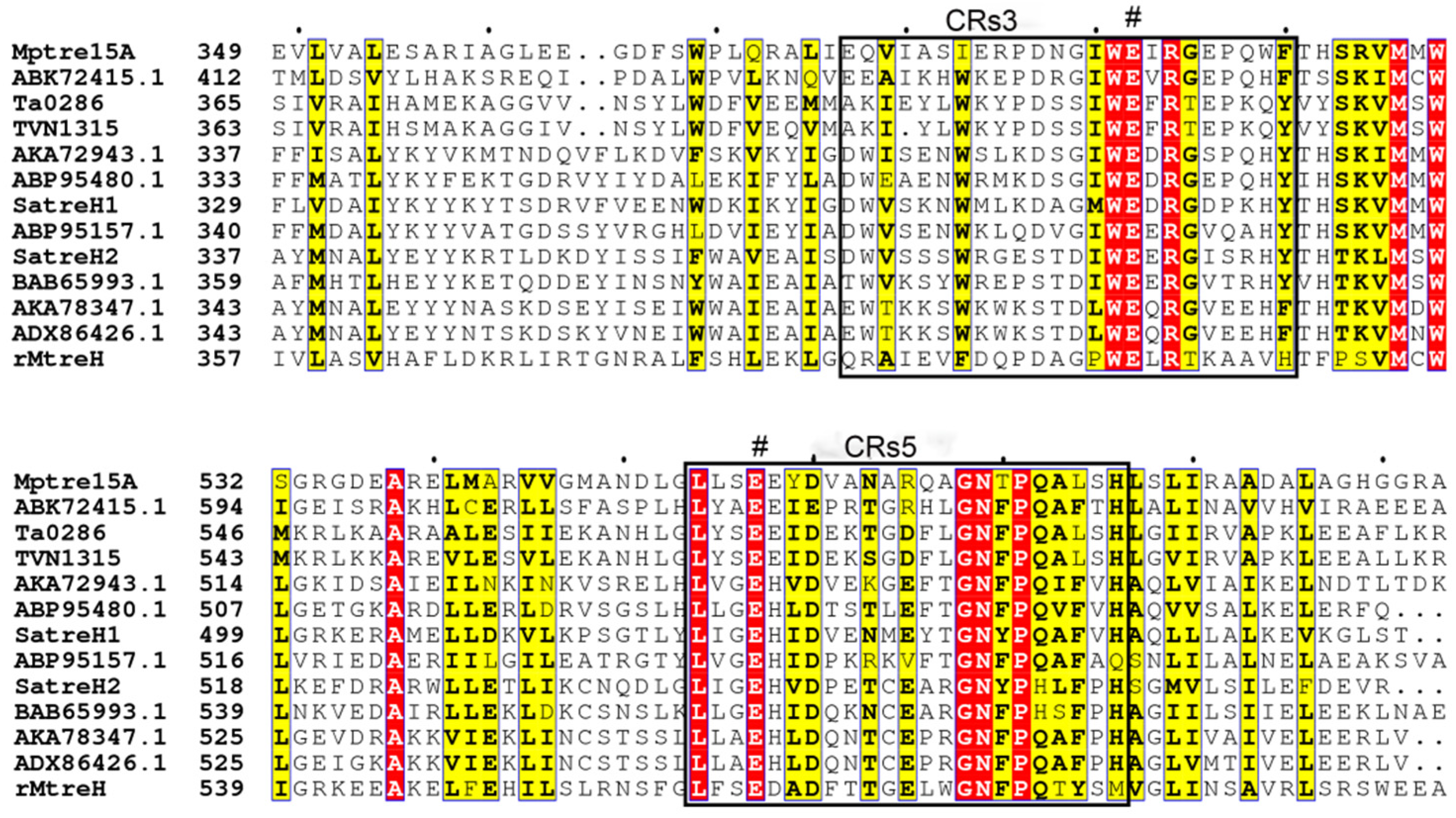
3.4. Characterization of the Catalytic Residues in MpTre15A
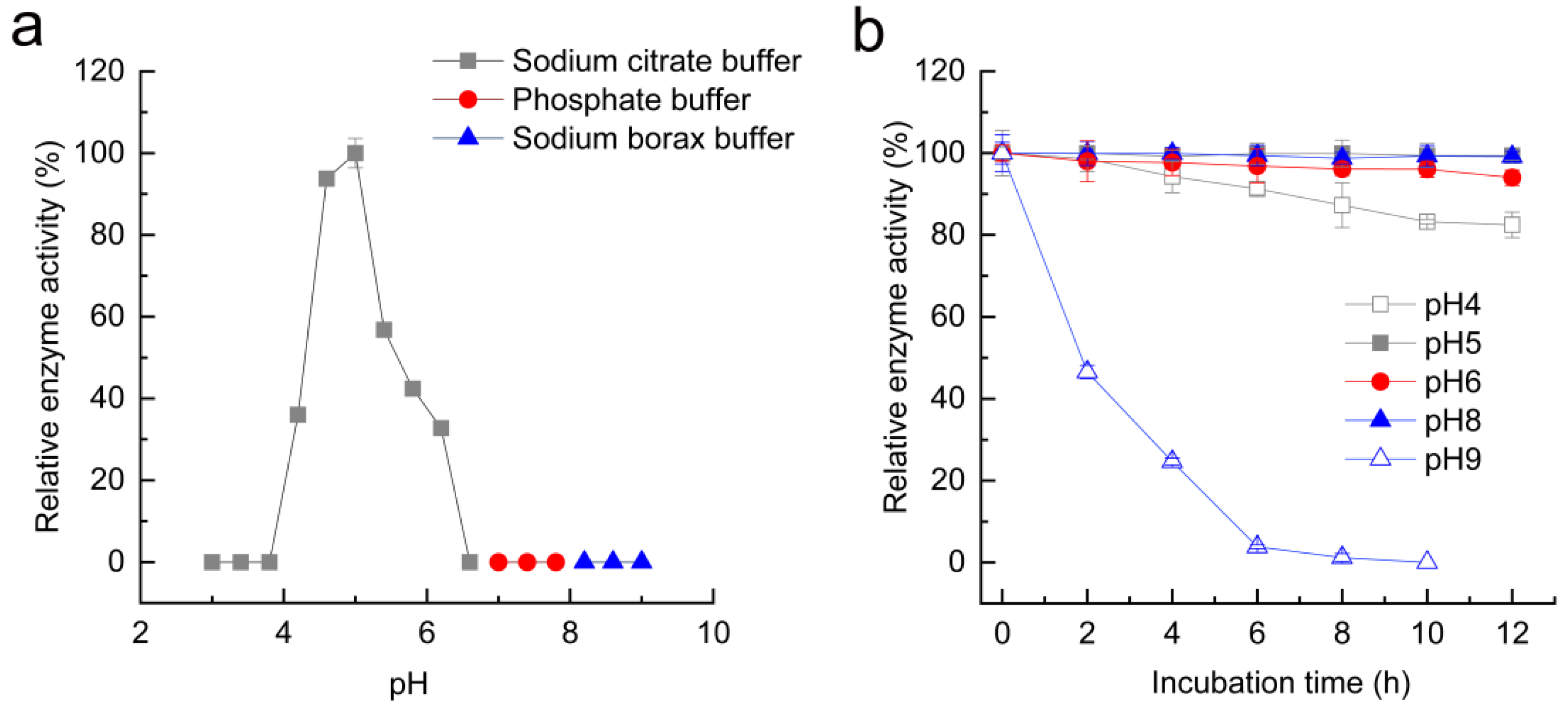
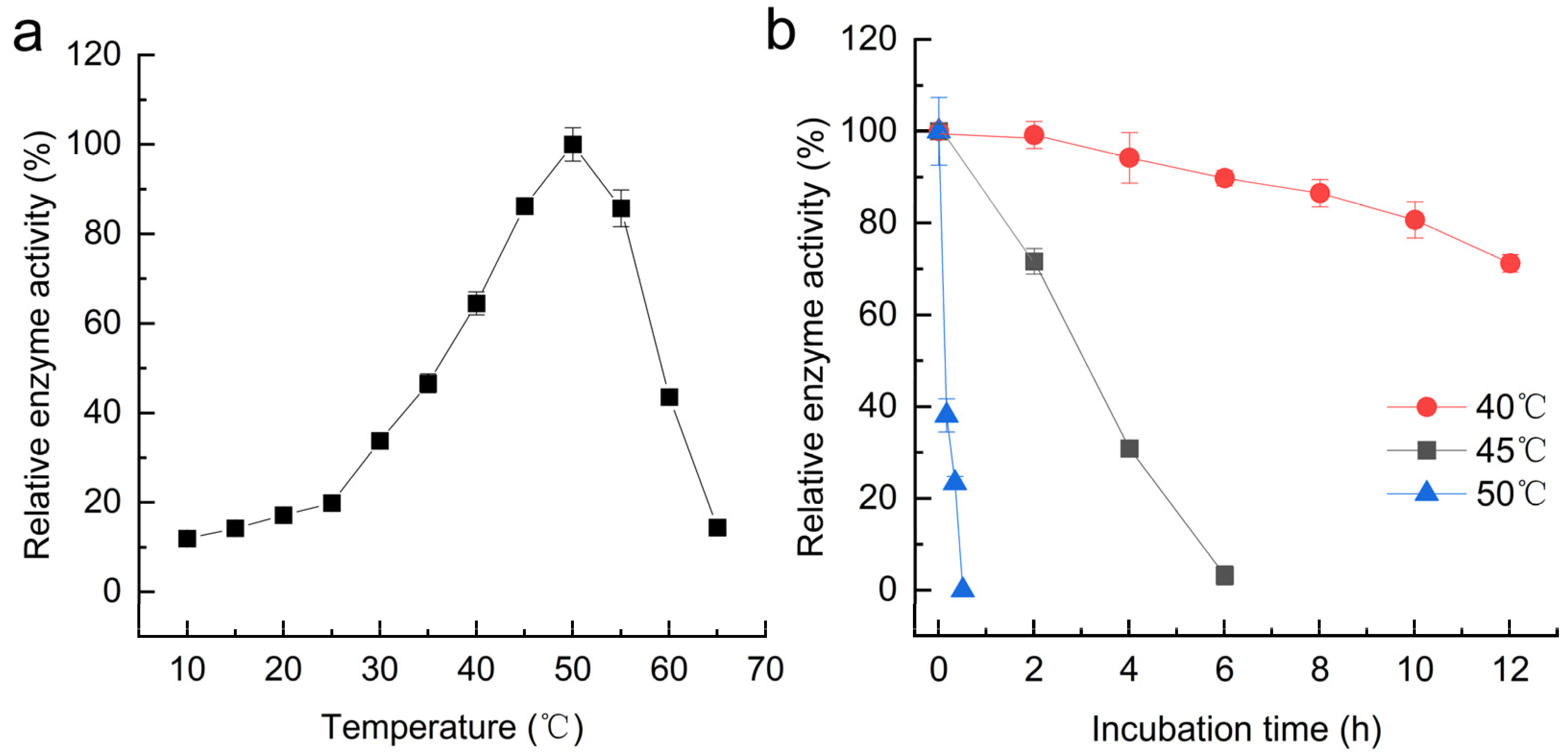
3.5. Effects of pH and Temperature on MpTre15A
3.6. Effects of Different Chemicals on the Activity of MpTre15A
3.7. Steady-State Kinetics Using Trehalose as the Substrate at Different Temperature
3.8. Time-Course Conversion of Trehalose into Glucose by MpTre15A
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elbein, A.D.; Pan, Y.T.; Pastuszak, I.; Carroll, D. New insights on trehalose: A multifunctional molecule. Glycobiology 2003, 13, 17R–27R. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.A.; Narrett, J.A.; Gardner, J.G. Trehalose degradation by Cellvibrio japonicus exhibits no functional redundancy and is solely dependent on the Tre37A enzyme. Appl. Environ. Microbiol. 2020, 86, 22. [Google Scholar] [CrossRef] [PubMed]
- Sussman, A.S.; Lingappa, B.T. Role of trehalose in ascospores of Neurospora tetrasperma. Science 1959, 130, 1343. [Google Scholar] [CrossRef]
- Argüelles, J.C. Physiological roles of trehalose in bacteria and yeasts: A comparative analysis. Arch. Microbiol. 2000, 174, 217–224. [Google Scholar] [PubMed]
- Thevelein, J.M.; Hohmann, S. Trehalose synthase: Guard to the gate of glycolysis in yeast? Trends Biochem. Sci. 1995, 20, 3–10. [Google Scholar] [CrossRef]
- Carroll, J.D.; Pastuszak, I.; Edavana, V.K.; Pan, Y.T.; Elbein, A.D. A novel trehalase from Mycobacterium smegmatis—Purification, properties, requirements. FEBS J. 2007, 274, 1701–1714. [Google Scholar] [CrossRef] [PubMed]
- Kandror, O.; DeLeon, A.; Goldberg, A.L. Trehalose synthesis is induced upon exposure of Escherichia coli to cold and is essential for viability at low temperatures. Proc. Natl. Acad. Sci. USA 2002, 99, 9727–9732. [Google Scholar] [CrossRef] [PubMed]
- Singer, M.A.; Lindquist, S. Multiple effects of trehalose on protein folding in vitro and in vivo. Mol. Cell 1998, 1, 639–648. [Google Scholar] [CrossRef]
- Higashiyama, T. Novel functions and applications of trehalose. Pure Appl. Chem. 2002, 74, 1263–1269. [Google Scholar] [CrossRef]
- Iturriaga, G.; Suárez, R.; Nova-Franco, B. Trehalose metabolism: From osmoprotection to signaling. Int. J. Mol. Sci. 2009, 10, 3793–3810. [Google Scholar] [CrossRef] [Green Version]
- Boos, W.; Ehmann, U.; Bremer, E.; Middendorf, A.; Postma, P. Trehalase of Escherichia coli. Mapping and cloning of its structural gene and identification of the enzyme as a periplasmic protein induced under high osmolarity growth conditions. J. Biol. Chem. 1987, 262, 13212–13218. [Google Scholar] [CrossRef]
- Horlacher, R.; Uhland, K.; Klein, W.; Ehrmann, M.; Boos, W. Characterization of a cytoplasmic trehalase of Escherichia coli. J. Bacteriol. 1996, 178, 6250–6257. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, M. Diverse and common features of trehalases and their contributions to microbial trehalose metabolism. Appl. Microbiol. Biotechnol. 2020, 104, 1837–1847. [Google Scholar] [CrossRef] [PubMed]
- Rimmele, M.; Boos, W. Trehalose-6-phosphate hydrolase of Escherichia coli. J. Bacteriol. 1994, 176, 5654–5664. [Google Scholar] [CrossRef]
- Moon, J.H.; Lee, W.; Park, J.; Choi, K.-H.; Cha, J. Characterization of a trehalose-degrading enzyme from the hyperthermophilic archaeon Sulfolobus acidocaldarius. J. Biosci. Bioeng. 2016, 122, 47–51. [Google Scholar] [CrossRef]
- Sakaguchi, M.; Shimodaira, S.; Ishida, S.; Amemiya, M.; Honda, S.; Sugahara, Y.; Oyama, F.; Kawakita, M. Identification of GH15 family thermophilic archaeal trehalases that function within a narrow acidic-pH range. Appl. Environ. Microbiol. 2015, 81, 4920–4931. [Google Scholar] [CrossRef]
- Yuasa, M.; Okamura, T.; Kimura, M.; Honda, S.; Shin, Y.; Kawakita, M.; Oyama, F.; Sakaguchi, M. Two trehalose-hydrolyzing enzymes from Crenarchaeon Sulfolobus acidocaldarius exhibit distinct activities and affinities toward trehalose. Appl. Microbiol. Biotechnol. 2018, 102, 4445–4455. [Google Scholar] [CrossRef]
- Dong, C.; Fan, Q.; Li, X.; Huang, Y.; Han, J.; Fang, X.; Huan, M.; Ye, X.; Li, Z.; Cui, Z. Expression and characterization of a novel trehalase from Microvirga sp. strain MC18. Protein Expr. Purif. 2021, 182, 105846. [Google Scholar] [CrossRef]
- Murray, I.A.; Coupland, K.; Smith, J.A.; Ansell, I.D.; Long, R.G. Intestinal trehalase activity in a UK population: Establishing a normal range and the effect of disease. Br. J. Nutr. 2000, 83, 241–245. [Google Scholar] [CrossRef]
- Kocot, B.; Vroemen, C.; Torres Pazmino, D.E.; Cannon, D.M., Jr.; Ward, D.E.; Foerster, H.; Shetty, J.K.; Clarkson, K.A.; Albers, K.; Van Brussel, M. Trehalase in fermentations. US Patent WO2015065978; filed 28 October 2014, and issued 7 May 2015,
- Feller, G.; Gerday, C. Psychrophilic enzymes: Hot topics in cold adaptation. Nat. Rev. Microbiol. 2003, 1, 200–208. [Google Scholar] [CrossRef]
- Mangiagalli, M.; Lotti, M. Cold-active β-galactosidases: Insight into cold adaptation mechanisms and biotechnological exploitation. Mar. Drugs. 2021, 19, 43. [Google Scholar] [CrossRef] [PubMed]
- Bruno, S.; Coppola, D.; di Prisco, G.; Giordano, D.; Verde, C. Enzymes from marine polar regions and their biotechnological applications. Mar. Drugs. 2019, 17, 544. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, L.; Li, M.; Gu, Y.; Guan, B.; Ni, Y. Isolation and identification of a Microbacterium sp. LW106 strain producing cold-active β-galactosidase, and study on its enzyme production conditions and enzymatic properties. Microbiol. China 2019, 46, 609–617. [Google Scholar]
- Wang, L.; Mou, Y.; Guan, B.; Hu, Y.; Zhang, Y.; Zeng, J.; Ni, Y. Genome sequence of the psychrophilic Cryobacterium sp. LW097 and characterization of its four novel cold-adapted β-galactosidases. Int. J. Biol. Macromol. 2020, 163, 2068–2083. [Google Scholar] [CrossRef] [PubMed]
- Gouet, P.; Robert, X. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar]
- Strom, A.R.; Kaasen, I. Trehalose metabolism in Escherichia coli: Stress protection and stress regulation of gene expression. Mol. Microbiol. 1993, 8, 205–210. [Google Scholar] [CrossRef]
- Morita, R.Y. Psychrophilic bacteria. Bacteriol. Rev. 1975, 39, 144–167. [Google Scholar] [CrossRef]
- Alley, R.B.; Cuffey, K.M.; Larson, G.J. How glaciers entrain and transport basal sediment: Physical constraints. Quat. Sci. Rev. 1997, 16, 1017–1038. [Google Scholar] [CrossRef]
- Lee, J.; Lee, A.; Moon, K.; Choi, K.-H.; Cha, J. Saci_1816: A Trehalase that Catalyzes Trehalose Degradation in the Thermoacidophilic Crenarchaeon Sulfolobus acidocaldarius. J. Mol. Biol. 2018, 28, 909–916. [Google Scholar] [CrossRef]
- Prasad, S.; Roy, I. Effect of disaccharides on the stabilization of bovine trypsin against detergent and autolysis. Biotechnol. Prog. 2009, 26, 627–635. [Google Scholar] [CrossRef]
- Rodriguez-Colinas, B.; Poveda, A.; Jimenez-Barbero, J.; Ballesteros, A.O.; Plou, F.J. Galacto-oligosaccharide synthesis from lactose solution or skim milk using the β-galactosidase from Bacillus circulans. J. Agric. Food Chem. 2012, 60, 6391–6398. [Google Scholar] [CrossRef] [PubMed]
- Cavicchioli, R.; Charlton, T.; Ertan, H.; Omar, S.M.; Siddiqui, K.S.; Williams, T.J. Biotechnological uses of enzymes from psychrophiles. Microb. Biotechnol. 2011, 4, 449–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Åqvist, J.; Isaksen, G.V.; Brandsdal, B.O. Computation of enzyme cold adaptation. Nat. Rev. Chem. 2017, 1, 0051. [Google Scholar] [CrossRef]
- Lonhienne, T.; Zoidakis, J.; Vorgias, C.E.; Feller, G.; Gerday, C.; Bouriotis, V. Modular structure, local flexibility and cold-activity of a novel chitobiase from a psychrophilic antarctic bacterium. J. Mol. Biol. 2001, 310, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Santiago, M.; Ramírez-Sarmiento, C.A.; Zamora, R.A.; Parra, L.P. Discovery, molecular mechanisms, and industrial applications of cold-active enzymes. Front. Microbiol. 2016, 7, 1408. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.C.P.; Terra, W.R.; Ferreira, C. The catalytic and other residues essential for the activity of the midgut trehalase from Spodoptera frugiperda. Insect Biochem. Mol. Biol. 2010, 40, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Q.; Gao, H.; Hu, N. A trehalase from Zunongwangia sp.: Characterization and improving catalytic efficiency by directed evolution. BMC Biotechnol. 2016, 16, 9. [Google Scholar] [CrossRef]
- Gibson, R.; Gloster, T.; Roberts, S.; Warren, R.; Storch de Gracia, I.; García, A.; Chiara, J.; Davies, G. Molecular basis for trehalase inhibition revealed by the structure of trehalase in complex with potent inhibitors. Angew. Chem. Int. Ed Engl. 2007, 46, 4115–4119. [Google Scholar] [CrossRef]
- Dong, L.; Yu, D.; Lin, X.; Wang, B.; Pan, L. Improving expression of thermostable trehalase from Myceliophthora sepedonium in Aspergillus niger mediated by the CRISPR/Cas9 tool and its purification, characterization. Protein Expr. Purif. 2020, 165, 105482. [Google Scholar] [CrossRef]
- Fang, X.; Mei, Z.; Chen, J.; Xiong, S.; Ning, J.; Jiang, C.; Gao, Y.; Yi, X.; Hui, G. A Trehalose quantitative sensor based on Ni foam material modified with graphene oxidated and non-linear analysis model. Food Anal. Methods 2021, 14, 1977–1985. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Relative Activity (%) |
|---|---|
| trehalose | 100 |
| maltose | 0 |
| cellobiose | 0 |
| sucrose | 0 |
| soluble starch | 0 |
| p-nitrophenyl-α-D-glucopyranoside | 0 |
| Enzyme | Relative Activity (%) |
|---|---|
| MpTre15A | 100 |
| E392Q | 0 |
| E557Q | 0 |
| Reagents | Relative Activity (%) | |
|---|---|---|
| 1 mM | 10 mM | |
| No addition a | 100.0 ± 1.2 | 100.0 ± 1.2 |
| NaCl | 100.4 ± 3.6 | 103.2 ± 4.9 |
| KCl | 100.5 ± 1.6 | 105.1 ± 2.4 |
| MgCl2 | 101.3 ± 2.6 | 126.4 ± 6.3 |
| MnCl2 | 114.8 ± 1.7 | 143.6 ± 6.4 |
| NiCl2 | 87.6 ± 4.1 | 57.2 ± 5.8 |
| CaCl2 | 84.0 ± 3.9 | 80.6 ± 4.3 |
| CoCl2 | 92.6 ± 6.7 | 90.6 ± 3.5 |
| FeCl2 | 93.2 ± 0.7 | 90.7 ± 3.8 |
| FeCl3 | 93.9 ± 5.8 | 60.1 ± 3.6 |
| AlCl3 | 96.0 ± 6.1 | 65.6 ± 1.2 |
| Cetyltrimethylammonium bromide (CTAB) | 77.3 ± 5.1 | 66.9 ± 5.5 |
| Ethylenediaminetetraacetic acid (EDTA) | 97.4 ± 4.6 | 90.1 ± 4.9 |
| Urea | 138.3 ± 1.8 | 156.8 ± 4.8 |
| Sodium dodecyl sulfate (SDS) | 0 | 0 |
| 1,4-Dithiothreitol (DTT) | 117.7 ± 5.4 | 265.3 ± 4.9 |
| β-Mercaptoethanol | 149.5 ± 4.9 | 251.6 ± 2.6 |
| Organism (Enzyme Name) | kcat (s−1) | Km (mM) | kcat/Km (s−1mM−1) | Temperature (°C) | pH | References |
|---|---|---|---|---|---|---|
| M. phyllosphaerae (Mptre15A) | 347.45 | 37.96 | 9.15 | 50 | 5.0 | This study |
| M. phyllosphaerae (Mptre15A) | 204.70 | 48.38 | 4.23 | 37 | 5.0 | This study |
| M. phyllosphaerae (Mptre15A) | 156.57 | 53.66 | 2.92 | 25 | 5.0 | This study |
| M. phyllosphaerae (Mptre15A) | 104.50 | 65.43 | 1.60 | 4 | 5.0 | This study |
| Mycolicibacterium smegmatis | ND a | 20.0 | ND a | 37 | 7.1 | Carroll et al. 2007 |
| Thermoplasma volcanium (TVN1315) | 63.0 | 48.7 | 1.29 | 50 | 3.7 | Sakaguchi et al. 2015 |
| T. acidophilum (Ta0286) | 66.7 | 40.2 | 1.66 | 50 | 3.7 | Sakaguchi et al. 2015 |
| Sulfolobus acidocaldarius (SaTreH1) | 77.0 | 41.8 | 1.84 | 50 | 4.0 | Yuasa et al. 2018 |
| S. acidocaldarius (SaTreH1) | 102.5 | 54.5 | 1.88 | 60 | 4.0 | Yuasa et al. 2018 |
| S. acidocaldarius (SaTreH2) | 1.60 | 2.39 | 0.67 | 50 | 3.7 | Yuasa et al. 2018 |
| S. acidocaldarius (SaTreH2) | 3.43 | 3.47 | 0.99 | 70 | 3.7 | Yuasa et al. 2018 |
| Microvirga sp. MC18 | ND a | 23.45 | ND a | 40 | 7.0 | Dong et al. 2021 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Yu, X.; Guan, B.; Hu, Y.; Li, X.; Zeng, J.; Ni, Y. Identification and Characterization of a Novel Cold-Adapted GH15 Family Trehalase from the Psychrotolerant Microbacterium phyllosphaerae LW106. Fermentation 2022, 8, 471. https://doi.org/10.3390/fermentation8100471
Zhang J, Yu X, Guan B, Hu Y, Li X, Zeng J, Ni Y. Identification and Characterization of a Novel Cold-Adapted GH15 Family Trehalase from the Psychrotolerant Microbacterium phyllosphaerae LW106. Fermentation. 2022; 8(10):471. https://doi.org/10.3390/fermentation8100471
Chicago/Turabian StyleZhang, Junhua, Xuehua Yu, Bo Guan, Youzhen Hu, Xu Li, Jun Zeng, and Yongqing Ni. 2022. "Identification and Characterization of a Novel Cold-Adapted GH15 Family Trehalase from the Psychrotolerant Microbacterium phyllosphaerae LW106" Fermentation 8, no. 10: 471. https://doi.org/10.3390/fermentation8100471