
Fruit Microbial Communities of the Bisucciu Sardinian Apricot Cultivar (Prunus armeniaca L.) as a Reservoir of New Brewing Starter Strains

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fruit Sampling, Apricot Puree, and Malt Wort Preparation
2.2. Isolation of Fungal and Bacterial Communities of Apricot Fruits
2.3. Identification of Fungi and Bacteria Isolated from Apricot Fruits
2.4. Fermentation Trials
2.5. Volatiles Profile of Fermented Substrates
2.6. Statistical Analysis
3. Results
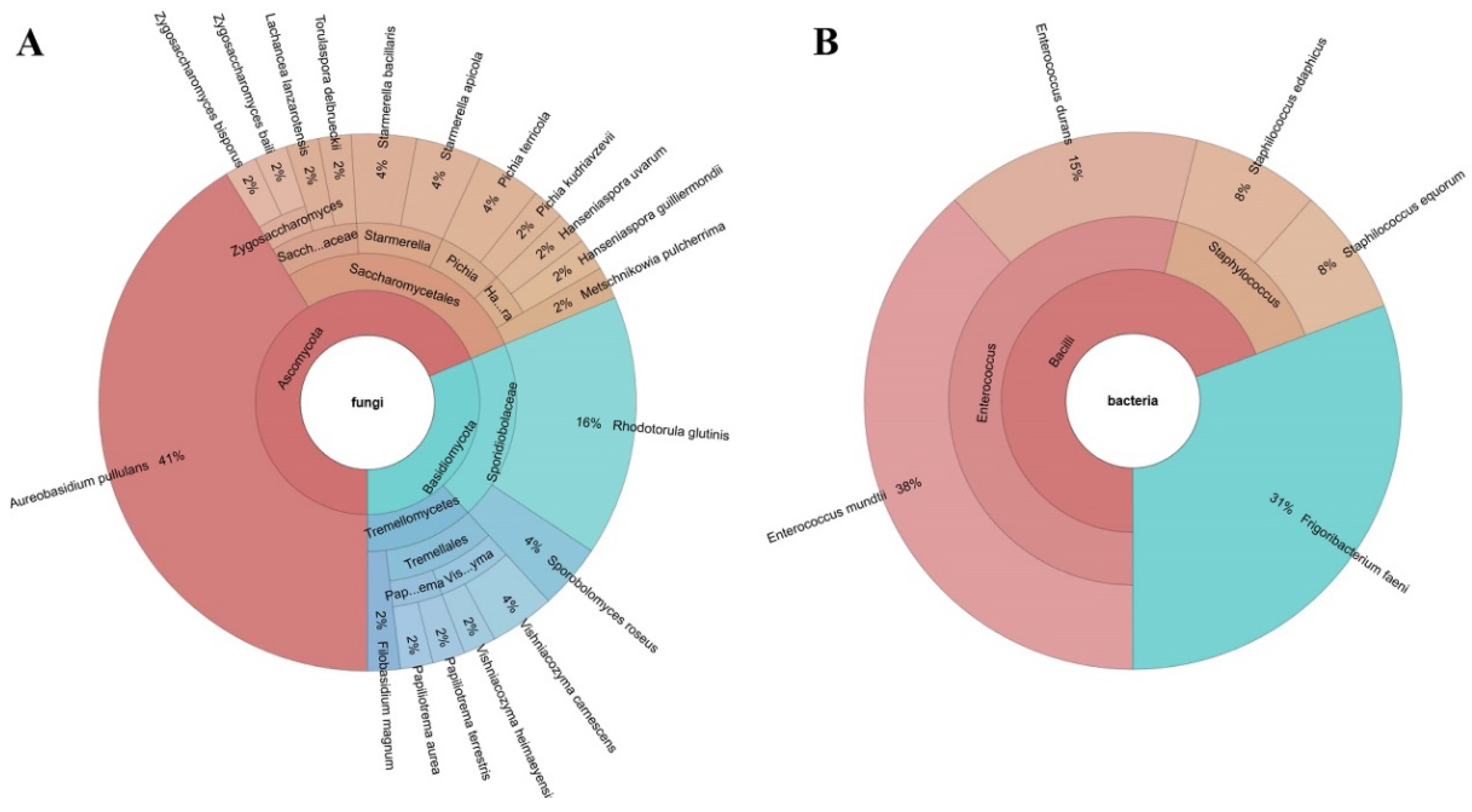
3.1. Bacterial and Fungal Communities of Bisucciu Apricot Fruits
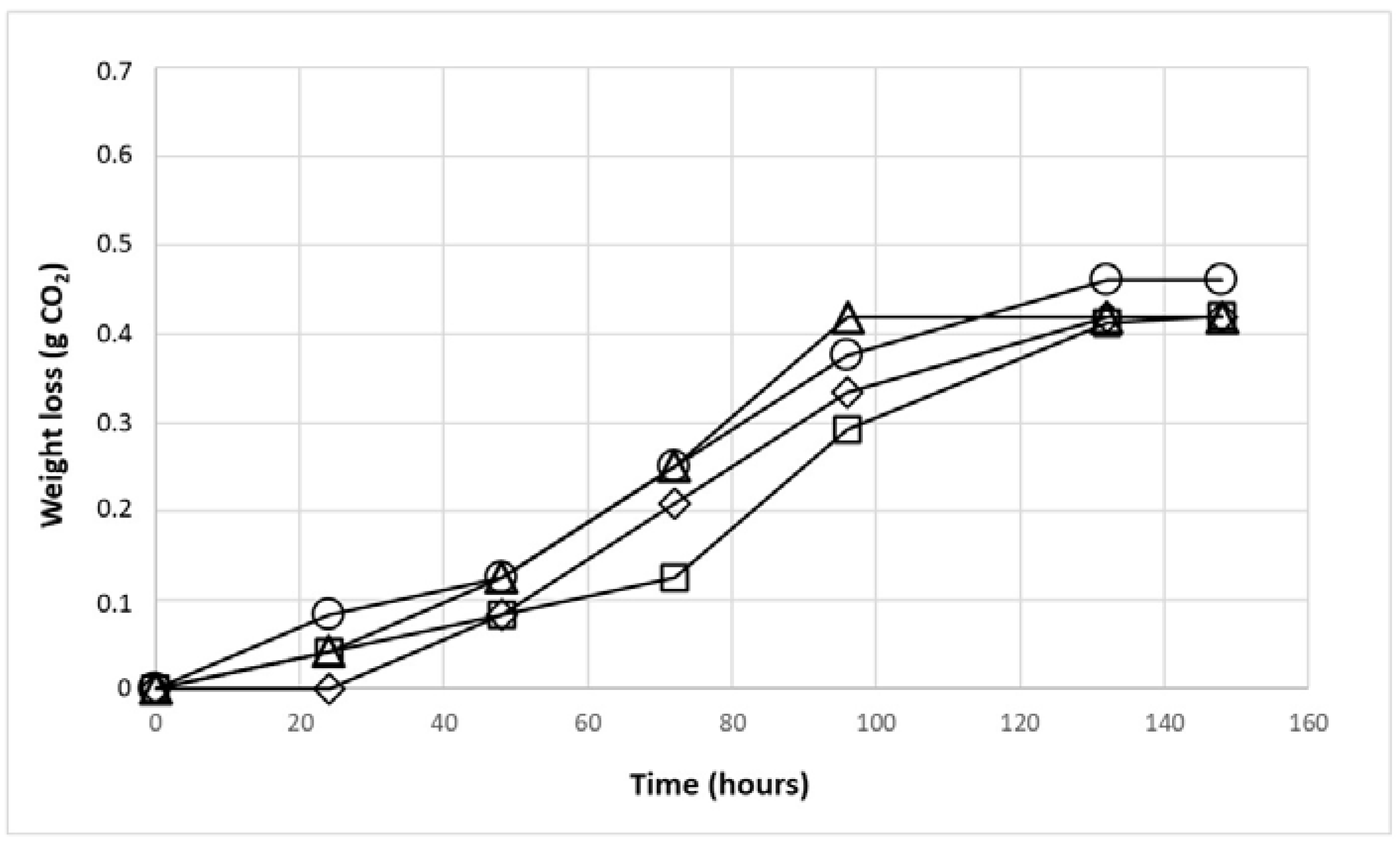
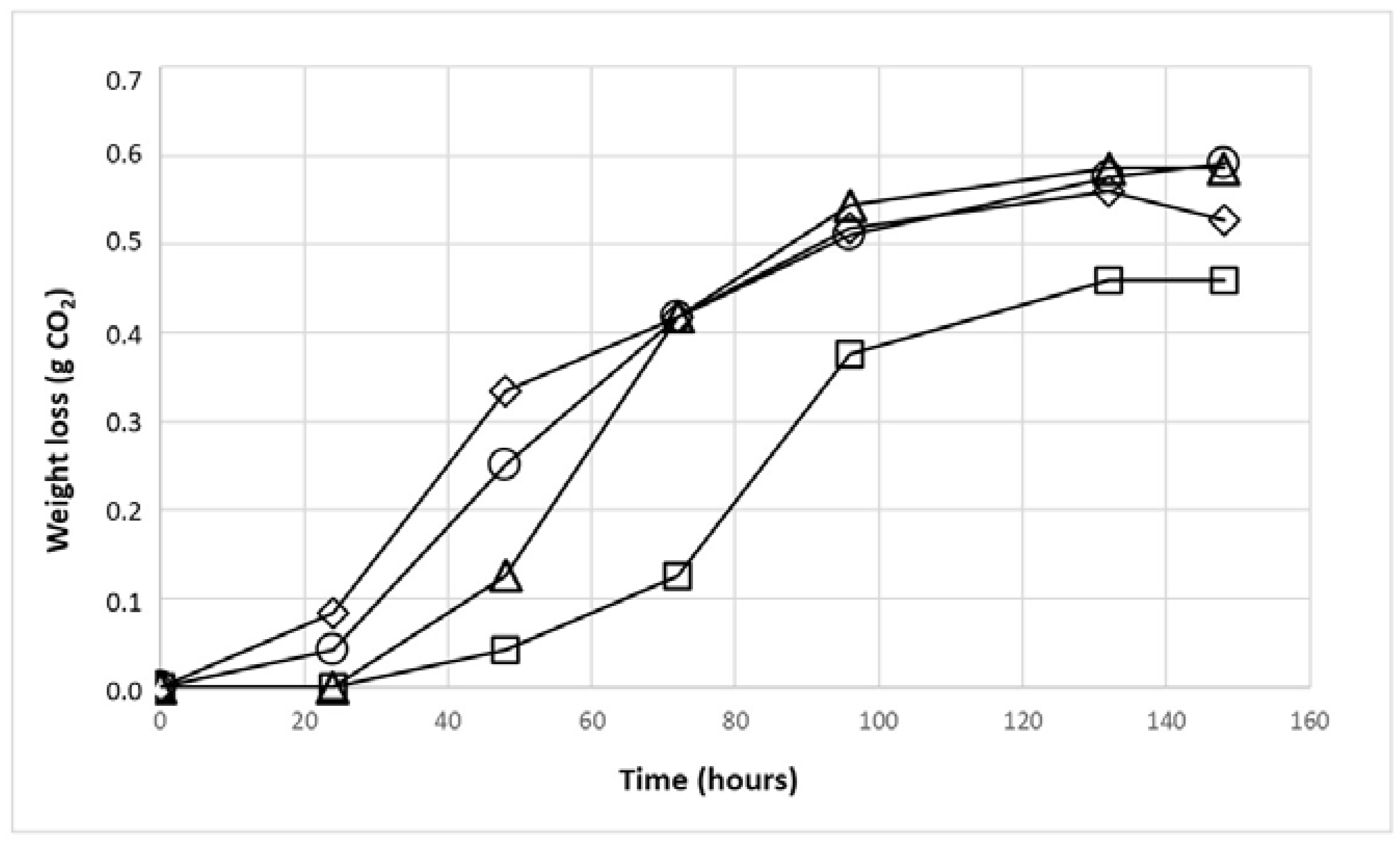
3.2. Fermentative Ability of Yeast Isolated from the Apricot Microbiota
4. Discussion
4.1. Bacterial and Fungal Communities Isolated from Bisucciu Apricot Fruit
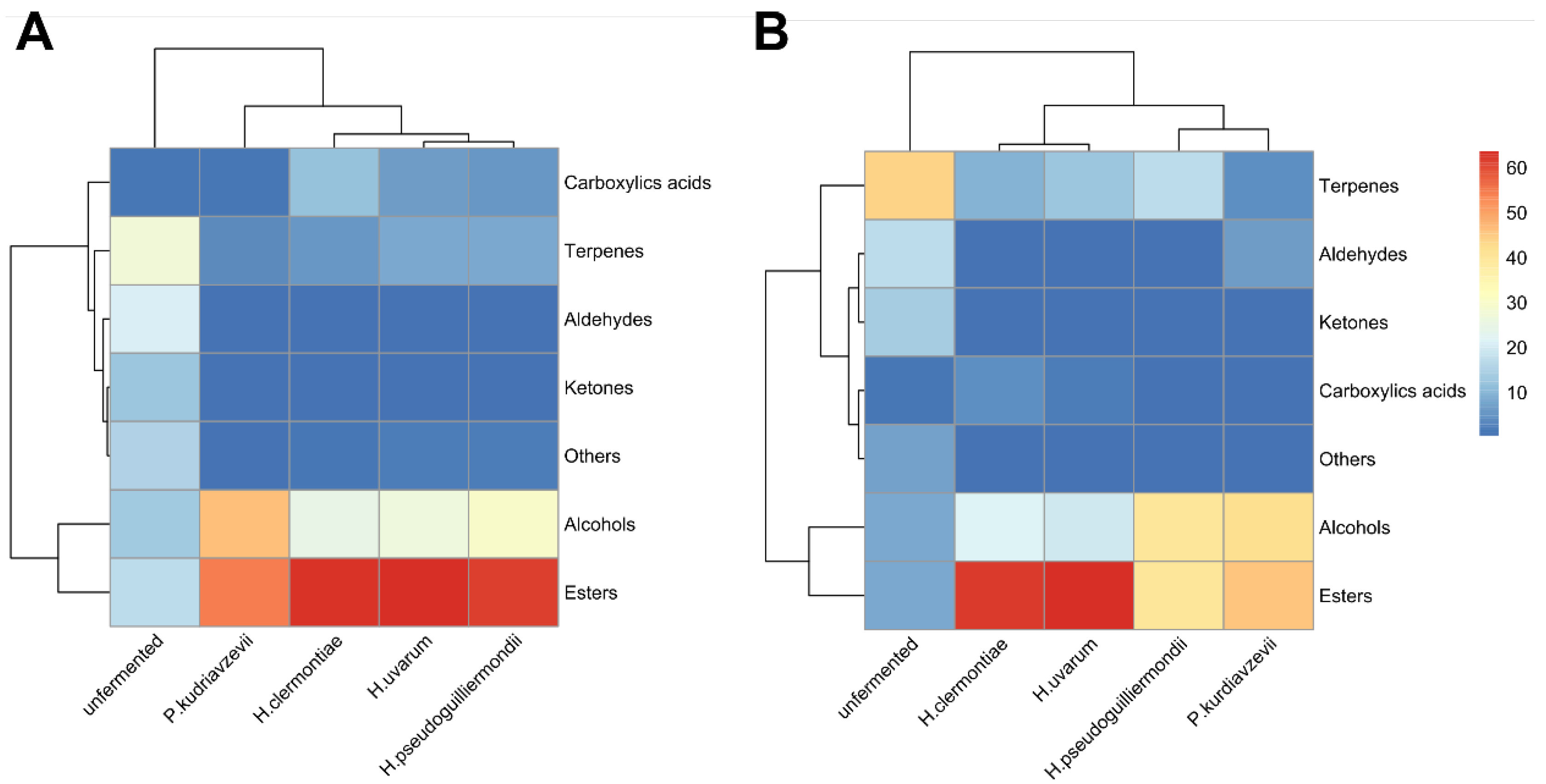
4.2. Yeast Isolated from Apricot Fruits as Potential Starters for Beer Production
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aquilani, B.; Laureti, T.; Poponi, S.; Secondi, L. Beer Choice and Consumption Determinants When Craft Beers Are Tasted: An Exploratory Study of Consumer Preferences. Food Qual. Prefer. 2015, 41, 214–224. [Google Scholar] [CrossRef]
- Fanari, M.; Forteschi, M.; Sanna, M.; Piu, P.P.; Porcu, M.C.; D’hallewin, G.; Secchi, N.; Zinellu, M.; Pretti, L. Pilot Plant Production of Craft Fruit Beer Using Ohmic-Treated Fruit Puree. J. Food Process. Preserv. 2020, 44, e14339. [Google Scholar] [CrossRef]
- McGovern, P.; Jalabadze, M.; Batiuk, S.; Callahan, M.P.; Smith, K.E.; Hall, G.R.; Kvavadze, E.; Maghradze, D.; Rusishvili, N.; Bouby, L.; et al. Early Neolithic Wine of Georgia in the South Caucasus. Proc. Natl. Acad. Sci. USA 2017, 114, E10309–E10318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helmold, M. Pricing Strategy in the Beer Industries. In Total Revenue Management (TRM): Case Studies, Best Practices and Industry Insights; Helmold, M., Ed.; Management for Professionals; Springer International Publishing: Cham, Switzerland, 2020; pp. 161–174. ISBN 978-3-030-46985-6. [Google Scholar]
- Koren, D.; Hegyesné Vecseri, B.; Kun-Farkas, G.; Urbin, Á.; Nyitrai, Á.; Sipos, L. How to Objectively Determine the Color of Beer? J. Food Sci. Technol. 2020, 57, 1183–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melewar, T.C.; Skinner, H. Territorial Brand Management: Beer, Authenticity, and Sense of Place. J. Bus. Res. 2020, 116, 680–689. [Google Scholar] [CrossRef]
- Suiker, I.M.; Wösten, H.A. Spoilage Yeasts in Beer and Beer Products. Curr. Opin. Food Sci. 2022, 44, 100815. [Google Scholar] [CrossRef]
- Bianco, A.; Fancello, F.; Balmas, V.; Zara, G.; Dettori, M.; Budroni, M. The Microbiome of Sardinian Barley and Malt. J. Inst. Brew. 2018, 124, 344–351. [Google Scholar] [CrossRef] [Green Version]
- Droby, S.; Wisniewski, M. The Fruit Microbiome: A New Frontier for Postharvest Biocontrol and Postharvest Biology. Postharvest Biol. Technol. 2018, 140, 107–112. [Google Scholar] [CrossRef]
- Bianco, A.; Fancello, F.; Balmas, V.; Dettori, M.; Motroni, A.; Zara, G.; Budroni, M. Microbial Communities and Malt Quality of Durum Wheat Used in Brewing. J. Inst. Brew. 2019, 125, 222–229. [Google Scholar] [CrossRef]
- Capece, A.; Romaniello, R.; Siesto, G.; Romano, P. Conventional and Non-Conventional Yeasts in Beer Production. Fermentation 2018, 4, 38. [Google Scholar] [CrossRef] [Green Version]
- Holt, S.; Mukherjee, V.; Lievens, B.; Verstrepen, K.J.; Thevelein, J.M. Bioflavoring by Non-Conventional Yeasts in Sequential Beer Fermentations. Food Microbiol. 2018, 72, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Canonico, L.; Galli, E.; Ciani, E.; Comitini, F.; Ciani, M. Exploitation of Three Non-Conventional Yeast Species in the Brewing Process. Microorganisms 2019, 7, E11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zara, G.; Mannazzu, I.; Del Caro, A.; Budroni, M.; Pinna, B.; Murru, M.; Farris, G.; Zara, S. Wine Quality Improvement through the Combined Utilisation of Yeast Hulls and Candida zemplinina/Saccharomyces cerevisiae Mixed Starter Cultures. Aust. J. Grape Wine Res. 2014, 20, 199–207. [Google Scholar] [CrossRef]
- Xiang, H.; Sun-Waterhouse, D.; Waterhouse, G.I.N.; Cui, C.; Ruan, Z. Fermentation-Enabled Wellness Foods: A Fresh Perspective. Food Sci. Hum. Wellness 2019, 8, 203–243. [Google Scholar] [CrossRef]
- Lindgren, S.E.; Dobrogosz, W.J. Antagonistic Activities of Lactic Acid Bacteria in Food and Feed Fermentations. FEMS Microbioliol. Rev. 1990, 7, 149–163. [Google Scholar] [CrossRef]
- Marongiu, A.; Zara, G.; Legras, J.L.; Del Caro, A.; Mascia, I.; Fadda, C.; Budroni, M. Novel Starters for Old Processes: Use of Saccharomyces cerevisiae Strains Isolated from Artisanal Sourdough for Craft Beer Production at a Brewery Scale. J. Ind. Microbiol. Biotechnol. 2015, 42, 85–92. [Google Scholar] [CrossRef]
- Mascia, I.; Fadda, C.; Dostálek, P.; Karabín, M.; Zara, G.; Budroni, M.; Del Caro, A. Is It Possible to Create an Innovative Craft Durum Wheat Beer with Sourdough Yeasts? A Case Study. J. Inst. Brew. 2015, 121, 283–286. [Google Scholar] [CrossRef]
- Abe Sato, S.T.; Marques, J.M.; da Luz de Freitas, A.; Sanches Progênio, R.C.; Nunes, M.R.T.; Mota de Vasconcelos Massafra, J.; Gomes Moura, F.; Rogez, H. Isolation and Genetic Identification of Endophytic Lactic Acid Bacteria from the Amazonian Açai Fruits: Probiotics Features of Selected Strains and Their Potential to Inhibit Pathogens. Front. Microbiol. 2021, 11, 610524. [Google Scholar] [CrossRef]
- Filannino, P.; Di Cagno, R.; Addante, R.; Pontonio, E.; Gobbetti, M. Metabolism of Fructophilic Lactic Acid Bacteria Isolated from the Apis Mellifera, L. Bee Gut: Phenolic Acids as External Electron Acceptors. Appl. Environ. Microbiol. 2016, 82, 6899–6911. [Google Scholar] [CrossRef] [Green Version]
- Masi, C.; Gemechu, G.; Tafesse, M. Isolation, Screening, Characterization, and Identification of Alkaline Protease-Producing Bacteria from Leather Industry Effluent. Ann. Microbioly 2021, 71, 24. [Google Scholar] [CrossRef]
- Stirling, D. DNA Extraction from Fungi, Yeast, and Bacteria. In PCR Protocols; Bartlett, J.M.S., Stirling, D., Eds.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2003; pp. 53–54. ISBN 978-1-59259-384-2. [Google Scholar]
- Godon, J.J.; Zumstein, E.; Dabert, P.; Habouzit, F.; Moletta, R. Molecular Microbial Diversity of an Anaerobic Digestor as Determined by Small-Subunit RDNA Sequence Analysis. Appl. Environ. Microbiol. 1997, 63, 2802–2813. [Google Scholar] [CrossRef] [Green Version]
- Esteve-Zarzoso, B.; Belloch, C.; Uruburu, F.; Querol, A. Identification of Yeasts by RFLP Analysis of the 5.8S RRNA Gene and the Two Ribosomal Internal Transcribed Spacers. Int. J. Syst. Bacteriol. 1999, 49 (Pt. 1), 329–337. [Google Scholar] [CrossRef] [Green Version]
- Ciani, M.; Maccarelli, F. Oenological Properties of Non-Saccharomyces Yeasts Associated with Wine-Making. World J. Microbiol. Biotechnol. 1997, 14, 199–203. [Google Scholar] [CrossRef]
- Fanari, M.; Forteschi, M.; Sanna, M.; Zinellu, M.; Porcu, M.C.; Pretti, L. Comparison of Enzymatic and Precipitation Treatments for Gluten-Free Craft Beers Production. Innov. Food Sci. Emerg. Technol. 2018, 49, 76–81. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. Available online: https://www.R-project.org/ (accessed on 25 June 2022).
- Badosa, E.; Trias, R.; Parés, D.; Pla, M.; Montesinos, E. Microbiological Quality of Fresh Fruit and Vegetable Products in Catalonia (Spain) Using Normalised Plate-Counting Methods and Real Time Polymerase Chain Reaction (QPCR). J. Sci. Food Agric. 2008, 88, 605–611. [Google Scholar] [CrossRef]
- Leff, J.W.; Fierer, N. Bacterial Communities Associated with the Surfaces of Fresh Fruits and Vegetables. PLoS ONE 2013, 8, e59310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.M.; Azad, M.O.K.; Uddain, J.; Adnan, M.; Ali, M.C.; Al-Mujahidy, S.K.M.J.; Roni, M.Z.K.; Rahman, M.S.; Islam, M.J.; Rahman, M.H.; et al. Microbial Quality Assessment and Efficacy of Low-Cost Disinfectants on Fresh Fruits and Vegetables Collected from Urban Areas of Dhaka, Bangladesh. Foods 2021, 10, 1325. [Google Scholar] [CrossRef] [PubMed]
- Vadkertiová, R.; Molnárová, J.; Vránová, D.; Sláviková, E. Yeasts and Yeast-like Organisms Associated with Fruits and Blossoms of Different Fruit Trees. Can. J. Microbiol. 2012, 58, 1344–1352. [Google Scholar] [CrossRef]
- Bösch, Y.; Britt, E.; Perren, S.; Naef, A.; Frey, J.E.; Bühlmann, A. Dynamics of the Apple Fruit Microbiome after Harvest and Implications for Fruit Quality. Microorganisms 2021, 9, 272. [Google Scholar] [CrossRef]
- Abdelfattah, A.; Wisniewski, M.; Droby, S.; Schena, L. Spatial and Compositional Variation in the Fungal Communities of Organic and Conventionally Grown Apple Fruit at the Consumer Point-of-Purchase. Hortic. Res. 2016, 3, 16047. [Google Scholar] [CrossRef] [Green Version]
- Wisniewski, M.; Droby, S.; Norelli, J.; Liu, J.; Schena, L. Alternative Management Technologies for Postharvest Disease Control: The Journey from Simplicity to Complexity. Postharvest Biol. Technol. 2016, 122, 3–10. [Google Scholar] [CrossRef]
- Kecskeméti, E.; Berkelmann-Löhnertz, B.; Reineke, A. Are Epiphytic Microbial Communities in the Carposphere of Ripening Grape Clusters (Vitis Vinifera L.) Different between Conventional, Organic, and Biodynamic Grapes? PLoS ONE 2016, 11, e0160852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenea, G.N.; Jarrin-V, P.; Yepez, L. Microbiota of Wild Fruits from the Amazon Region of Ecuador: Linking Diversity and Functional Potential of Lactic Acid Bacteria with Their Origin; IntechOpen Limited: London, UK, 2020; ISBN 978-1-83962-813-9. [Google Scholar]
- Jeong, D.-W.; Heo, S.; Ryu, S.; Blom, J.; Lee, J.H. Genomic Insights into the Virulence and Salt Tolerance of Staphylococcus equorum. Sci. Rep. 2017, 7, 5383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franz, C.M.A.P.; Huch, M.; Abriouel, H.; Holzapfel, W.; Gálvez, A. Enterococci as Probiotics and Their Implications in Food Safety. Int. J. Food Microbiol. 2011, 151, 125–140. [Google Scholar] [CrossRef] [Green Version]
- Barata, A.; Malfeito-Ferreira, M.; Loureiro, V. The Microbial Ecology of Wine Grape Berries. Int. J. Food Microbiol. 2012, 153, 243–259. [Google Scholar] [CrossRef]
- Lukša, J.; Vepštaitė-Monstavičė, I.; Apšegaitė, V.; Blažytė-Čereškienė, L.; Stanevičienė, R.; Strazdaitė-Žielienė, Ž.; Ravoitytė, B.; Aleknavičius, D.; Būda, V.; Mozūraitis, R.; et al. Fungal Microbiota of Sea Buckthorn Berries at Two Ripening Stages and Volatile Profiling of Potential Biocontrol Yeasts. Microorganisms 2020, 8, E456. [Google Scholar] [CrossRef] [Green Version]
- Isaeva, O.V.; Glushakova, A.M.; Garbuz, S.A.; Kachalkin, A.V.; Chernov, I.I. Endophytic yeast fungi in plant storage tissues. Biol. Bull. Russ. Acad. Sci. 2010, 37, 26–34. [Google Scholar] [CrossRef]
- Gramisci, B.R.; Lutz, M.C.; Lopes, C.A.; Sangorrín, M.P. Enhancing the Efficacy of Yeast Biocontrol Agents against Postharvest Pathogens through Nutrient Profiling and the Use of Other Additives. Biol. Control 2018, 121, 151–158. [Google Scholar] [CrossRef]
- Castoria, R.; Miccoli, C.; Barone, G.; Palmieri, D.; De Curtis, F.; Lima, G.; Heitman, J.; Ianiri, G. Molecular Tools for the Yeast Papiliotrema terrestris LS28 and Identification of Yap1 as a Transcription Factor Involved in Biocontrol Activity. Appl. Environ. Microbiol. 2021, 87, e02910–e02920. [Google Scholar] [CrossRef]
- Di Francesco, A.; Ugolini, L.; Lazzeri, L.; Mari, M. Production of Volatile Organic Compounds by Aureobasidium pullulans as a Potential Mechanism of Action against Postharvest Fruit Pathogens. Biol. Control 2015, 81, 8–14. [Google Scholar] [CrossRef]
- Zhang, D.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Efficacy of the Antagonist Aureobasidium pullulans PL5 against Postharvest Pathogens of Peach, Apple and Plum and Its Modes of Action. Biol. Control 2010, 54, 172–180. [Google Scholar] [CrossRef]
- Filonow, A.B.; Vishniac, H.S.; Anderson, J.A.; Janisiewicz, W.J. Biological Control of Botrytis cinerea in Apple by Yeasts from Various Habitats and Their Putative Mechanisms of Antagonism. Biol. Control 1996, 7, 212–220. [Google Scholar] [CrossRef]
- Janisiewicz, W.J.; Kurtzman, C.P.; Buyer, J.S. Yeasts Associated with Nectarines and Their Potential for Biological Control of Brown Rot. Yeast 2010, 27, 389–398. [Google Scholar] [CrossRef] [Green Version]
- Janisiewicz, W.J.; Tworkoski, T.J.; Kurtzman, C.P. Biocontrol Potential of Metchnikowia pulcherrima Strains Against Blue Mold of Apple. Phytopathology 2001, 91, 1098–1108. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Niu, C.; Liu, B.; Yuan, Y.; Yue, T. Identification and Characterization of Epiphytic Yeasts on Apples in China. RSC Adv. 2017, 7, 44766–44772. [Google Scholar] [CrossRef] [Green Version]
- Budroni, M.; Zara, G.; Ciani, M.; Comitini, F. Saccharomyces and Non-Saccharomyces Starter Yeasts. In Brewing Technology; Kanauchi, M., Ed.; IntechOpen: London, UK, 2017; Available online: https://www.intechopen.com/chapters/55278 (accessed on 25 June 2022).
- Chanprasartsuk, O.; Prakitchaiwattana, C.; Sanguandeekul, R.; Fleet, G.H. Autochthonous Yeasts Associated with Mature Pineapple Fruits, Freshly Crushed Juice and Their Ferments; and the Chemical Changes during Natural Fermentation. Bioresour. Technol. 2010, 101, 7500–7509. [Google Scholar] [CrossRef]
- Matos, Í.T.S.R.; de Souza, V.A.; D’Angelo, G.; do, R.; Astolfi Filho, S.; do Carmo, E.J.; Vital, M.J.S. Yeasts with Fermentative Potential Associated with Fruits of Camu-Camu (Myrciaria Dubia, Kunth) from North of Brazilian Amazon. Sci. World J. 2021, 2021, 9929059. [Google Scholar] [CrossRef] [PubMed]
- Nyanga, L.K.; Nout, M.J.R.; Gadaga, T.H.; Theelen, B.; Boekhout, T.; Zwietering, M.H. Yeasts and Lactic Acid Bacteria Microbiota from Masau (Ziziphus Mauritiana) Fruits and Their Fermented Fruit Pulp in Zimbabwe. Int. J. Food Microbiol. 2007, 120, 159–166. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, S.T.; da Paz, M.F.; Altemio, Â.D.C. Evaluation of Two Brazilian Native Yeast Strains (Pichia kudriavzevii) in Craft Beer. Res. Soc. Dev. 2022, 11, e17311124783. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T. The Yeasts: A Taxonomic Study, 5th ed.; Elsevier Science: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Bellut, K.; Michel, M.; Zarnkow, M.; Hutzler, M.; Jacob, F.; De Schutter, D.P.; Daenen, L.; Lynch, K.M.; Zannini, E.; Arendt, E.K. Application of Non-Saccharomyces Yeasts Isolated from Kombucha in the Production of Alcohol-Free Beer. Fermentation 2018, 4, 66. [Google Scholar] [CrossRef] [Green Version]
- Matraxia, M.; Alfonzo, A.; Prestianni, R.; Francesca, N.; Gaglio, R.; Todaro, A.; Alfeo, V.; Perretti, G.; Columba, P.; Settanni, L.; et al. Non-Conventional Yeasts from Fermented Honey by-Products: Focus on Hanseniaspora uvarum Strains for Craft Beer Production. Food Microbiol. 2021, 99, 103806. [Google Scholar] [CrossRef] [PubMed]
- King, A.J.; Dickinson, J.R. Biotransformation of Hop Aroma Terpenoids by Ale and Lager Yeasts. FEMS Yeast Res. 2003, 3, 53–62. [Google Scholar] [CrossRef]
- Carrau, F.M.; Medina, K.; Boido, E.; Farina, L.; Gaggero, C.; Dellacassa, E.; Versini, G.; Henschke, P.A. De Novo Synthesis of Monoterpenes by Saccharomyces cerevisiae Wine Yeasts. FEMS Microbiol. Lett. 2005, 243, 107–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holt, S.; Mikš, M.; de Carvalho, B.T.; Foulquié-Moreno, M.R.; Thevelein, J. The Molecular Biology of Fruity and Floral Aromas in Beer and Other Alcoholic Beverages. FEMS Microbiol. Rev. 2019, 43, 193–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microbial Groups | Isolation Media | Log 10 CFU/g |
|---|---|---|
| Total mesophilic count | PCA | 4.75 ± 4.58 |
| Fungal count | RBA | 4.44 ± 4.07 |
| Lactic acid bacteria | MRS | 3.63 ± 3.52 |
| Fructophilic lactic acid bacteria | FLAB | 2.37 ± 2.40 |
| W | WPP | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| C | L1 | L2 | L3 | L4 | C | L1 | L2 | L3 | L4 | |
| pH | 5.22 a ± 0.01 | 4.73 b ± 0.02 | 4.48 c ± 0.03 | 4.67 b ± 0.03 | 4.53 c ± 0.02 | 4.59 a ± 0.02 | 4.58 a ± 0.01 | 4.42 b ± 0.01 | 4.44 b ± 0.04 | 4.42 b ± 0.06 |
| °Brix | 13.25 a ± 0.12 | 12.37 b ± 0.06 | 12.48 b ± 0.51 | 12.49 b ± 0.42 | 12.49 b ± 0.19 | 13.55 a ± 0.27 | 12.21 c ± 0.12 | 12.47 bc ± 0.42 | 12.35 c ± 0.12 | 12.79 b ± 0.27 |
| Specific Gravity | 1.057 a ± 0.01 | 1.053 b ± 0.01 | 1.053 b ± 0.01 | 1.053 b ± 0.02 | 1.053 b ± 0.01 | 1.058 a ± 0.01 | 1.052 c ± 0.01 | 1.053 bc ± 0.01 | 1.052 c ± 0.00 | 1.054 b ± 0.01 |
| Ethanol (% vol/vol) | - | 0.59 a ± 0.01 | 0.52 ab ± 0.04 | 0.52 b ± 0.05 | 0.52 b ± 0.02 | - | 0.74 a ± 0.02 | 0.66 a ± 0.06 | 0.73 a ± 0.03 | 0.57 b ± 0.03 |
| Attenuation (%) | - | 7.9 a ± 0.1 | 6.9 b ± 0.2 | 6.9 b ± 0.1 | 7.0 b ± 0.1 | - | 9.9 a ± 0.2 | 8.8 b ± 0.2 | 9.8 a ± 0.1 | 7.7 c ± 0.1 |
| W | WPP | p-Value | |
|---|---|---|---|
| Carboxylics acids | 1.0420 ± 0.287 | 1.082 ± 0.469 | 0.869 |
| Alcohols * | 12.161 ± 0.246 | 8.344 ± 0.375 | 0.001 |
| Esters * | 16.198 ± 0.394 | 8.221 ± 0.089 | 0.002 |
| Aldehydes | 19.345 ± 0.538 | 16.990 ± 0.965 | 0.086 |
| Ketones * | 11.587 ± 0.117 | 14.326 ± 0.112 | 0.000 |
| Terpenes * | 25.127 ± 0.302 | 43.867 ± 0.869 | 0.001 |
| Others * | 14.192 ± 0.246 | 7.008 ± 0.662 | 0.003 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fancello, F.; Bianco, A.; Niccolai, M.; Zara, G.; Coronas, R.; Serra, E.; D’Hallewin, G.; Valentoni, A.; Santoru, A.; Pretti, L.; et al. Fruit Microbial Communities of the Bisucciu Sardinian Apricot Cultivar (Prunus armeniaca L.) as a Reservoir of New Brewing Starter Strains. Fermentation 2022, 8, 364. https://doi.org/10.3390/fermentation8080364
Fancello F, Bianco A, Niccolai M, Zara G, Coronas R, Serra E, D’Hallewin G, Valentoni A, Santoru A, Pretti L, et al. Fruit Microbial Communities of the Bisucciu Sardinian Apricot Cultivar (Prunus armeniaca L.) as a Reservoir of New Brewing Starter Strains. Fermentation. 2022; 8(8):364. https://doi.org/10.3390/fermentation8080364
Chicago/Turabian StyleFancello, Francesco, Angela Bianco, Marta Niccolai, Giacomo Zara, Roberta Coronas, Elisa Serra, Guy D’Hallewin, Antonio Valentoni, Antonio Santoru, Luca Pretti, and et al. 2022. "Fruit Microbial Communities of the Bisucciu Sardinian Apricot Cultivar (Prunus armeniaca L.) as a Reservoir of New Brewing Starter Strains" Fermentation 8, no. 8: 364. https://doi.org/10.3390/fermentation8080364