
In Vitro Fermentation and Degradation Characteristics of Rosemary Extract in Total Mixed Ration of Lactating Dairy Cows

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.1.1. Preparation of Rosemary Extract
2.1.2. In Vitro Process
2.2. Gas Production
2.3. In Vitro Degradability
2.4. Fermentation Parameters
2.5. Microbial Analysis
2.6. Statistical Analysis
3. Results
3.1. Gas Production
3.2. In Vitro Degradability
3.3. Fermentation Parameters
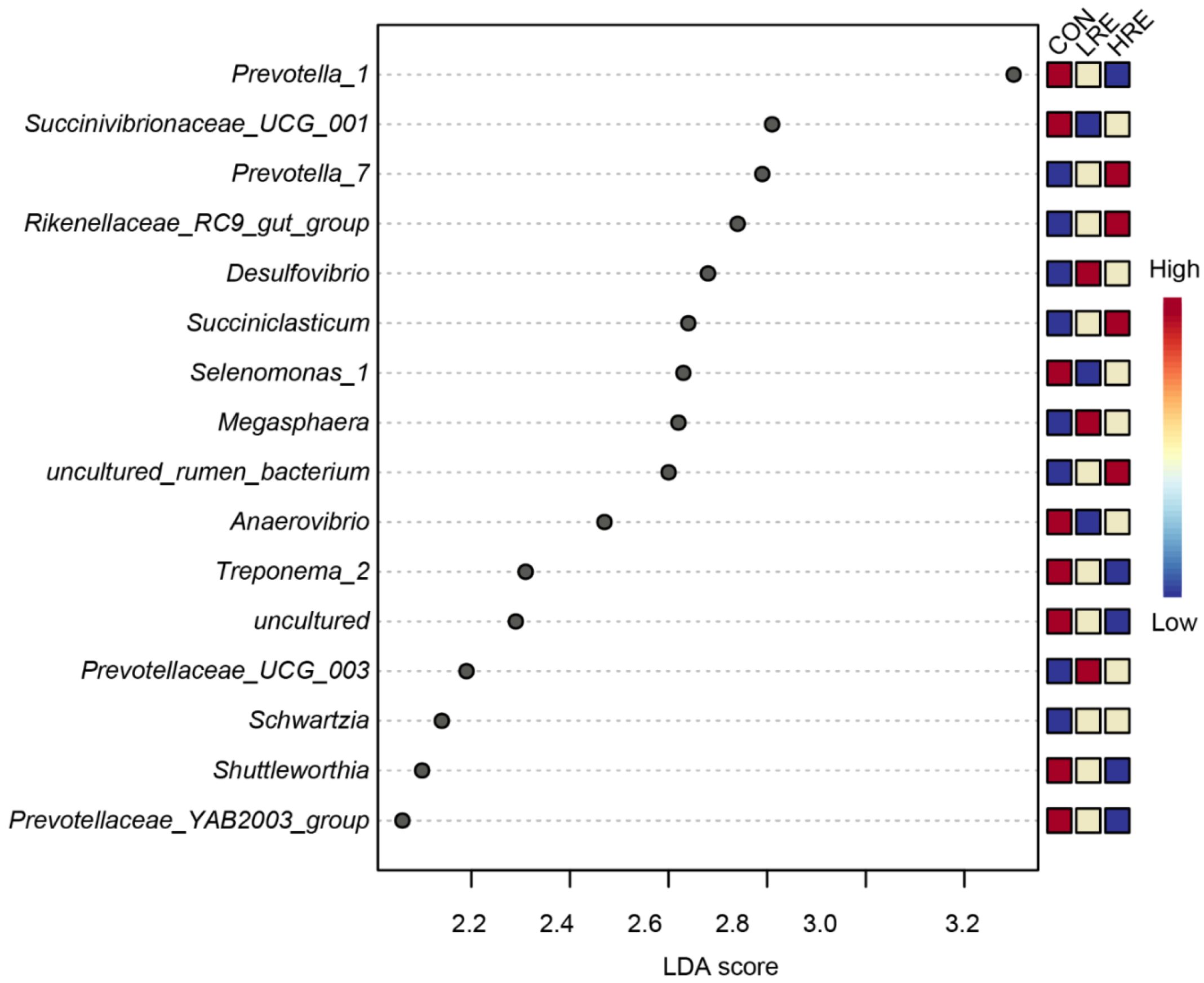
3.4. Microbial Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mizrahi, I.; Wallace, R.J.; Moraïs, S. The rumen microbiome: Balancing food security and environmental impacts. Nat. Rev. Microb. 2021, 19, 553–566. [Google Scholar] [CrossRef]
- Kim, E.T.; Kim, C.H.; Min, K.S.; Lee, S.S. Effects of Plant Extracts on Microbial Population, Methane Emission and Ruminal Fermentation Characteristics in In vitro. Asian-Australas. J. Anim. Sci. 2012, 25, 806–811. [Google Scholar] [CrossRef]
- Kolling, G.J.; Stivanin, S.C.B.; Gabbi, A.M.; Machado, F.S.; Ferreira, A.L.; Campos, M.M.; Tomich, T.R.; Cunha, C.S.; Dill, S.W.; Pereira, L.G.R.; et al. Performance and methane emissions in dairy cows fed oregano and green tea extracts as feed additives. J. Dairy Sci. 2018, 101, 4221–4234. [Google Scholar] [CrossRef] [PubMed]
- Okamura, N.; Fujimoto, Y.; Kuwabara, S.; Yagi, A. High-performance liquid chromatographic determination of carnosic acid and carnosol in Rosmarinus officinalis and Salvia officinalis. J. Chromatogr. A 1994, 679, 381–386. [Google Scholar] [CrossRef]
- Cuvelier, M.E.; Berset, C.; Richard, H. Antioxidant constituents in sage (Salvia officinalis). J. Agr. Food Chem. 1994, 42, 665–669. [Google Scholar] [CrossRef]
- de Macedo, L.M.; Santos, É.M.d.; Militão, L.; Tundisi, L.L.; Ataide, J.A.; Souto, E.B.; Mazzola, P.G. Rosemary (Rosmarinus officinalis L., syn Salvia rosmarinus Spenn.) and Its Topical Applications: A Review. Plants 2020, 9, 651. [Google Scholar] [CrossRef]
- Nieto, G.; Ros, G.; Castillo, J. Antioxidant and Antimicrobial Properties of Rosemary (Rosmarinus officinalis, L.): A Review. Medicines 2018, 5, 98. [Google Scholar] [CrossRef]
- Delgadillo-Ruiz, L.; Bañuelos-Valenzuela, R.; Gallegos-Flores, P.; Echavarría-Cháirez, F.; Meza-López, C.; Gaytán-Saldaña, N. Modification of ruminal fermentation in vitro for methane mitigation by adding essential oils from plants and terpenoid compounds. Abanico Vet. 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Güney, M.; Karaca, S.; Erdogan, S.; Kor, A.; Kale, C.; Onalan, S.; Demirel, M.; Bingol, N.T. Effects of dietary supplementation with rosemary oil on methanogenic bacteria density, blood and rumen parameters and meat quality of fattening lambs. Ital. J. Anim. Sci. 2021, 20, 794–805. [Google Scholar] [CrossRef]
- Molho-Ortiz, A.A.; Romero-Pérez, A.; Ramírez-Bribiesca, E.; Marquez-Mota, C.; Ramírez-Orejel, J.C.; Corona, L. PSXIV-13 Effect of phytochemicals on in vitro rumen fermentation and methane production. J. Anim. Sci. 2019, 97, 445–446. [Google Scholar] [CrossRef]
- Joch, M.; Cermak, L.; Hakl, J.; Hucko, B.; Duskova, D.; Marounek, M. In vitro screening of essential oil active compounds for manipulation of rumen fermentation and methane mitigation. Asian Australas. J. Anim. Sci. 2016, 29, 952. [Google Scholar] [CrossRef]
- Vegara, S.; Funes, L.; Martí, N.; Saura, D.; Micol, V.; Valero, M. Bactericidal activities against pathogenic bacteria by selected constituents of plant extracts in carrot broth. Food Chem. 2011, 128, 872–877. [Google Scholar] [CrossRef]
- Ivanovic, J.; Misic, D.; Zizovic, I.; Ristic, M. In vitro control of multiplication of some food-associated bacteria by thyme, rosemary and sage isolates. Food Control 2012, 25, 110–116. [Google Scholar] [CrossRef]
- Moreno, S.; Scheyer, T.; Romano, C.S.; Vojnov, A.A. Antioxidant and antimicrobial activities of rosemary extracts linked to their polyphenol composition. Free Radic. Res. 2006, 40, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Ruiz, D.R.; Bannink, A.; Dijkstra, J.; Kebreab, E.; Morgavi, D.P.; O’Kiely, P.; Reynolds, C.K.; Schwarm, A.; Shingfield, K.J.; Yu, Z.; et al. Design, implementation and interpretation of in vitro batch culture experiments to assess enteric methane mitigation in ruminants—A review. Anim. Feed Sci. Tech 2016, 216, 1–18. [Google Scholar] [CrossRef]
- Kong, F.; Lu, N.; Liu, Y.; Zhang, S.; Jiang, H.; Wang, H.; Wang, W.; Li, S. Aspergillus oryzae and Aspergillus niger Co-Cultivation Extract Affects In Vitro Degradation, Fermentation Characteristics, and Bacterial Composition in a Diet-Specific Manner. Animals 2021, 11, 1248. [Google Scholar] [CrossRef]
- Zhang, D.F.; Yang, H.J. In vitro ruminal methanogenesis of a hay-rich substrate in response to different combination supplements of nitrocompounds; pyromellitic diimide and 2-bromoethanesulphonate. Anim. Feed Sci. Tech. 2011, 163, 20–32. [Google Scholar] [CrossRef]
- Hao, Y.; Huang, S.; Liu, G.; Zhang, J.; Liu, G.; Cao, Z.; Wang, Y.; Wang, W.; Li, S. Effects of Different Parts on the Chemical Composition, Silage Fermentation Profile, In Vitro and In Situ Digestibility of Paper Mulberry. Animals 2021, 11, 413. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Liu, Y.; Wang, S.; Zhang, Y.; Wang, W.; Yang, H.; Lu, N.; Li, S. Nutrient Digestibility, Microbial Fermentation, and Response in Bacterial Composition to Methionine Dipeptide: An In Vitro Study. Biology 2022, 11, 93. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [PubMed]
- France, J.; Dijkstra, J.; Dhanoa, M.; Lopez, S.; Bannink, A. Estimating the extent of degradation of ruminant feeds from a description of their gas production profiles observed in vitro: Derivation of models and other mathematical considerations. Brit. J. Nutr. 2000, 83, 143–150. [Google Scholar] [CrossRef]
- Institute, S. Base SAS 9.4 Procedures Guide: Statistical Procedures; SAS Institute: Cary, NC, USA, 2017. [Google Scholar]
- Cobellis, G.; Petrozzi, A.; Forte, C.; Acuti, G.; Orrù, M.; Marcotullio, M.; Aquino, A.; Nicolini, A.; Mazza, V.; Trabalza-Marinucci, M. Evaluation of the Effects of Mitigation on Methane and Ammonia Production by Using Origanum vulgare L. and Rosmarinus officinalis L. Essential Oils on in Vitro Rumen Fermentation Systems. Sustainability 2015, 7, 12856–12869. [Google Scholar] [CrossRef]
- Tawfeeq, J.A.; Al-Omrani, H.A.; Shaker, R.M.; Hamza, Z.R.; Abbas, S.F.; Jabbar, R.H. Effect of Peppermint and Rosemary Extractions on Ruminant In-vitro Digestibility. Adv. Anim. Vet. Sci. 2019, 7, 910–913. [Google Scholar] [CrossRef]
- Johnson, K.A.; Johnson, D.E. Methane emissions from cattle. J. Anim. Sci. 1995, 73, 2483–2492. [Google Scholar] [CrossRef] [PubMed]
- Leahy, S.C.; Janssen, P.H.; Attwood, G.T.; Mackie, R.I.; McAllister, T.A.; Kelly, W.J. Electron flow: Key to mitigating ruminant methanogenesis. Trends Microbiol. 2022, 30, 209–212. [Google Scholar] [CrossRef]
- Cobellis, G.; Yu, Z.; Forte, C.; Acuti, G.; Trabalza-Marinucci, M. Dietary supplementation of Rosmarinus officinalis L. leaves in sheep affects the abundance of rumen methanogens and other microbial populations. J. Anim Sci. Biotechnol. 2016, 7, 27. [Google Scholar] [CrossRef]
- Castillejos, L.; Calsamiglia, S.; Martín-Tereso, J.; Ter Wijlen, H. In vitro evaluation of effects of ten essential oils at three doses on ruminal fermentation of high concentrate feedlot-type diets. Anim Feed Sci. Tech. 2008, 145, 259–270. [Google Scholar] [CrossRef]
- Sahraei, M.; Pirmohammadi, R.; Payvastegan, S. The effect of rosemary (Rosmarinus officinalis L.) essential oil on digestibility, ruminal fermentation and blood metabolites of Ghezel sheep fed barley-based diets. Span. J. Agric. Res. 2014, 12, 448–454. [Google Scholar] [CrossRef]
- Kholif, A.E.; Matloup, O.H.; Morsy, T.A.; Abdo, M.M.; Abu Elella, A.A.; Anele, U.Y.; Swanson, K.C. Rosemary and lemongrass herbs as phytogenic feed additives to improve efficient feed utilization, manipulate rumen fermentation and elevate milk production of Damascus goats. Livest Sci. 2017, 204, 39–46. [Google Scholar] [CrossRef]
- Kaneda, T. Iso- and anteiso-fatty acids in bacteria: Biosynthesis, function, and taxonomic significance. Microbiol. Rev. 1991, 55, 288–302. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, J.A.B.; Álvarez-Rivera, G.; Alves, R.C.; Costa, A.S.G.; Machado, S.; Cifuentes, A.; Ibáñez, E.; Oliveira, M.B.P.P. Comprehensive Phenolic and Free Amino Acid Analysis of Rosemary Infusions: Influence on the Antioxidant Potential. Antioxidants 2021, 10, 500. [Google Scholar] [CrossRef]
- Torres, R.N.S.; Moura, D.C.; Ghedini, C.P.; Ezequiel, J.M.B.; Almeida, M.T.C. Meta-analysis of the effects of essential oils on ruminal fermentation and performance of sheep. Small Rumin. Res. 2020, 189, 106148. [Google Scholar] [CrossRef]
- Jordán, M.J.; Castillo, J.; Bañón, S.; Martínez-Conesa, C.; Sotomayor, J.A. Relevance of the carnosic acid/carnosol ratio for the level of rosemary diterpene transfer and for improving lamb meat antioxidant status. Food Chem. 2014, 151, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Krause, D.O.; Denman, S.E.; Mackie, R.I.; Morrison, M.; Rae, A.L.; Attwood, G.T.; McSweeney, C.S. Opportunities to improve fiber degradation in the rumen: Microbiology, ecology, and genomics. FEMS Microbiol. Rev. 2003, 27, 663–693. [Google Scholar] [CrossRef]
- Cox, M.S.; Deblois, C.L.; Suen, G. Assessing the Response of Ruminal Bacterial and Fungal Microbiota to Whole-Rumen Contents Exchange in Dairy Cows. Front. Microbiol. 2021, 12, 665776. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Caro, E.; Holman, D.B.; Gzyl, K.E.; Moate, P.J.; Chaves, A.V. Ozone Decreased Enteric Methane Production by 20% in an in vitro Rumen Fermentation System. Front. Microbiol. 2020, 11, 571537. [Google Scholar] [CrossRef]
- De Oliveira Santos, E.; Thompson, F. The Family Succinivibrionaceae; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Pitta, D.W.; Pinchak, W.E.; Dowd, S.E.; Osterstock, J.; Gontcharova, V.; Youn, E.; Dorton, K.; Yoon, I.; Min, B.R.; Fulford, J. Rumen bacterial diversity dynamics associated with changing from bermudagrass hay to grazed winter wheat diets. Microb. Ecol. 2010, 59, 511–522. [Google Scholar] [CrossRef]
- Van Gylswyk, N. Succiniclasticum ruminis gen. nov., sp. nov., a ruminal bacterium converting succinate to propionate as the sole energy-yielding mechanism. Int. J. Syst. Evol. Mic. 1995, 45, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Van Gylswyk, N.; Hippe, H.; Rainey, F. Schwartzia succinivorans gen. nov., sp. nov., another ruminal bacterium utilizing succinate as the sole energy source. Int. J. Syst. Evol. Mic. 1997, 47, 155–159. [Google Scholar] [CrossRef]
- Liu, K.; Xu, Q.; Wang, L.; Wang, J.; Guo, W.; Zhou, M. The impact of diet on the composition and relative abundance of rumen microbes in goat. Asian-Australas. J. Anim. Sci. 2017, 30, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Huws, S.A.; Kim, E.J.; Lee, M.R.; Scott, M.B.; Tweed, J.K.; Pinloche, E.; Wallace, R.J.; Scollan, N.D. As yet uncultured bacteria phylogenetically classified as Prevotella, Lachnospiraceae incertae sedis and unclassified Bacteroidales, Clostridiales and Ruminococcaceae may play a predominant role in ruminal biohydrogenation. Environ. Microbiol. 2011, 13, 1500–1512. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Pu, Y.-Y.; Xie, Q.; Wang, J.-K.; Liu, J.-X. Pectin induces an in vitro rumen microbial population shift attributed to the pectinolytic Treponema group. Curr. Microbiol. 2015, 70, 67–74. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Items | Groups | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|
| CON | LRE | HRE | G | L | Q | ||
| GP, mL/g | 140.99 | 138.15 | 145.82 | 2.854 | 0.12 | 0.28 | 0.18 |
| A, mL/g | 142.37 | 139.75 | 160.34 | 6.499 | 0.07 | 0.10 | 0.20 |
| C, h−1 | 0.04 | 0.04 | 0.06 | 0.017 | 0.46 | 0.52 | 0.60 |
| HT, h | 1.76 | 1.94 | 1.81 | 0.075 | 0.14 | 0.61 | 0.14 |
| AGPR, mL/h | 49.58 | 41.16 | 58.16 | 4.732 | 0.25 | 0.25 | 0.07 |
| Items | Groups | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|
| CON | LRE | HRE | G | L | Q | ||
| CO2, % | 77.40 b | 79.77 a | 80.29 a | 0.244 | <0.01 | <0.01 | 0.02 |
| CH4, % | 12.50 a | 10.08 b | 9.52 b | 0.279 | <0.01 | <0.01 | 0.03 |
| H2, % | 10.10 | 10.16 | 10.19 | 0.172 | 0.94 | 0.75 | 0.96 |
| CO2, mL/g | 109.11 | 110.21 | 117.09 | 2.288 | 0.11 | 0.04 | 0.34 |
| CH4, mL/g | 17.64 a | 13.92 b | 13.88 b | 0.623 | 0.01 | 0.01 | 0.05 |
| H2, mL/g | 14.23 | 14.03 | 14.85 | 0.195 | 0.06 | 0.07 | 0.08 |
| Items | Groups | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|
| CON | LRE | HRE | G | L | Q | ||
| DM, % | 52.97 | 53.03 | 53.45 | 2.723 | 0.91 | 0.90 | 0.96 |
| NDF, % | 45.33 | 44.47 | 43.45 | 1.762 | 0.51 | 0.50 | 0.99 |
| ADF, % | 41.30 | 38.56 | 41.14 | 2.292 | 0.22 | 0.59 | 0.24 |
| CP, % | 62.49 | 62.59 | 63.12 | 2.413 | 0.86 | 0.98 | 0.94 |
| Items | Groups | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|
| CON | LRE | HRE | G | L | Q | ||
| pH value | 6.21 | 6.19 | 6.18 | 0.021 | 0.86 | 0.52 | 0.83 |
| NH3-N, mg/dL | 16.81 | 16.53 | 17.79 | 0.423 | 0.11 | 0.12 | 0.15 |
| Concentration, mmol/L | |||||||
| TVFA | 89.98 | 81.16 | 81.35 | 3.332 | 0.17 | 0.08 | 0.28 |
| Acetate | 57.76 a | 48.80 b | 44.79 b | 2.476 | <0.01 | <0.01 | 0.42 |
| Propionate | 19.31 b | 20.60 b | 25.27 a | 0.921 | <0.01 | <0.01 | 0.15 |
| Butyrate | 9.31 a | 7.60 b | 7.16 b | 0.431 | <0.01 | <0.01 | 0.24 |
| Valerate | 1.51 | 1.52 | 1.57 | 0.032 | 0.24 | 0.83 | 0.09 |
| BVFA | 2.08 b | 2.59 a | 2.61 a | 0.109 | <0.01 | <0.01 | 0.05 |
| Molar proportion, % | |||||||
| Acetate | 64.14 a | 59.85 b | 54.99 c | 0.954 | <0.01 | <0.01 | 0.819 |
| Propionate | 21.53 c | 25.40 b | 31.13 a | 0.741 | <0.01 | <0.01 | 0.314 |
| Butyrate | 10.35 | 9.48 | 8.80 | 0.467 | 0.06 | 0.03 | 0.861 |
| Valerate | 1.69 b | 2.00 a | 1.87 ab | 0.085 | 0.04 | 0.14 | 0.05 |
| BVFA | 2.30 b | 3.28 a | 3.21 a | 0.137 | <0.01 | <0.01 | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, F.; Wang, S.; Cao, Z.; Wang, Y.; Li, S.; Wang, W. In Vitro Fermentation and Degradation Characteristics of Rosemary Extract in Total Mixed Ration of Lactating Dairy Cows. Fermentation 2022, 8, 461. https://doi.org/10.3390/fermentation8090461
Kong F, Wang S, Cao Z, Wang Y, Li S, Wang W. In Vitro Fermentation and Degradation Characteristics of Rosemary Extract in Total Mixed Ration of Lactating Dairy Cows. Fermentation. 2022; 8(9):461. https://doi.org/10.3390/fermentation8090461
Chicago/Turabian StyleKong, Fanlin, Shuo Wang, Zhijun Cao, Yajing Wang, Shengli Li, and Wei Wang. 2022. "In Vitro Fermentation and Degradation Characteristics of Rosemary Extract in Total Mixed Ration of Lactating Dairy Cows" Fermentation 8, no. 9: 461. https://doi.org/10.3390/fermentation8090461