
Mapping Archaeal Diversity in Soda Lakes by Coupling 16S rRNA PCR-DGGE Analysis with Remote Sensing and GIS Technology

 ,
,  , , , , ,
, , , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
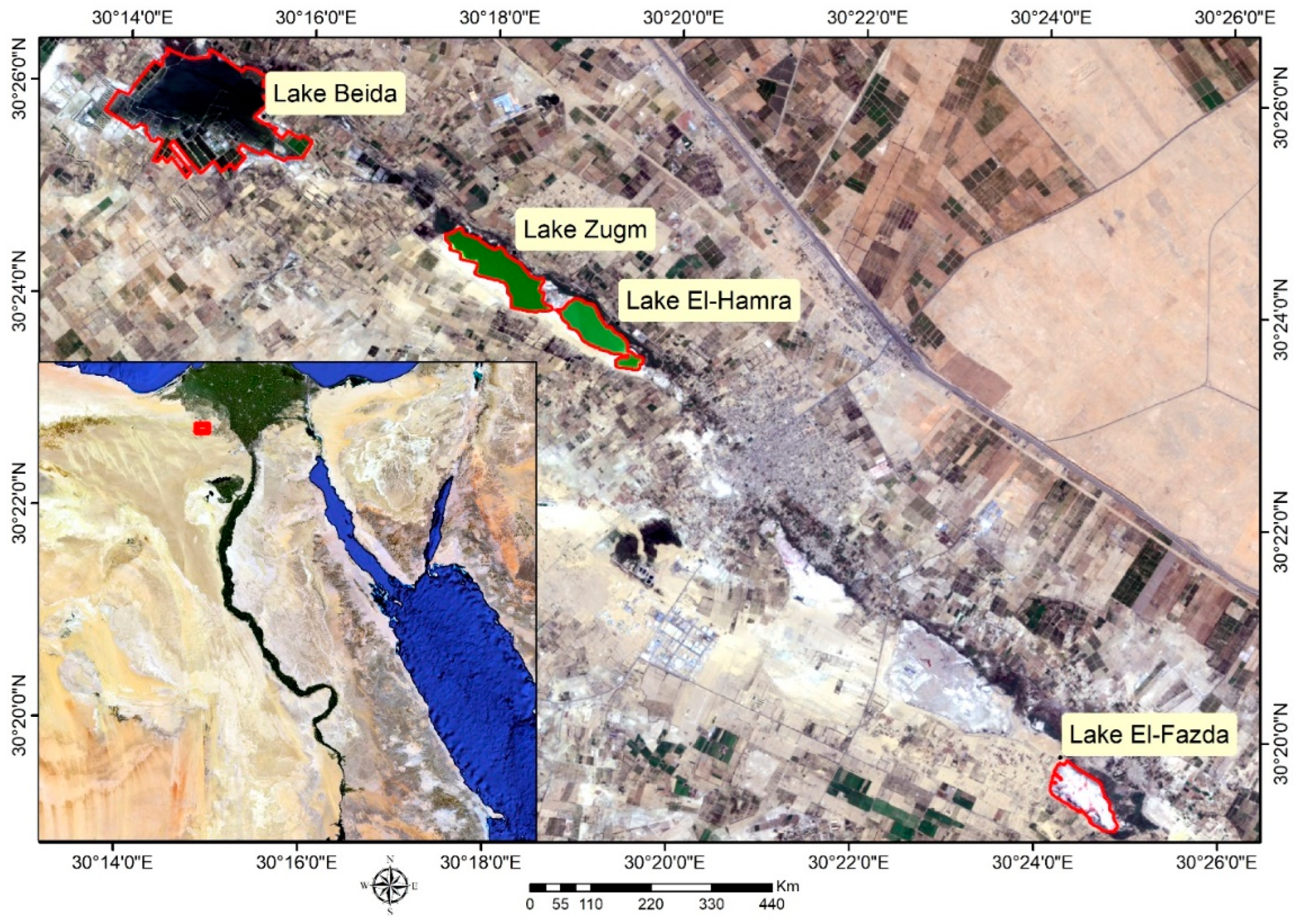
2.1. Site Description and Sample Collection
2.2. Molecular Assays
2.2.1. DNA Extraction
2.2.2. Quantitative PCR (qPCR)
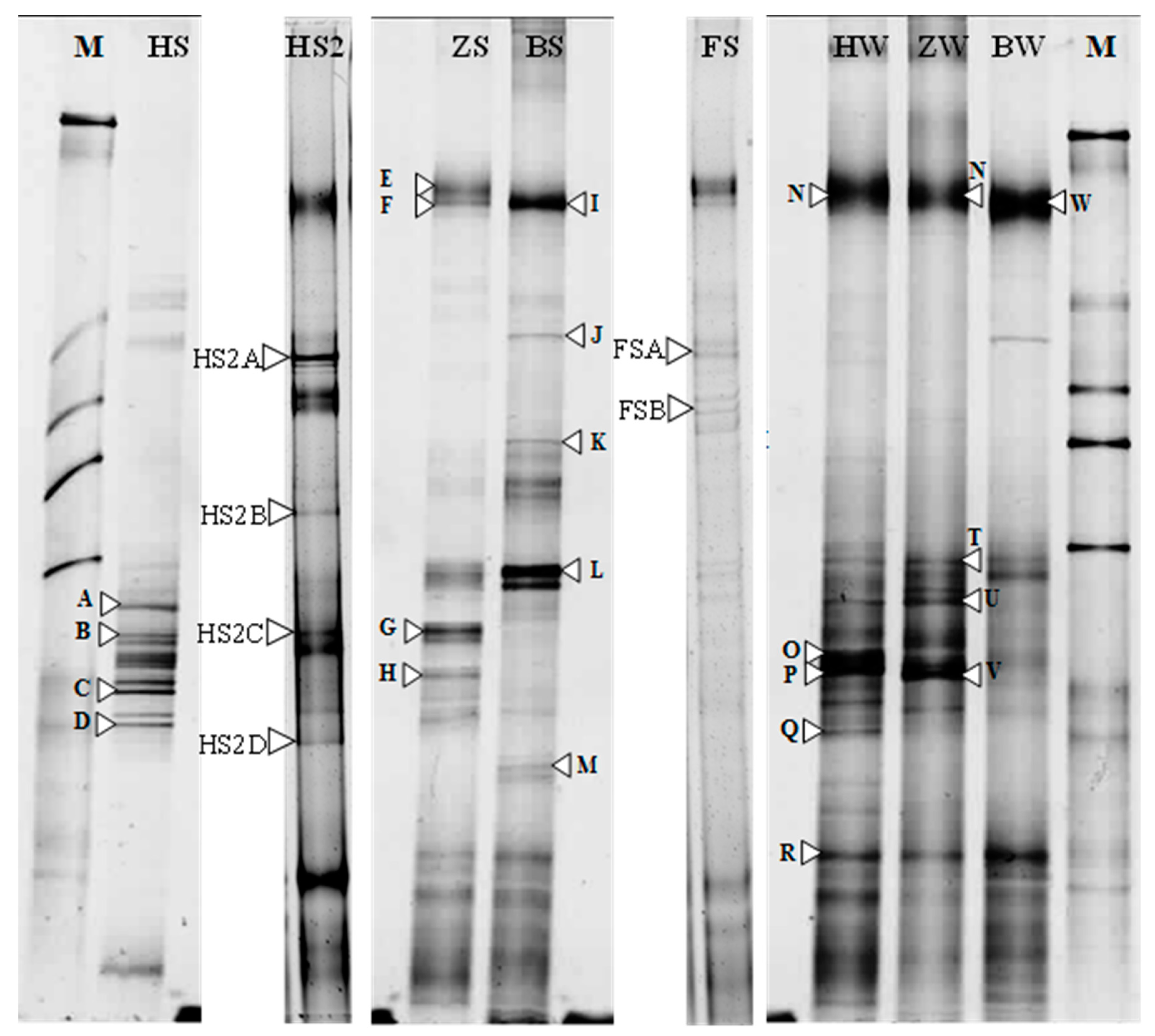
2.2.3. DGGE Analysis
2.3. Remote Sensing
2.3.1. Image Acquisition
2.3.2. Image Processing
2.4. Statistical Analysis
3. Results and Discussion
3.1. Physicochemical Analysis
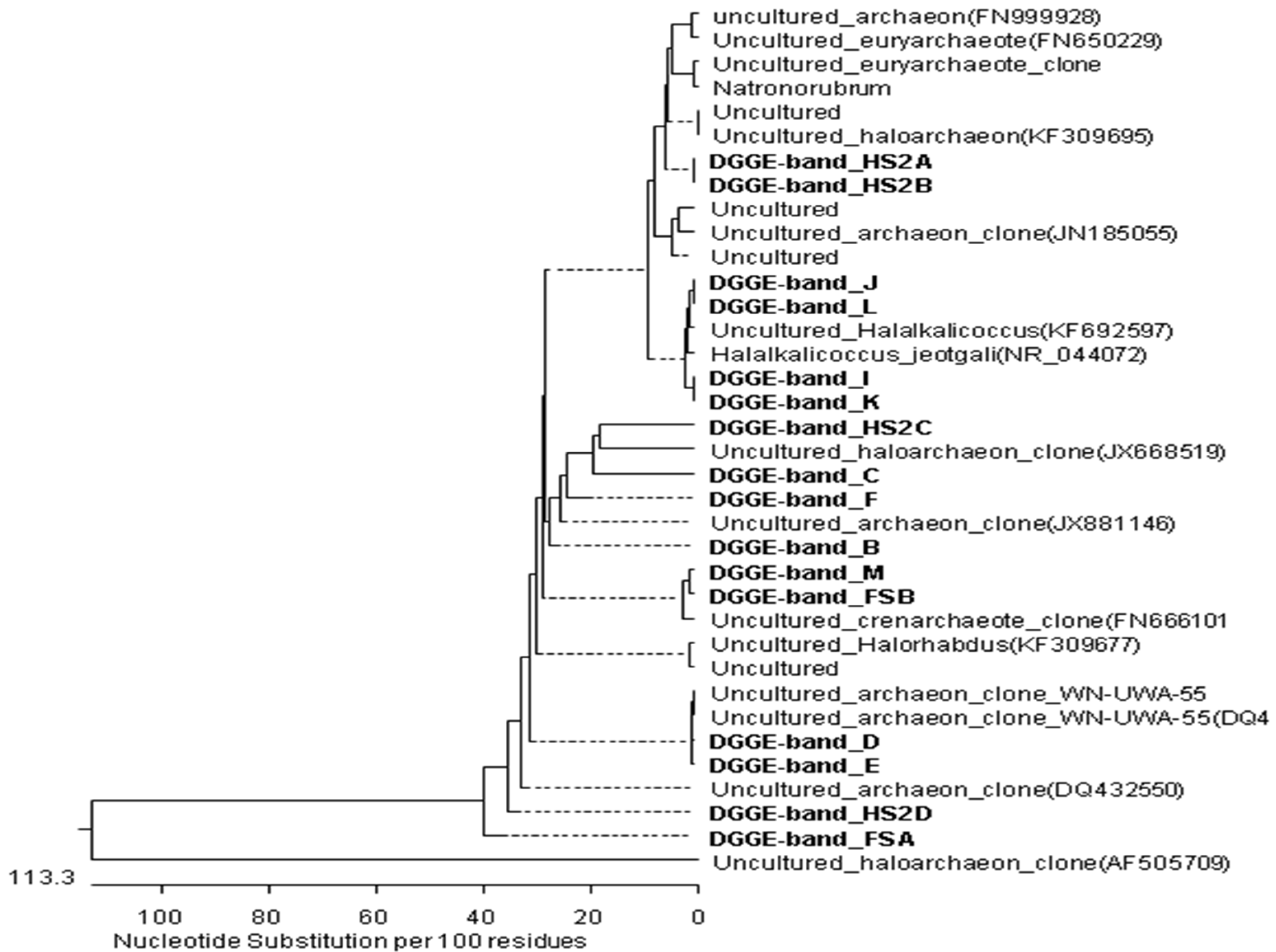
3.2. PCR-DGGE Analysis
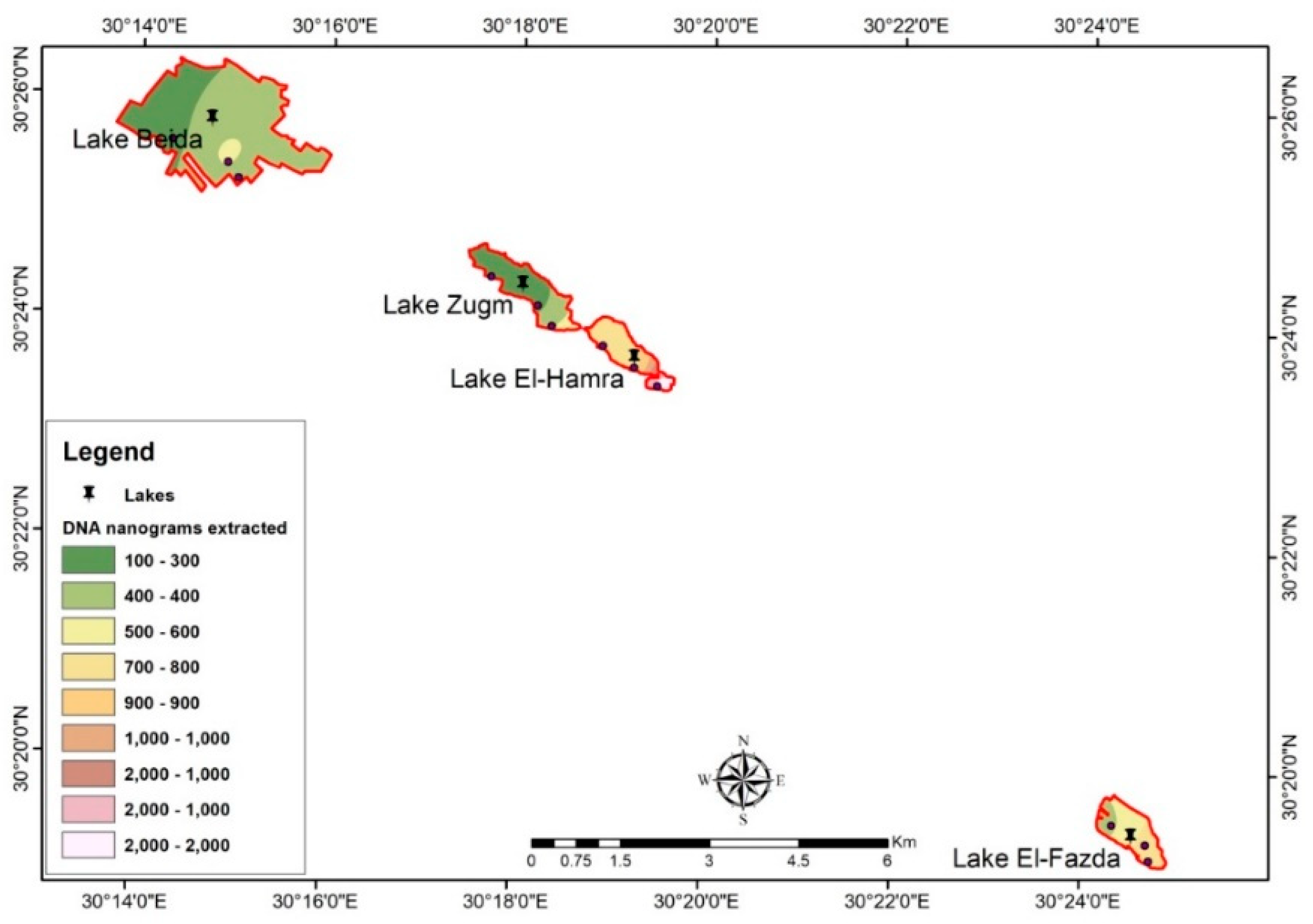
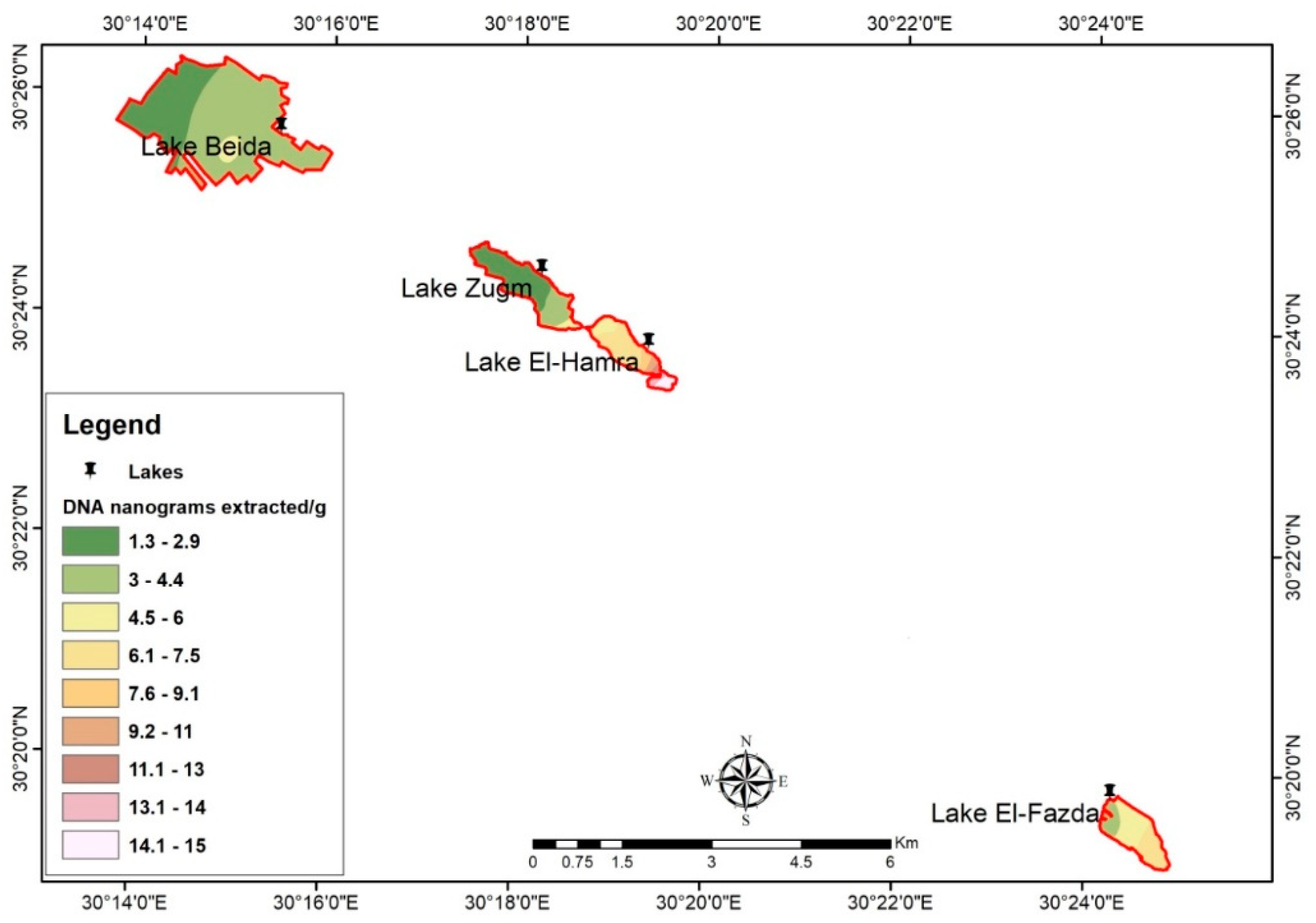
3.3. Spatial Distribution of Archaeal DNA by GIS and Remote Sensing
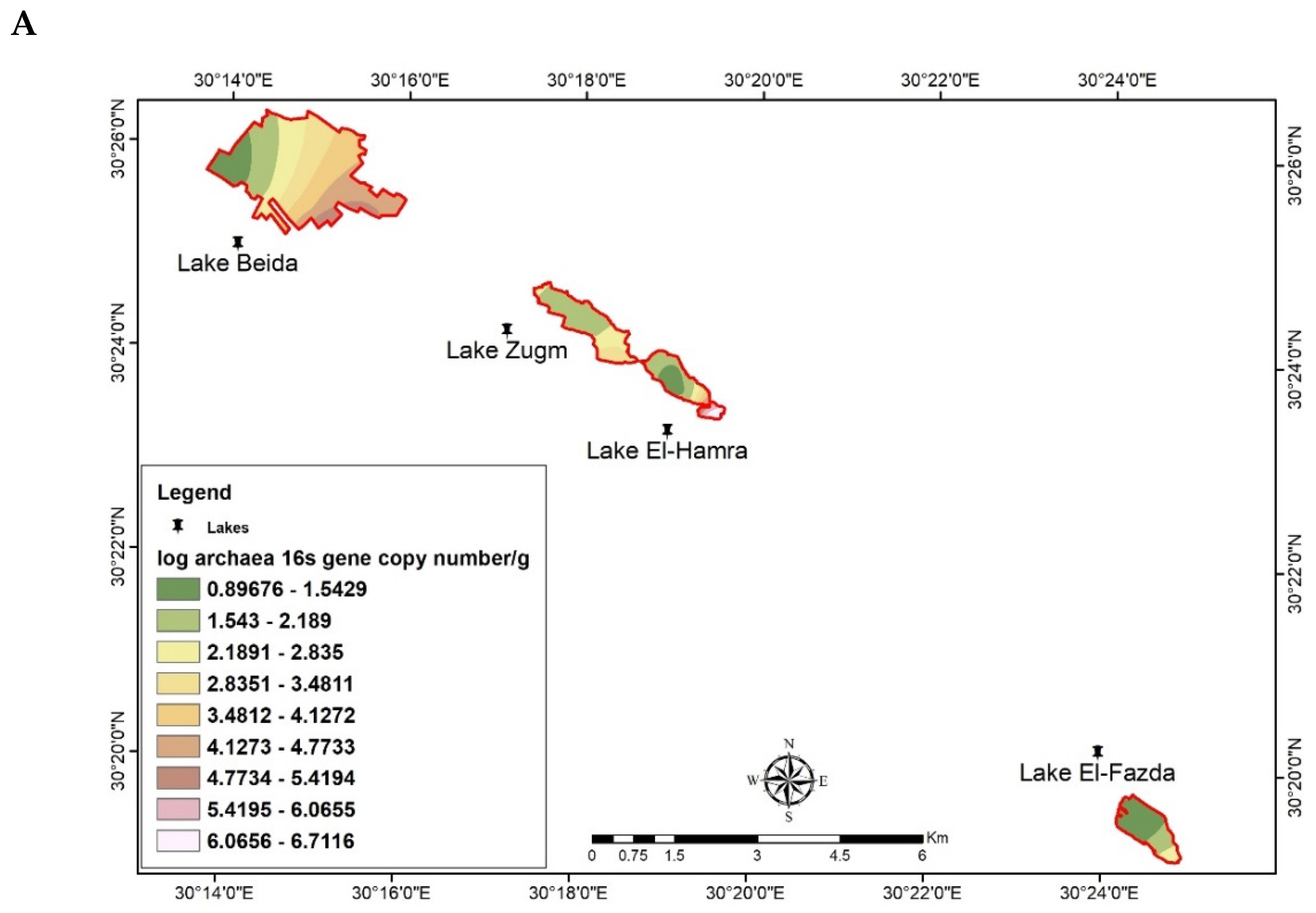
3.4. Spatial Distribution of Archaeal 16S rRNA Gene by GIS
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sayed, M.F.; Abdo, M.H. Assessment of environmental impact on, Wadi El-Natrun depression lakes water, Egypt. World J. Fish Mar. Sci. 2009, 1, 129–136. Available online: https://www.idosi.org/wjfms/wjfms1(2)09/9.pdf (accessed on 20 February 2022).
- Taher, A. Inland saline lakes of Wadi El Natrun depression, Egypt. Int. J. Salt Lake Res. 1999, 8, 149–169. [Google Scholar] [CrossRef]
- Boros, E.; Kolpakova, M. A review of the defining chemical properties of soda lakes and pans: An assessment on a large geographic scale of Eurasian inland saline surface waters. PLoS ONE 2018, 13, e0202205. [Google Scholar] [CrossRef] [PubMed]
- El-Malky, M.; Shalaby, A.; Khalifa, A.; Gabreel, M. Impact of agricultural expansion and urbanization on groundwater quality using spatial analysis of GIS and remote sensing techniques at Wadi El-Natrun area, Egypt. J. Environ. Sci. 2017, 40, 23–50. [Google Scholar]
- Masoud, A.; Atwia, A.M.G. Spatio-temporal characterization of the Pliocene aquifer conditions in Wadi El-Natrun area, Egypt, Environ. Earth Sci. 2011, 62, 1361–1374. [Google Scholar] [CrossRef]
- Sorokin, D.Y.; Kuenen, J.G. Alkaliphilic chemolithotrophs from sodas lakes. FEMS Microbiol. Ecol. 2005, 52, 287–295. [Google Scholar] [CrossRef]
- Foti, M.J.; Sorokin, D.Y.; Zacharova, E.E.; Pimenov, N.V.; Kuenen, J.G.; Muyzer, G. Bacterial diversity and activity along a salinity gradient in soda lakes of the Kulunda Steppe (Altai, Russia). Extremophiles 2008, 12, 133–145. [Google Scholar] [CrossRef]
- Sorokin, D.Y.; van Pelt, S.; Tourova, T.P.; Muyzer, G. Microbial isobutyronitrile utilization under haloalkaline conditions. Appl. Environ. Microbiol. 2007, 73, 5574–5579. [Google Scholar] [CrossRef]
- Rees, H.C.; Grant, W.D.; Jones, B.E.; Heaphy, S. Diversity of Kenyan soda lake alkaliphiles assessed by molecular methods. Extremophiles 2004, 8, 63–71. [Google Scholar] [CrossRef]
- Oremland, R.S.; Stolz, J.F.; Hollibaugh, J.T. The microbial arsenic cycle in Mono Lake, California. FEMS Microbial. Ecol. 2004, 48, 15–27. [Google Scholar] [CrossRef]
- Oremland, R.S.; Kulp, T.R.; Blum, J.S.; Hoeft, S.E.; Baesman, S.; Miller, L.G.; Stolz, J.F. A microbial arsenic cycle in a salt-saturated, Extreme environment. Science 2005, 308, 1305–1308. [Google Scholar] [CrossRef] [PubMed]
- Mesbah, N.M.; Abou-El-Ela, S.H.; Wiegel, J. Novel and unexpected prokaryotic diversity in water and sediments of the alkaline, hypersaline lakes of the Wadi An Natrun, Egypt. Microb. Ecol. 2007, 54, 598–617. [Google Scholar] [CrossRef] [PubMed]
- Enache, M.; Itoh, T.; Fukusguna, T.; Usami, R.; Dumitru, L.; Kamekura, M. Phylogenetic relationships within the family Halobacteriaceae inferred from rpoB’ gene and protein sequences. Int. J. Syst. Evol. Microbiol. 2007, 57, 2289–2295. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. Taxonomy of the family Halobacteriaceae: A paradigm for changing concepts in prokaryote systematics. Int. J. Syst. Evol. Microbiol. 2012, 62, 263–271. [Google Scholar] [CrossRef]
- Sorokin, D.Y.; Messina, E.; Smedile, F.; Roman, P.; Damste, J.S.S.; Ciordia, S.; Mena, M.C.; Ferrer, M.; Golyshin, P.N.; Kublanov, I.V.; et al. Discovery of anaerobic lithoheterotrophic haloarchaea, ubiquitous in hypersaline habitats. ISME J. 2017, 11, 1245–1260. [Google Scholar] [CrossRef]
- Minegishi, H.; Enomoto, S.; Echigo, A.; Shimane, Y.; Kondo, Y.; Inoma, A.; Kamekura, M.; Takai, K.; Itoh, T.; Ohkuma, M.; et al. Salinarchaeum chitinilyticum sp. nov., a chitin-degrading haloarchaeon isolated from commercial salt. Int. J. Syst. Evol. Microbiol. 2017, 67, 2274–2278. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, Y.; Lü, Z.Z.; Cui, H.L. Halomarina rubra sp. nov., isolated from a marine solar saltern. Arch. Microbiol. 2017, 199, 1431–1435. [Google Scholar] [CrossRef]
- Sorokin, D.Y.; Khijniak, T.V.; Elcheninov, A.G.; Toshchakov, S.V.; Kostrikina, N.A.; Bale, N.J.; Sinninghe Damsté, J.S.; Kublanov, I.V. Halococcoides cellulosivorans gen. nov., sp. nov., an extremely halophilic cellulose-utilizing haloarchaeon from hypersaline lakes. Int. J. Syst. Evol. Microbiol. 2019, 69, 1327–1335. [Google Scholar] [CrossRef]
- Youssef, N.H.; Savage-Ashlock, K.N.; McCully, A.L.; Luedtke, B.; Shaw, E.I.; Hoff, W.D.; Elshahed, M.S. Trehalose/2-sulfotrehalose biosynthesis and glycine-betaine uptake are widely spread mechanisms for osmoadaptation in the Halobacteriales. ISME J. 2014, 8, 636–649. [Google Scholar] [CrossRef]
- Grant, W.D.; Gemmell, R.T.; McGenity, T.J. Halobacteria: The evidence for longevity. Extremophiles 1998, 2, 279–287. [Google Scholar] [CrossRef]
- Oren, A. Life at high salt concentrations, intracellular KCl concentrations, and acidic proteomes. Front. Microbiol. 2013, 4, 315. [Google Scholar] [CrossRef] [PubMed]
- Oren, A.; Pri-El, N.; Shapiro, O.; Siboni, N. Buoyancy studies in natural communities of square gas-vacuolate archaea in saltern crystallizer ponds. Saline Syst. 2006, 2, 4. [Google Scholar] [CrossRef] [PubMed]
- Soppa, J. From genomes to function: Haloarchaea as model organisms. Microbiology 2006, 152, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Tsiamis, G.; Katsaveli, K.; Ntougias, S.; Kyrpides, N.; Andersen, G.; Piceno, Y.; Bourtzis, K. Prokaryotic community profiles at different operational stages of a Greek solar saltern. Res. Microbiol. 2008, 159, 609–627. [Google Scholar] [CrossRef]
- Kharroub, K.; Quesada, T.; Ferrer, R.; Fuentes, S.; Aguilera, M.; Boulahrouf, A.; Ramos-Cormenzana, A.; Monteoliva-Sánchez, M. Halorubrum ezzemoulense sp. nov., a halophilic archaeon isolated from Ezzemoul sabkha, Algeria. Int. J. Syst. Evol. Microbiol. 2006, 56, 1583–1588. [Google Scholar] [CrossRef] [PubMed]
- Luque, R.; González-Domenech, C.M.; Llamas, I.; Quesada, E.; Béjar, V. Diversity of culturable halophilic archaea isolated from Rambla Salada, Murcia (Spain). Extremophiles 2012, 16, 205–213. [Google Scholar] [CrossRef]
- Sorokin, D.Y.; Berben, T.; Melton, E.D.; Overmars, L.; Vavourakis, C.D.; Muyzer, G. Microbial diversity and biogeochemical cycling in soda lakes. Extremophiles 2014, 18, 791–809. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.J.; Allen, M.A.; DeMaere, M.Z.; Kyrpides, N.C.; Tringe, S.G.; Woyke, T.; Cavicchioli, R. Microbial ecology of an antarctic hypersaline lake: Genomic assessment of ecophysiology among dominant haloarchaea. ISME J. 2014, 8, 1645–1658. [Google Scholar] [CrossRef] [PubMed]
- Parte, A.C.; Sardà Carbasse, J.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of prokaryotic names with standing in nomenclature (LPSN) moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef] [PubMed]
- Ventosa, A.; Nieto, J.J. Biotechnological applications, and potentialities of halophilic microorganisms. World J. Microbiol. Biotechnol. 1995, 11, 85–94. [Google Scholar] [CrossRef]
- Schiraldi, C.; Mariateresa, G.; Mario, D. Perspectives on biotechnological applications of archaea. Archaea 2002, 1, 75–86. [Google Scholar] [CrossRef]
- Hamad, A.A.; Sharaf, M.; Hamza, M.A.; Selim, S.; Hetta, H.F.; El-Kazzaz, W. Investigation of the Bacterial Contamination and Antibiotic Susceptibility Profile of Bacteria Isolated from Bottled Drinking Water. Microbiol. Spectr. 2022, 10, e01516-21. [Google Scholar] [CrossRef] [PubMed]
- Najjari, A.; Elshahed, M.S.; Cherif, A.; Youssef, N.H. Patterns and determinants of halophilic archaea (Class Halobacteria) diversity in Tunisian endorheic salt lakes and sebkhet systems. Appl. Environ. Microbiol. 2015, 81, 4432–4441. [Google Scholar] [CrossRef] [PubMed]
- Baati, H.; Guermazi, S.; Amdouni, R.; Gharsallah, N.; Sghir, A.; Ammar, E. Prokaryotic diversity of a Tunisian multipond solar saltern. Extremophiles 2008, 12, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Youssef, N.H.; Ashlock-Savage, K.N.; Elshahed, M.S. Phylogenetic diversities and community structure of members of the extremely halophilic archaea (Order Halobacteriales) in multiple saline sediment habitats. Appl. Environ. Microbiol. 2012, 78, 1332–1344. [Google Scholar] [CrossRef] [PubMed]
- Henriet, O.; Fourmentin, J.; Delincé, B.; Mahillon, J. Exploring the diversity of extremely halophilic archaea in food-grade salts. Int. J. Food Microbiol. 2014, 191, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Durán-Viseras, A.; Andrei, A.-S.; Ghai, R.; Sánchez-Porro, C.; Ventosa, A. New halonotius species provide genomics-based insights into cobalamin synthesis in Haloarchaea. Front. Microbiol. 2019, 10, 1928. [Google Scholar] [CrossRef]
- Duarte, S.; Cássio, F.; Pascoal, C. Denaturing gradient gel electrophoresis (DGGE) in microbial ecology-insights from freshwaters. In Gel Electrophoresis-Principles and Basics; Magdeldin, S., Ed.; IntechOpen: Rijeka, Croatia, 2012; pp. 173–196. [Google Scholar] [CrossRef]
- Tapia, E.; Donoso-Bravo, A.; Cabrol, L.; Alves, M.M.; Pereira, A.; Rapaport, A.; Ruiz-Filippi, G. A methodology for a functional interpretation of DGGE with the help of mathematical modelling. Application in bio-hydrogen production. Water Sci. Technol. 2014, 69, 511–517. [Google Scholar] [CrossRef]
- Benítez-Cabello, A.; Bautista-Gallego, J.; Garrido-Fernández, A.; Rantsiou, K.; Cocolin, L.; Jiménez-Díaz, R.; Arroyo-López, F.N. RT-PCR–DGGE analysis to elucidate the dominant bacterial species of industrial Spanish-style green table olive fermentations. Front. Microbiol. 2016, 7, 1291. [Google Scholar] [CrossRef]
- Sharaf, M.; Arif, M.; Khan, S.; Abdalla, M.; Shabana, S.; Chi, Z.; Liu, C. Co-delivery of hesperidin and clarithromycin in a nanostructured lipid carrier for the eradication of Helicobacter pylori in vitro. Bioorganic Chem. 2021, 112, 104896. [Google Scholar] [CrossRef]
- Sörbo, B. Sulfate: Turbidometric and nephelometric methods. Methods Enzymol. 1987, 143, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Sharaf, M.; Sewid, A.H.; Hamouda, H.I.; Elharrif, M.G.; El-Demerdash, A.S.; Alharthi, A.; Hashim, N.; Hamad, A.A.; Selim, S.; Alkhalifah, D.H.M.; et al. Rhamnolipid-coated iron oxide nanoparticles as a novel multitarget candidate against major foodborne E. coli serotypes and methicillin-resistant S. aureus. Microbiol. Spectrum 2022, e00250-22. [Google Scholar] [CrossRef]
- Pérez, L.M.; Fittipaldi, M.; Adrados, B.; Morató, J.; Codony, F. Error estimation in environmental DNA targets quantification due to PCR efficiencies differences between real samples and standards. Folia Microbiol. 2013, 58, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Muyzer, G.; De, W.E.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microb. 1993, 59, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Lucena-Padrós, H.; Jiménez, E.; Maldonado-Barragán, A.; Rodríguez, J.M.; Ruiz-Barba, J.L. PCR-DGGE assessment of the bacterial diversity in Spanish-style green table-olive fermentations. Int. J. Food Microbiol. 2015, 205, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Webster, G.; Parkes, R.J.; Cragg, B.A.; Newberry, C.J.; Weightman, A.J.; Fry, J.C. Prokaryotic community composition and biogeochemical processes in deep subseafloor sediments from the Peru margin. FEMS Microbiol. Ecol. 2006, 58, 65–85. [Google Scholar] [CrossRef]
- O’Sullivan, L.A.; Webster, G.; Fry, J.C.; Parkes, R.J.; Weightman, A.J. Modified linker-PCR primers facilitate complete sequencing of DGGE DNA fragments. J. Microbiol. Methods 2008, 75, 579–581. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSIBLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematic; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Nicol, G.W.; Glover, L.A.; Prosser, J.I. Molecular analysis of methanogenic archaeal communities in managed and natural upland pasture soils. Global Change Biol. 2003, 9, 1451–1457. [Google Scholar] [CrossRef]
- Øvreås, L.; Forney, L.; Daae, F.L.; Torsvik, V. Distribution of bacterioplankton in meromictic lake Saelen-vannet, as determined by denaturing gradient gel electrophoresis of PCR-amplified gene fragments coding for 16S rRNA. Appl. Environ. Microbiol. 1997, 63, 3367–3373. [Google Scholar] [CrossRef] [PubMed]
- Sims, D.A.; Gamon, J.A. Estimation of vegetation water content and photosynthetic tissue area from spectral reflectance: A comparison of indices based on liquid water and chlorophyll absorption features. Remote Sens. Environ. 2003, 84, 526–537. [Google Scholar] [CrossRef]
- Salem, S.M.; El Gammal, E.-S.A. Salt minerals at Wadi El Natrun saline lakes, Egypt. New implications from remote sensing data. Eur. Chem. Bull. 2018, 7, 72–80. [Google Scholar] [CrossRef]
- DasSarma, S.; DasSarma, P. Halophiles and their enzymes: Negativity put to good use. Curr. Opin. Microbiol. 2015, 25, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Jabborova, D.; Annapurna, K.; Paul, S.; Kumar, S.; Saad, H.A.; Desouky, S.; Ibrahim, M.F.M.; Elkelish, A. Beneficial Features of Biochar and Arbuscular Mycorrhiza for Improving Spinach Plant Growth, Root Morphological Traits, Physiological Properties, and Soil Enzymatic Activities. JoF 2021, 7, 571. [Google Scholar] [CrossRef]
- Sharaf, M.; Hamouda, H.; Shabana, S.; Khan, S.; Arif, M.; Rozan, H.E.; Abdallae, M.; Chia, Z.; Liu, C. Design of lipid-based nanocarrier for drug delivery has a double therapy for six common pathogens eradication. Colloids Surfaces A: Physicochem. Eng. Aspects 2021, 625, 126662. [Google Scholar] [CrossRef]
- Grant, W.D. Alkaline environments. In Encyclopaedia of Microbiology, 1st ed.; Lederberg, J., Ed.; Academic Publishers: London, UK, 1992; pp. 73–80. [Google Scholar]
- Shannon, C.E.; Wiener, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949; p. 177. [Google Scholar]
- Margalef, R. Temporal succession and spatial heterogeneity in phytoplankton. In Perspectives in Marine Biology; Buzzati-Traverso, A.A., Ed.; University of California Press: Berkeley, CA, USA, 1958; pp. 323–347. [Google Scholar]
- Gugliandolo, C.; Lentini, V.; Maugeri, T. Distribution, and diversity of bacteria in a saline meromictic lake as determined by PCR-DGGE of 16S rRNA gene fragments. Curr. Microbiol. 2011, 62, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Makhdoumi-Kakhki, A.; Amoozegar, M.A.; Kazemi, B.; Pašić, L.; Ventosa, A. Prokaryotic diversity in Aran-Bidgol salt lake, the largest hypersaline playa in Iran. Microbes Environ. 2012, 27, 87–93. [Google Scholar] [CrossRef]
- Cytryn, E.; Minz, D.; Oremland, R.S.; Cohen, Y. Distribution and diversity of archaea corresponding to the limnological cycle of a hypersaline stratified lake (Solar Lake, Sinai, Egypt). Appl. Environ. Microbiol. 2000, 66, 3269–3276. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, K.B.; Canfield, D.E.; Teske, A.P.; Oren, A. Community composition of a hypersaline endoevaporitic microbial mat. Appl. Environ. Microbiol. 2005, 71, 7352–7365. [Google Scholar] [CrossRef] [PubMed]
- Ben Abdallah, M.; Karray, F.; Kallel, N.; Armougom, F.; Mhiri, N.; Quéméneur, M.; Cayol, J.-L.; Erauso, G.; Sayadi, S. Abundance and diversity of prokaryotes in ephemeral hypersaline lake Chott El Jerid using Illumina Miseq sequencing, DGGE and qPCR assays. Extremophiles 2018, 22, 811–823. [Google Scholar] [CrossRef]
- Najjari, A.; Stathopoulou, P.; Elmnasri, K.; Hasnaoui, F.; Zidi, I.; Sghaier, H.; Ouzari, H.I.; Cherif, A.; Tsiamis, G. Assessment of 16S rRNA gene-based phylogenetic diversity of archaeal communities in halite-crystal salts processed from natural Saharan saline systems of Southern Tunisia. Biology 2021, 10, 397. [Google Scholar] [CrossRef]
- Ochsenreiter, T.; Pfeifer, F.; Schleper, C. Diversity of Archaea in hypersaline environments characterized by molecular-phylogentic and cultivation studies. Extremophiles 2002, 6, 267–274. [Google Scholar] [CrossRef]
- Mohammed, A.B.; Mohamed, A.; El-Naggar, N.E.-A.; Mahrous, H.; Nasr, G.M.; Abdella, A.; Ahmed, R.H.; Irmak, S.; Elsayed, M.S.; Selim, S. Antioxidant and antibacterial activities of silver nanoparticles biosynthesized by Moringa Oleifera through response surface methodology. J. Nanomater. 2022, 2022, 9984308. [Google Scholar] [CrossRef]
- Wei, L.; Zhang, Y.; Huang, C.; Wang, Z.; Huang, Q.; Yin, F.; Guo, Y.; Cao, L. Inland lakes mapping for monitoring water quality using a detail/smoothing-balanced conditional random field based on Landsat-8/levels data. Sensors 2020, 20, 1345. [Google Scholar] [CrossRef] [PubMed]
- Hamed, A.F.; Salem, B.B.; El-Fatah, H.M.A. Floristic survey of blue-green algae/cyanobacteria in saline-alkaline lakes of Wadi El-Natrun (Egypt) by remote sensing application. Res. J. Appl. Sci. 2007, 3, 495–506. Available online: http://www.aensiweb.com/old/jasr/jasr/2007/495-506.pdf (accessed on 20 February 2022).
- Arif, M.; Sharaf, M.; Samreen; Dong, Q.; Wang, L.; Chi, Z.; Liu, C.-G. Bacteria-targeting chitosan/carbon dots nanocomposite with membrane disruptive properties improve eradication rate of Helicobacter pylori. J. Biomater. Sci. Polym. Ed. 2021, 32, 2423–2447. [Google Scholar] [CrossRef] [PubMed]
- Deole, R.; Challacombe, J.; Raiford, D.W.; Hoff, W.D. An extremely halophilic proteobacterium combines a highly acidic proteome with a low cytoplasmic potassium content. J. Biol. Chem. 2013, 288, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Selim, S.; Faried, O.A.; Almuhayawi, M.S.; Saleh, F.M.; Sharaf, M.; El Nahhas, N.; Warrad, M. Incidence of vancomycin-resistant Staphylococcus aureus strains among patients with urinary tract infections. Antibiotics 2022, 11, 408. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Pair | Sequence (5′–3′) a | Target of 16S rRNA Gene | Binding Position b | Reference |
|---|---|---|---|---|
| 1492R | GGTTACCTTGTTACGACT T | Archaea 16S rRNA | 1492–1510 | [50] |
| A2F | TTCCGGTTGATCCYGCCGGA | |||
| SAF-GC c,d | SA1f-CCTAYGGGGCGCAGCAGG | 341–358 | [51] | |
| SA2f-CCTACGGGGCGCAGAGGG | ||||
| PARCH519R | TTA CCG CGG CKG CTG | 519–533 | [52] | |
| M13R-GC | SAaf-CAGGAAACAGCTATGACG GGCGGGGCGGGGGCACGGGG GGCCTACGGGGCGCAGCAGG | [48] | ||
| SAbf-CAGGAAACAGCTATGACG GGGGCGGGGCGGGGGCACGG GGCCTATGGGGCGCAGCAGG | ||||
| M13F e | GTAAAACGACGGCCAG |
| Sample ID | pH | EC (dSm−1) | Na+ | K+ | Ca2+ | Mg2+ | CO32− | HCO3− | Cl− | SO42− |
|---|---|---|---|---|---|---|---|---|---|---|
| H | 9.03 ± 0.38 a | 151.3 ± 2.5 c | 24.24 ± 1.14 b | 0.110 ± 0.010 a | 0.10 ± 0.01 c | 0.48 ± 0.04 c | 10.80 ± 0.54 b | 7.75 ± 0.28 b | 26.18 ± 1.20 bc | 6.00 ± 0.24 b |
| Z | 8.89 ± 0.44 a | 188.3 ± 3.8 b | 31.42 ± 1.11 a | 0.057 ± 0.003 c | 0.23 ± 0.01 b | 0.72 ± 0.05 a | 21.30 ± 0.11 a | 13.95 ± 1.10 a | 28.95 ± 1.20 b | 5.48 ± 0.35 bc |
| B | 8.70 ± 0.31 a | 137.4 ± 2.7 d | 21.96 ± 1.20 b | 0.076 ± 0.005 b | 0.32 ± 0.01 a | 0.42 ± 0.03 c | 9.33 ± 0.46 c | 6.60 ± 0.24 bc | 24.65 ± 1.10 c | 5.04 ± 0.25 c |
| F * | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. |
| HS | 8.58 ± 0.28 a | 100.8 ± 2.1 f | 16.01 ± 1.10 c | 0.105 ± 0.010 a | 0.21 ± 0.03 b | 0.12 ± 0.01 d | 5.12 ± 0.25 d | 5.27 ± 0.16 d | 18.75 ± 0.90 d | 2.28 ± 0.11 e |
| BS | 8.64 ± 0.33 a | 141.5 ± 3.3 d | 23.17 ± 1.28 b | 0.066 ± 0.005 bc | 0.11 ± 0.01 c | 0.18 ± 0.01 d | 3.31 ± 0.16 e | 1.46 ± 0.27 e | 32.30 ± 1.30 a | 3.72 ± 0.18 d |
| ZS | 8.68 ± 0.43 a | 197.7 ± 4.9 a | 29.95 ± 1.40 a | 0.031 ± 0.002 d | 0.16 ± 0.01 b | 0.63 ± 0.03 b | 11.6 ± 0.58 b | 6.10 ± 0.31 cd | 26.60 ± 1.33 bc | 2.10 ± 0.11 e |
| FS | 8.56 ±0.41 a | 119.2 ± 2.9 e | 18.46 ± 1.21 c | 0.038 ± 0.002 d | 0.20 ± 0.02 b | 0.60 ± 0.03 b | 0.32 ± 0.15 f | 0.84 ± 0.05 e | 26.52 ± 1.21 bc | 8.12 ± 0.51 a |
| Data Type | Sediment Samples | Water Samples | ||||||
|---|---|---|---|---|---|---|---|---|
| El-Hamra | Zugm | Beida | El-Fazda | El-Hamra | Zugm | Beida | El-Fazda | |
| S | 17 | 16 | 20 | 12 | 18 | 16 | 14 | N.D. |
| N | 706 | 600 | 1270 | 608 | 1286 | 1235 | 977 | N.D. |
| Shannon (S) | 2.65 | 2.70 | 2.75 | 2.24 | 2.67 | 2.65 | 2.48 | N.D. |
| Margalef’s index | 2.43 | 2.34 | 2.65 | 1.46 | 2.37 | 2.10 | 1.80 | N.D. |
| Evenness index (e) | 0.156 | 0.169 | 0.130 | 0.094 | 0.153 | 0.165 | 0.170 | N.D. |
| Band | Close Relative in Database (Accession Number) | Taxonomic Description | Similarity (%) |
|---|---|---|---|
| B | Halogranum sp. SS5-1 (JN196479.1) | Euryarchaeota | 94 |
| C | Uncultured Halorhabdus sp. clone M3-B02 (KF452246) | 93 | |
| D | Uncultured archaeon clone WN-UWA-55 (DQ432585.1) | 96 | |
| E | Uncultured archaeon clone WN-UWA-55 (DQ432585.1) | 97 | |
| F | Uncultured archaeon clone CBA3919.b1 (JX881146.1) | 97 | |
| I | Uncultured archaeon clone WN-FWA-110 (DQ432514) | 99 | |
| J | Halalkalicoccus tibetensis: JCM 11,890 (AB663349) | 98 | |
| K | Halalkalicoccus jeotgali B3 (NR_102920.1) | 95 | |
| L | Halalkalicoccus jeotgali strain: JCM 14,584 (AB477223) | 98 | |
| M | Uncultured haloarchaeon clone TX4CA_31 (EF690586.1) | 97 | |
| N | Uncultured Halorubrum sp. isolate DGGE gel band JG06 (HQ110063) | 97 | |
| O | Archaeon DSFBPS_UR1C (KC465572) | Unclassified Archaea | 97 |
| P | Archaeon DSFBPG_5R3A (KC465564) | 99 | |
| Q | Halobacteriaceae archaeon YC93 (JQ237117.1) | Euryarchaeota | 90 |
| R | Uncultured euryarchaeote clone FR-M-R-S1-B11 (KC661808) | 92 | |
| S | Uncultured haloarchaeon clone TX4CA_24 (EF690579) | 98 | |
| T | Uncultured haloarchaeon clone ZB-A56 (AF505709) | 98 | |
| U | Uncultured archaeon clone ARC182 (JN185055) | 97 | |
| V | Uncultured haloarchaeon clone TX4CA_24 (EF690579) | 93 | |
| W | Uncultured crenarchaeote clone P1A2RS32 (FN666101) | Crenarchaeota | 97 |
| HS2A | Uncultured haloarchaeon clone HPA-18 (AY430113.1) | Euryarchaeota | 93 |
| HS2B | Uncultured haloarchaeon isolate DGGE gel band ESSA-A1_12 (KF309695) | 98 | |
| HS2C | Natronorubrum aibiense strain G23 (JQ522950) | 98 | |
| HS2D | Uncultured archaeon clone WN-USA-38 (DQ432550) | 94 | |
| FSA | Uncultured archaeon clone WN-USA-38(DQ432550) | 94 | |
| FSB | Uncultured archaeon clone P11_3-7C (KF814487) | 98 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elshafey, N.; Selim, S.; Mohammed, A.H.; Hagagy, N.; Samy, M.; Mostafa, E.M.; Safhi, F.A.; Alshamrani, S.M.; Saddiq, A.; Alsharari, S.S.; et al. Mapping Archaeal Diversity in Soda Lakes by Coupling 16S rRNA PCR-DGGE Analysis with Remote Sensing and GIS Technology. Fermentation 2022, 8, 365. https://doi.org/10.3390/fermentation8080365
Elshafey N, Selim S, Mohammed AH, Hagagy N, Samy M, Mostafa EM, Safhi FA, Alshamrani SM, Saddiq A, Alsharari SS, et al. Mapping Archaeal Diversity in Soda Lakes by Coupling 16S rRNA PCR-DGGE Analysis with Remote Sensing and GIS Technology. Fermentation. 2022; 8(8):365. https://doi.org/10.3390/fermentation8080365
Chicago/Turabian StyleElshafey, Naglaa, Samy Selim, Asmaa H. Mohammed, Nashwa Hagagy, Mennatalla Samy, Ehab M. Mostafa, Fatmah A. Safhi, Salha M. Alshamrani, Amna Saddiq, Salam S. Alsharari, and et al. 2022. "Mapping Archaeal Diversity in Soda Lakes by Coupling 16S rRNA PCR-DGGE Analysis with Remote Sensing and GIS Technology" Fermentation 8, no. 8: 365. https://doi.org/10.3390/fermentation8080365