
The Impact of Indigenous Non-Saccharomyces Yeasts Inoculated Fermentations on ‘Semillon’ Icewine



 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Culture Media
2.2. Chemicals and Standards
2.3. Icewine Samples Fermentation Trials
2.4. Oenological Parameters Analysis
2.5. Fermentation Kinetics and Yeast Biomass
2.6. Volatile Aroma Compounds Analysis
2.7. Sensory Analysis
2.8. Data Analysis
3. Results and Discussion
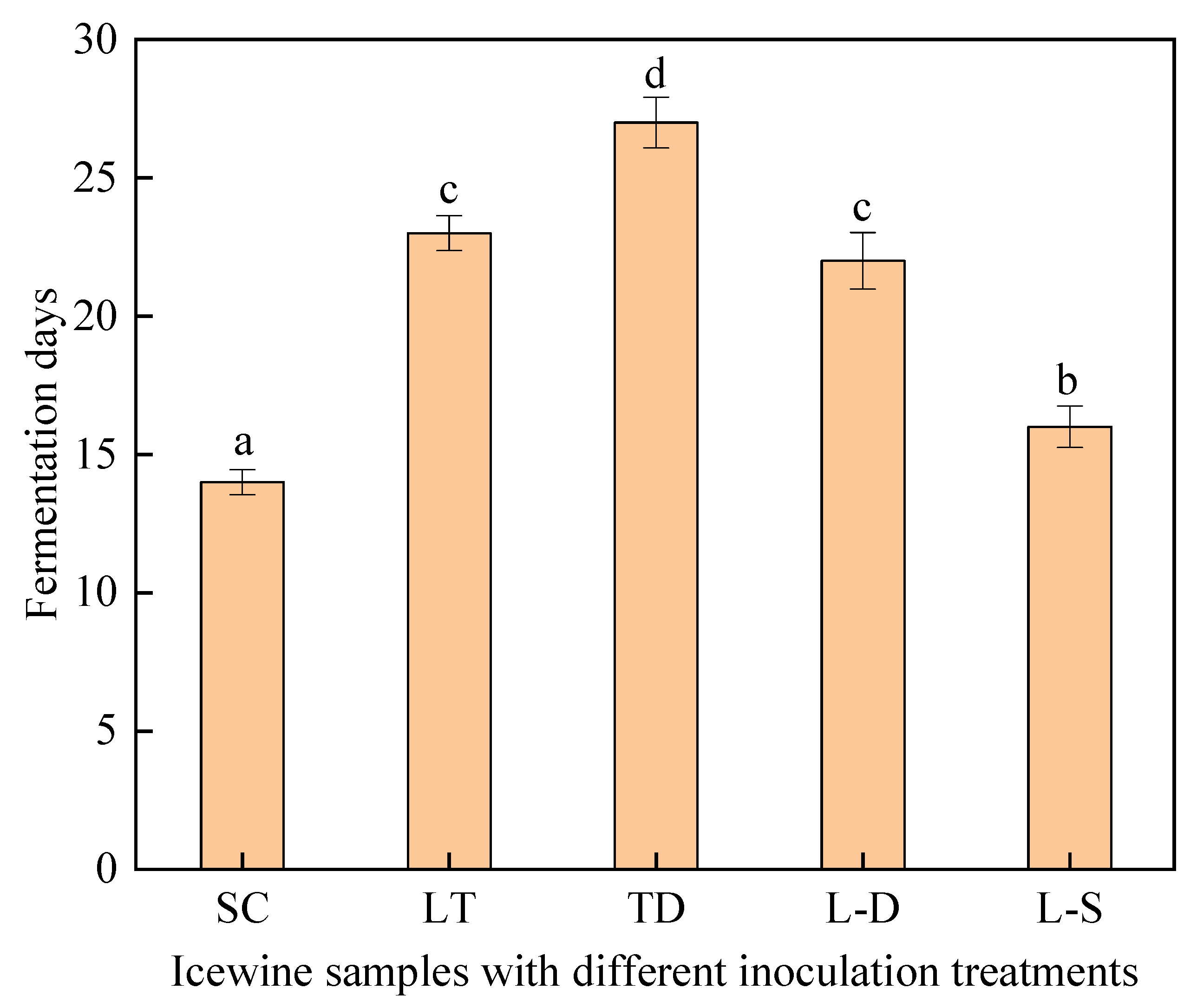
3.1. Fermentation Process
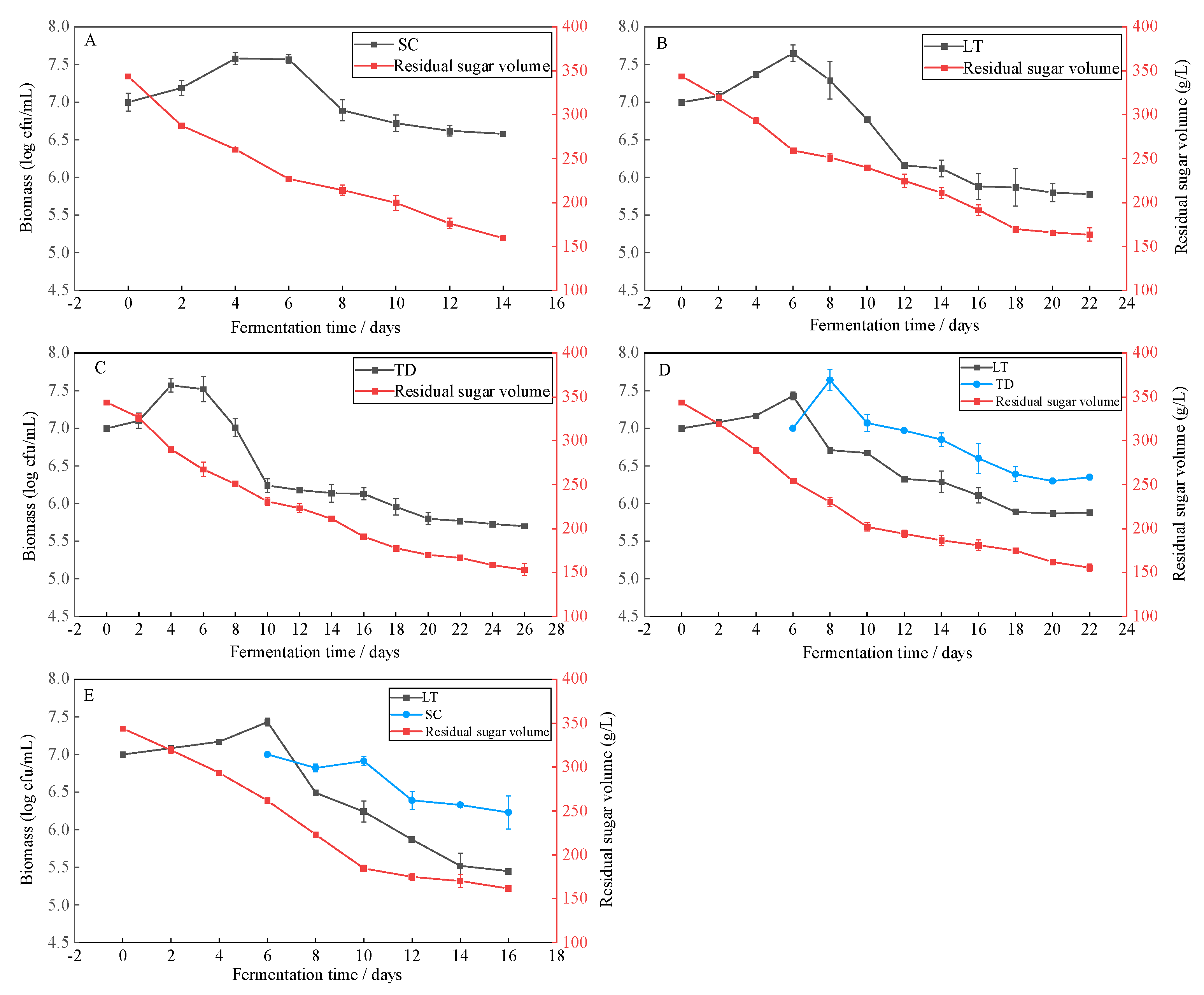
3.2. Yeast Population Dynamics and the Sugar Consumption Trend
3.3. Oenological Parameters Analysis
3.4. Analysis of the Main Volatile Compounds in icewines
3.4.1. Esters
3.4.2. Higher alcohols
3.4.3. Acids
3.4.4. Terpenes
3.4.5. Others
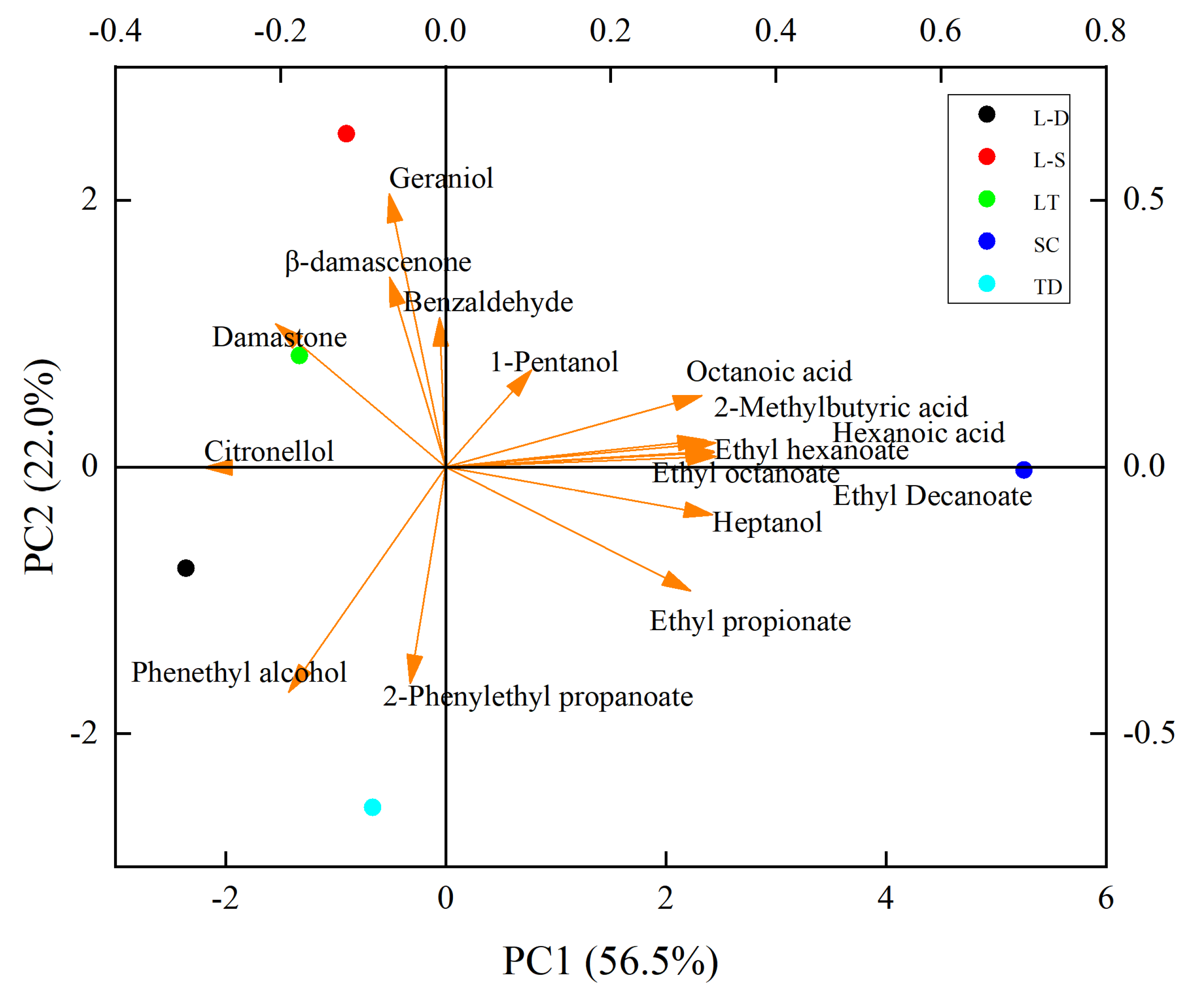
3.5. Principal Component Analysis of Volatile Aroma Compounds
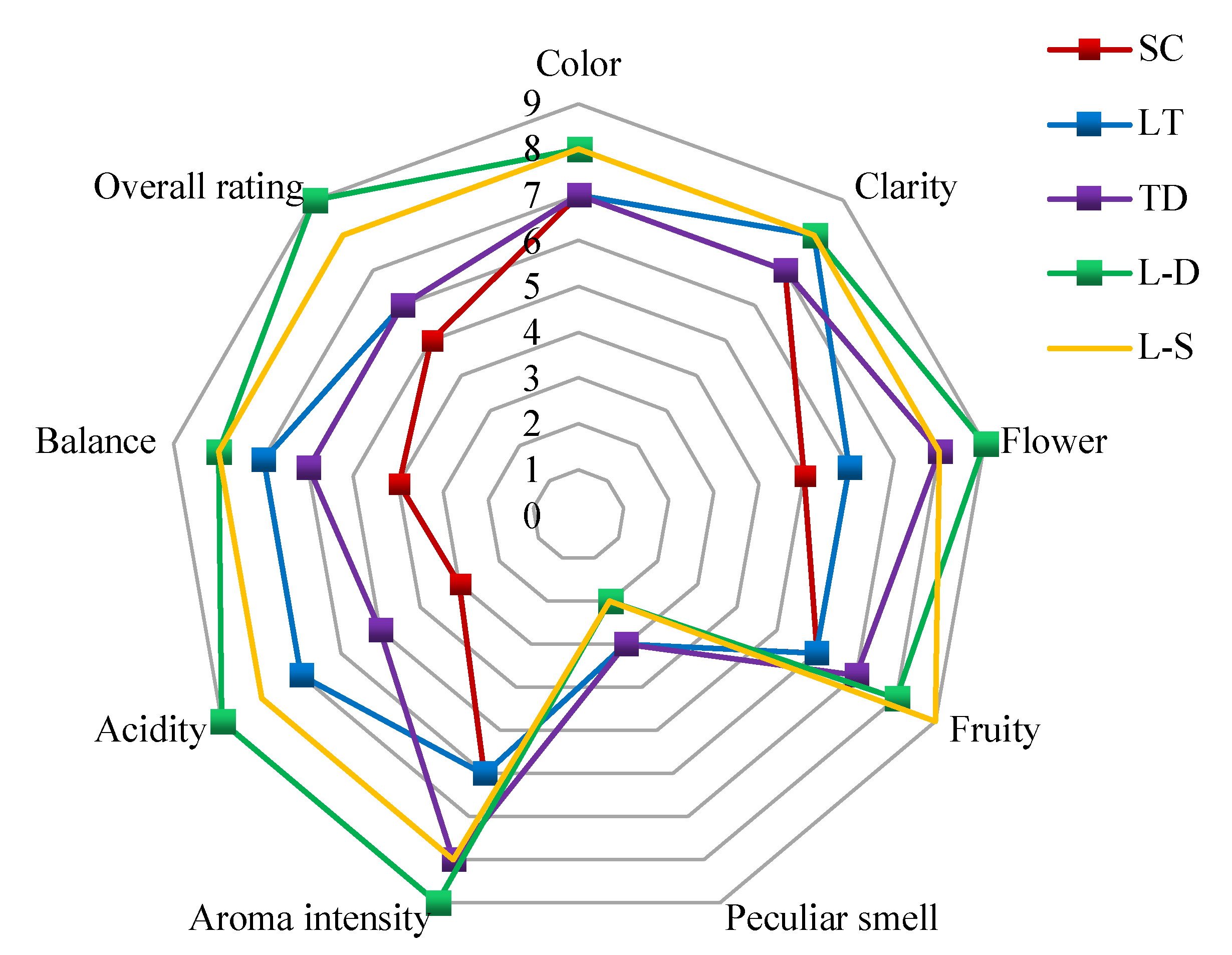
3.6. Sensory Evaluation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, Y.; Xu, Y.; Tang, K. Aroma of icewine: A review on how environmental, viticultural, and oenological factors affect the aroma of icewine. J. Agric. Food Chem. 2021, 69, 6943–6957. [Google Scholar] [CrossRef] [PubMed]
- Ostapenko, V. Analysis on application of different grape varieties in the production of icewine. A review. Ukr. Food J. 2016, 5, 678–694. [Google Scholar] [CrossRef]
- Wang, J.; Li, M.; Li, J.; Ma, T.; Han, S.; Antonio, M.; Suárez Lepe, J.A. Biotechnology of Ice Wine Production. In Advances in Biotechnology for Food Industry; Elsevier: Amsterdam, The Netherlands, 2018; Volume 14, pp. 267–300. [Google Scholar]
- Borren, E.; Tian, B. The important contribution of non-Saccharomyces yeasts to the aroma complexity of wine: A review. Foods 2021, 10, 13. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, W.; Yi, H.; Wang, B.; Xiao, J.; Zhou, X.; Jiankun, X.; Jiang, L.; Shi, X. Microbial community composition and its role in volatile compound formation during the spontaneous fermentation of ice wine made from Vidal grapes. Process Biochem. 2020, 92, 365–377. [Google Scholar] [CrossRef]
- Pigeau, G.M.; Bozza, E.; Kaiser, K.; Inglis, D.L. Concentration effect of Riesling Icewine juice on yeast performance and wine acidity. J. Appl. Microbiol. 2007, 103, 1691–1698. [Google Scholar] [CrossRef] [Green Version]
- Canonico, L.; Agarbati, A.; Comitini, F.; Ciani, M. Torulaspora delbrueckii in the brewing process: A new approach to enhance bioflavour and to reduce ethanol content. Food Microbiol. 2016, 56, 45–51. [Google Scholar] [CrossRef]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Azzolini, M.; Tosi, E.; Lorenzini, M.; Finato, F.; Zapparoli, G. Contribution to the aroma of white wines by controlled Torulaspora delbrueckii cultures in association with Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2015, 31, 277–293. [Google Scholar] [CrossRef]
- Belda, I.; Ruiz, J.; Beisert, B.; Navascués, E.; Marquina, D.; Calderón, F.; Rauhut, D.; Benito, S.; Santos, A. Influence of Torulaspora delbrueckii in varietal thiol (3-SH and 4-MSP) release in wine sequential fermentations. Int. J. Food Microbiol. 2017, 257, 183–191. [Google Scholar] [CrossRef]
- Bely, M.; Stoeckle, P.; Masneuf-Pomarède, I.; Dubourdieu, D. Impact of mixed Torulaspora delbrueckii–Saccharomyces cerevisiae culture on high-sugar fermentation. Int. J. Food Microbiol. 2008, 122, 312–320. [Google Scholar] [CrossRef]
- Benito, S. The impacts of Lachancea thermotolerans yeast strains on winemaking. Appl. Microbiol. Biotechnol. 2018, 102, 6775–6790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 2013, 33, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Lu, Y.; Liu, S.Q. Effects of different yeasts on physicochemical and oenological properties of red dragon fruit wine fermented with Saccharomyces cerevisiae, Torulaspora delbrueckii and Lachancea thermotolerans. Microorganisms 2020, 8, 315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berbegal, C.; Fragasso, M.; Russo, P.; Bimbo, F.; Grieco, F.; Spano, G.; Capozzi, V. Climate changes and food quality: The potential of microbial activities as mitigating strategies in the wine sector. Fermentation 2019, 5, 85. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Jia, M.W.; Dong, Q.Y.; Yu, Y.S.; Yi, Q.; Yan, L.L. Yeast population dynamics during spontaneous fermentation of icewine and selection of indigenous Saccharomyces cerevisiae strains for the winemaking in Qilian, China. Food Agric. 2020, 100, 5385–5394. [Google Scholar]
- Benito, S.; Hofmann, T.; Laier, M.; Lochbühler, B.; Schüttler, A.; Ebert, K.; Fritsch, S.; Röcker, J.; Rauhut, D. Effect on quality and composition of Riesling wines fermented by sequential inoculation with non-Saccharomyces and Saccharomyces cerevisiae. Eur. Food Res. Technol. 2015, 241, 707–717. [Google Scholar] [CrossRef]
- Hu, K.; Jin, G.J.; Xu, Y.H.; Tao, Y.S. Wine aroma response to different participation of selected Hanseniaspora uvarum in mixed fermentation with Saccharomyces cerevisiae. Food Res. Int. 2018, 108, 119–127. [Google Scholar] [CrossRef]
- OIV-MA-AS313-01: R2015; Compendium of International Methods of Wine and Must Analysis. Chemical Analysis: Acids, Total Acidity (Oeno 551/2015). Organisation Internationale de la Vigne et du Vin: France, Paris, 2015.
- Pérez-Ruiz, T.; Martínez-Lozano, C.; Tomás, V.; Martín, J. High-performance liquid chromatographic separation and quantification of citric, lactic, malic, oxalic and tartaric acids using a post-column photochemical reaction and chemiluminescence detection. J. Chromatogr. A 2004, 1026, 57–64. [Google Scholar] [CrossRef]
- Buglass, A.J.; Lee, S.H. Sequential analysis of malic acid and both enantiomers of lactic acid in wine using a high-performance liquid chromatographic column-switching procedure. J. Chromatogr. Sci. 2001, 39, 453–458. [Google Scholar] [CrossRef] [Green Version]
- Pallmann, C.L.; Brown, J.A.; Olineka, T.L.; Cocolin, L.; Mills, D.A.; Bisson, L.F. Use of WL medium to profile native flora fermentations. Am. J. Enol. Vitic. 2001, 52, 198–203. [Google Scholar]
- Polizzotto, G.; Barone, E.; Ponticello, G.; Fasciana, T.; Barbera, D.; Corona, O.; Amore, G.; Giammanco, A.; Oliva, D. Isolation, identification and oenological characterization of non-Saccharomyces yeasts in a Mediterranean island. Lett. Appl. Microbiol. 2016, 63, 131–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, P.; Sam, F.E.; Zhang, B.; Peng, S.; Li, M.; Wang, J. Enzymatic characterization of purified β-glucosidase from non-Saccharomyces yeasts and application on chardonnay aging. Foods 2022, 11, 852. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.Z.; Gong, P.F.; Lu, R.R.; Zhang, B.; Morata, A.; Han, S.Y. Effect of different clarification treatments on the volatile composition and aromatic attributes of “Italian riesling” icewine. Molecules 2020, 25, 2657. [Google Scholar] [CrossRef] [PubMed]
- Benito, S. The impact of Torulaspora delbrueckii yeast in winemaking. Appl. Microbiol. Biotechnol. 2018, 102, 3081–3094. [Google Scholar] [CrossRef] [PubMed]
- Binati, R.L.; Lemos Junior, W.J.F.; Luzzini, G.; Slaghenaufi, D.; Ugliano, M.; Torriani, S. Contribution of non-Saccharomyces yeasts to wine volatile and sensory diversity: A study on Lachancea thermotolerans, Metschnikowia spp. and Starmerella bacillaris strains isolated in Italy. Int. J. Food Microbiol. 2020, 318, 108470. [Google Scholar] [CrossRef] [PubMed]
- Câmara, J.S.; Herbert, P.; Marques, J.C.; Alves, M.A. Varietal flavour compounds of four grape varieties producing Madeira wines. Anal. Chim. Acta 2004, 513, 203–207. [Google Scholar] [CrossRef]
- Cai, J.; Zhu, B.Q.; Wang, Y.H.; Lu, L.; Lan, Y.B.; Reeves, M.J.; Duan, C.Q. Influence of pre-fermentation cold maceration treatment on aroma compounds of Cabernet Sauvignon wines fermented in different industrial scale fermenters. Food Chem. 2014, 154, 217–229. [Google Scholar] [CrossRef]
- Zhang, B.Q.; Luan, Y.; Duan, C.Q.; Yan, G.L. Use of Torulaspora delbrueckii co-fermentation with two Saccharomyces cerevisiae strains with different aromatic characteristic to improve the diversity of red wine aroma profile. Front. Microbiol. 2018, 9, 606. [Google Scholar] [CrossRef]
- Lage, P.; Barbosa, C.; Mateus, B.; Vasconcelos, I.; Mendes-Faia, A.; Mendes-Ferreira, A.H. guilliermondii impacts growth kinetics and metabolic activity of S. cerevisiae: The role of initial nitrogen concentration. Int. J. Food Microbiol. 2014, 172, 62–69. [Google Scholar] [CrossRef]
- Simonin, S.; Alexandre, H.; Nikolantonaki, M.; Coelho, C.; Tourdot-Maréchal, R. Inoculation of Torulaspora delbrueckii as a bio-protection agent in winemaking. Food Res. Int. 2018, 107, 451–461. [Google Scholar] [CrossRef]
- Balikci, E.K.; Tanguler, H.; Jolly, N.P.; Erten, H. Influence of Lachancea thermotolerans on cv. Emir wine fermentation. Yeast 2016, 33, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brice, C.; Cubillos, F.A.; Dequin, S.; Camarasa, C.; Martínez, C. Adaptability of the Saccharomyces cerevisiae yeasts to wine fermentation conditions relies on their strong ability to consume nitrogen. PLoS ONE 2018, 13, e0192383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inokuma, K.; Iwamoto, R.; Bamba, T.; Hasunuma, T.; Kondo, A. Improvement of xylose fermentation ability under heat and acid co-stress in Saccharomyces cerevisiae using genome shuffling technique. Front. Bioeng. Biotechol. 2017, 5, 81. [Google Scholar] [CrossRef] [PubMed]
- Augustyn, O.; Pretorius, I.; Jolly, N. The role and use of non-saccharomyces yeasts in wine production. S. Afr. J. Enol. Vitic. 2006, 27, 15–39. [Google Scholar]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef]
- Heit, C.; Martin, S.J.; Yang, F.; Inglis, D.L. Osmoadaptation of wine yeast (Saccharomyces cerevisiae) during icewine fermentation leads to high levels of acetic acid. J. Appl. Microbiol. 2018, 124, 1506–1520. [Google Scholar] [CrossRef]
- Chen, K.; Escott, C.; Loira, I.; del Fresno, J.M.; Morata, A.; Tesfaye, W.; Calderon, F.; Suárez-Lepe, J.A.; Han, S.; Benito, S. Use of non-Saccharomyces yeasts and oenological tannin in red winemaking: Influence on colour, aroma and sensorial properties of young wines. Food Microbiol. 2018, 69, 51–63. [Google Scholar] [CrossRef]
- Kapsopoulou, K.; Kapaklis, A.; Spyropoulos, H. Growth and fermentation characteristics of a strain of the wine yeast Kluyveromyces thermotolerans isolated in Greece. World J. Microbiol. Biotechnol. 2005, 21, 1599–1602. [Google Scholar] [CrossRef]
- Banilas, G.; Sgouros, G.; Nisiotou, A. Development of microsatellite markers for Lachancea thermotolerans typing and population structure of wine-associated isolates. Microbiol. Res. 2016, 193, 1–10. [Google Scholar] [CrossRef]
- Gao, P.; Peng, S.; Sam, F.E.; Zhu, Y.; Liang, L.; Li, M.; Wang, J. Indigenous non-Saccharomyces yeasts with β-glucosidase activity in sequential fermentation with Saccharomyces cerevisiae: A strategy to improve the volatile composition and sensory characteristics of wines. Front. Microbiol. 2022, 13, 845837. [Google Scholar] [CrossRef]
- Du Plessis, H.; Du Toit, M.; Nieuwoudt, H.; Van Der Rijst, M.; Kidd, M.; Jolly, N. Effect of Saccharomyces, non-Saccharomyces yeasts and malolactic fermentation strategies on fermentation kinetics and flavor of Shiraz wines. Fermentation 2017, 3, 64. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.C.; Li, A.H.; Dizy, M.; Ullah, N.; Sun, W.X.; Tao, Y.S. Evaluation of aroma enhancement for “Ecolly” dry white wines by mixed inoculation of selected Rhodotorula mucilaginosa and Saccharomyces cerevisiae. Food Chem. 2017, 228, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Lan, Y.B.; Xiang, X.F.; Qian, X.; Wang, J.M.; Ling, M.Q.; Zhu, B.Q.; Liu, T.; Sun, L.B.; Shi, Y.; Reynolds, A.G.; et al. Characterization and differentiation of key odor-active compounds of ‘Beibinghong’ icewine and dry wine by gas chromatography-olfactometry and aroma reconstitution. Food Chem. 2019, 287, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.C.; Wolff, S.R.; Bisson, L.F.; Ebeler, S.E. Yeast strain and nitrogen supplementation: Dynamics of volatile ester production in chardonnay juice fermentations. Am. J. Enol. Vitic. 2007, 58, 470–483. [Google Scholar]
- Miranda-Lopez, R.; Libbey, L.M.; Watson, B.T.; Mcdaniel, M.R. Odor analysis of pinot noir wines from grapes of different maturities by a gas chromatography-olfactometry technique (Osme). J. Food Sci. 1992, 57, 985–993. [Google Scholar] [CrossRef]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and future of non-Saccharomyces yeasts: From spoilage microorganisms to biotechnological tools for improving wine aroma complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef] [Green Version]
- Andorrà, I.; Berradre, M.; Rozès, N.; Mas, A.; Guillamón, J.M.; Esteve-Zarzoso, B. Effect of pure and mixed cultures of the main wine yeast species on grape must fermentations. Eur. Food Res. Technol. 2010, 231, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Molina, A.M.; Guadalupe, V.; Varela, C.; Swiegers, J.H.; Pretorius, I.S.; Agosin, E. Differential synthesis of fermentative aroma compounds of two related commercial wine yeast strains. Food Chem. 2009, 117, 189–195. [Google Scholar] [CrossRef]
- Saerens, S.M.G.; Delvaux, F.; Verstrepen, K.J.; Van Dijck, P.; Thevelein, J.M.; Delvaux, F.R. Parameters affecting ethyl ester production by Saccharomyces cerevisiae during fermentation. Appl. Environ. Microbiol. 2008, 74, 454–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coetzee, C.; Du Toit, W.J. Sauvignon blanc wine: Contribution of ageing and oxygen on aromatic and non-aromatic compounds and sensory composition: A review. S. Afr. J. Enol. Vitic. 2015, 36, 347–365. [Google Scholar] [CrossRef] [Green Version]
- Whitener, M.E.B.; Stanstrup, J.; Carlin, S.; Divol, B.; Du Toit, M.; Vrhovsek, U. Effect of non-Saccharomyces yeasts on the volatile chemical profile of Shiraz wine. Aust. J. Grape Wine Res. 2017, 23, 179–192. [Google Scholar] [CrossRef]
- Ristic, R.; Bindon, K.; Francis, L.I.; Herderich, M.J.; Iland, P.G. Flavonoids and C13-norisoprenoids in Vitis vinifera L. cv. Shiraz: Relationships between grape and wine composition, wine colour and wine sensory properties. Aust. J. Grape Wine Res. 2016, 16, 369–388. [Google Scholar] [CrossRef]
- Morata, A.; Bañuelos, M.A.; Vaquero, C.; Loira, I.; Cuerda, R.; Palomero, F.; González, C.; Suárez-Lepe, J.A.; Wang, J.; Han, S.; et al. Lachancea thermotolerans as a tool to improve pH in red wines from warm regions. Eur. Food Res. Technol. 2019, 245, 885–894. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Single Fermentation | Sequential Fermentation | |||
|---|---|---|---|---|---|
| SC | LT | TD | L-D | L-S | |
| Residual sugar (g/L) | 159.67 ± 0.21b | 163.67 ± 1.86a | 153.33 ± 1.73d | 158.00 ± 0.08c | 161.67 ± 0.51b |
| Ethanol (% v/v) | 10.33 ± 0.15a | 10.07 ± 0.15a | 10.53 ± 0.45a | 10.77 ± 0.23a | 10.40 ± 0.53a |
| Glycerol (g/L) | 6.40 ± 0.3d | 11.05 ± 0.64c | 11.23 ± 1.89c | 13.00 ± 0.72a | 12.04 ± 0.20b |
| Acetic acid (g/L) | 1.88 ± 0.04a | 0.50 ± 0.08d | 0.84 ± 0.09b | 0.49 ± 0.11d | 0.68 ± 0.11c |
| Total acidity(g/L) | 6.57 ± 0.16e | 8.08 ± 0.41bc | 7.18 ± 0.66d | 9.89 ± 0.38a | 9.29 ± 0.92b |
| pH | 4.03 ± 0.02a | 3.73 ± 0.04c | 3.87 ± 0.12b | 3.67 ± 0.02c | 3.65 ± 0.04c |
| Lactic acid (g/L) | 0.08 ± 0.00d | 2.20 ± 0.04a | 0.67 ± 0.03c | 2.04 ± 0.11b | 2.06 ± 0.05b |
| Malic acid (g/L) | 0.46 ± 0.04d | 1.15 ± 0.087b | 1.41 ± 0.31a | 1.09 ± 0.01c | 1.15 ± 0.08b |
| No. | Compound | Icewines | OTV (μg/L) | ODE | Reference | ||||
|---|---|---|---|---|---|---|---|---|---|
| SC | LT | TD | L-D | L-S | |||||
| Esters | |||||||||
| A1 | Ethyl acetate | 798.96 ± 3.66a | 632.4 ± 12.35c | 585.24 ± 12.53d | 563.34 ± 3.62d | 710.41 ± 4.77b | 7500 | Pineapple, balsam | [25] |
| A2 | Hexyl acetate | 72.59 ± 8.99a | 1.58 ± 0.84d | 5.16 ± 0.56c | 12.72 ± 10.76b | 13.08 ± 5.48b | 1500 | Apple, cherries | [42] |
| A3 | Isoamyl acetate | 111.44 ± 3.56c | 94.23 ± 7.0d | 26.37 ± 2.33e | 156.90 ± 2.72b | 169.54 ± 8.19a | 30 | Sweet, banana | [25] |
| A4 | Heptyl acetate | 3.25 ± 3.25 | ND | ND | ND | ND | 1400 | Rika, Apricot | [25] |
| A5 | Phenethyl acetate | 313.65 ± 13.10b | 16.38 ± 4.94e | 492.66 ± 48.38a | 121.96 ± 1.37c | 31.26 ± 9.30d | 250 | Rose, jasmine | [25] |
| A6 | Ethyl butyrate | 45.32 ± 37.68a | 13.13 ± 3.74c | 6.93 ± 4.30d | 5.65 ± 4.28e | 15.02 ± 7.42b | 20 | Banana, strawberry | [25] |
| A7 | Ethyl hexanoate | 237.83 ± 49.59a | 54.14 ± 6.75c | 97.46 ± 19.10b | 28.76 ± 1.07d | 104.91 ± 0.53b | 5 | Strawberry, apple | [25] |
| A8 | Ethyl heptanoate | 12.65 ± 2.35a | ND | 1.62 ± 0.43b | ND | ND | 220 | Pineapple, fruity | [25] |
| A9 | Ethyl octanoate | 908.02 ± 9.9a | 62.48 ± 4.18e | 193.85 ± 30.56c | 83.77 ± 3.01d | 250.29 ± 8.76b | 240 | Ripe fruits, pear, | [25] |
| A10 | Ethyl Decanoate | 360.3 ± 9.04a | 32.74 ± 18.23d | 59.86 ± 26.00c | 53.03 ± 6.23c | 92.44 ± 30.95b | 200 | Pleasant fruity | [25] |
| A11 | Diethyl succinate | 2.84 ± 1.51a | ND | 1.22 ± 2.18b | 0.76 ± 1.45d | 0.95 ± 0.41c | 200,000 | Fruity, cheese | [25] |
| A12 | Ethyl trans-4-decenoate | 16.09 ± 2.97d | 23.30 ± 15.75c | 196.67 ± 57.50a | 24.06 ± 20.34c | 89.71 ± 1.23b | / | / | |
| A13 | Ethyl Myristate | 9.15 ± 0.94a | 2.38 ± 1.34c | 3.85 ± 1.52b | 4.10 ± 1.31b | 1.99 ± 0.45c | 500 | Mild waxy, soapy | [25] |
| A14 | Ethyl hexadecanoate | 3.57 ± 0.63a | 1.07 ± 0.06b | 0.46 ± 0.92d | 0.74 ± 0.17c | 0.70 ± 0.02c | 1500 | Apple, pineapple | [25] |
| A15 | Ethyl propionate | 12.79 ± 2.93a | ND | 7.02 ± 6.02b | 0.52 ± 0.21c | ND | 10 | Pineapple | [44] |
| A16 | Ethyl dodecanoate | 82.64 ± 0.24a | ND | ND | 32.51 ± 14.4b | 1.76 ± 0.71c | 1500 | Honey, sweety, fruity | [44] |
| A17 | Ethyl lactate | ND | 8.25 ± 0.02b | ND | 13.45 ± 0.15a | 8.23 ± 2.21b | 1500 | Fruity | [42] |
| A18 | 3-Methylbutyl decanoate | 6.21 ± 0.54a | 1.71 ± 0.26b | ND | 0.98 ± 0.21d | 1.28 ± 0.04c | / | / | [44] |
| A19 | Methyl salicylate | 2.39 ± 1.08b | 1.85 ± 0.72c | 3.11 ± 0.67a | 1.82 ± 0.15c | ND | 40 | / | [44] |
| A20 | 2-Phenylethyl propanoate | ND | 1.87 ± 0.72b | 88.55 ± 10.78a | ND | ND | 220 | Pineapple, fruity | |
| Total | 2999.69 | 947.51 | 1770.03 | 1105.07 | 1491.57 | ||||
| Higher alcohols | |||||||||
| B1 | Propanol | 239.9 ± 5.88a | 107.98 ± 3.56d | 103.44 ± 49.83d | 134.02 ± 5.17b | 120.10 ± 6.05c | 50,000 | Fruity | [43] |
| B2 | Isobutanol | 278.2 ± 2.79a | 140.14 ± 7.69b | 111.51 ± 14.95d | 120.18 ± 3.09c | 65.91 ± 8.54e | 75,000 | Fusel oil | [42] |
| B3 | 1-Butanol | 16.50 ± 5.18d | 47.82 ± 15.26c | 21.93 ± 10.27b | 20.64 ± 13.35b | 15.29 ± 5.07c | 150,000 | Medicinal, resinous | [42] |
| B4 | 1-Penten-3-ol | 4.91 ± 0.57a | 2.60 ± 0.41b | 2.62 ± 1.72b | 2.23 ± 1.69c | 2.18 ± 1.44c | / | / | |
| B5 | 1-Pentanol | 3209.74 ± 0.78b | 3773.13 ± 1.30a | 3392.15 ± 2.78b | 9.32 ± 1.09c | 3886.97 ± 5.54a | 8000 | Balsamic, bitter almond | [25] |
| B6 | Isoamyl alcohol | 2091.88 ± 8.83a | ND | ND | ND | 666.80 ± 41.20b | 30,000 | Bitter almond | [43] |
| B7 | 4-Methyl-1-pentanol | ND | 3.52 ± 0.15a | ND | ND | 2.39 ± 0.84b | 50,000 | Almond, toasted | [42] |
| B8 | Heptanol | 37.73 ± 6.85a | 11.76 ± 0.19c | 13.96 ± 0.92b | 8.14 ± 0.86d | 6.77 ± 1.40e | 200 | Grass, oily | [25] |
| B9 | 2-Nonanol | 2.24 ± 0.67a | 1.68 ± 0.03c | ND | 1.59 ± 0.21c | 1.83 ± 0.67b | 58 | Unpleasant floral | [43] |
| B10 | 2,3-Butanediol | 43.85 ± 20.5a | 8.89 ± 2.02c | 8.04 ± 1.93c | 15.56 ± 9.81b | 6.67 ± 4.39c | 120,000 | Aromatic plant | [42] |
| B11 | 1-Octanol | 3.61 ± 0.56c | 3.50 ± 0.81c | 5.33 ± 0.35a | 3.88 ± 1.85b | 3.54 ± 0.35c | 900 | Jasmine, lemon | [25] |
| B12 | 2-Heptanol | ND | 0.43 ± 0.29a | ND | 0.48 ± 0.12a | ND | 200 | Rubbery | [44] |
| B13 | 3-Methyl-1-pentanol | 5.40 ± 1.18b | 10.30 ± 3.71a | 4.74 ± 2.86c | 5.30 ± 3.22b | 4.93 ± 1.71c | 50,000 | Herbaceous, cocoa | [42] |
| B14 | 3-Ethoxypropanol | 10.65 ± 1.65b | 2.70 ± 1.38e | 16.71 ± 1.45a | 7.05 ± 1.43c | 3.98 ± 2.05d | / | Green, citrus | |
| B15 | Cis-2-Hexen-1-ol | 1.08 ± 0.38b | 0.98 ± 0.01b | ND | 0.29 ± 0.34c | 3.80 ± 0.22a | / | / | |
| B16 | 4-Penten-1-ol | 2.71 ± 0.88b | 2.14 ± 0.04c | 1.92 ± 1.32c | 6.11 ± 4.33a | ND | / | Slight fruity aroma | |
| B17 | 1-Decanol | 8.06 ± 2.89a | ND | ND | ND | 0.59 ± 0.54b | 400 | Waxy, fatty | [25] |
| B18 | 1-Hexanol | 339.29 ± 53.56c | 514.16 ± 20.8a | 486.84 ± 76.71b | 363.40 ± 8.51c | 305.94 ± 21.50c | 8000 | Herbaceous, woody | [25] |
| B19 | 3-Hexen-1-ol | 15.51 ± 3.56a | 11.75 ± 3.16b | 10.05 ± 4.05c | ND | 6.58 ± 1.08d | 400 | Orange, fruity | [25] |
| B20 | Benzyl alcohol | 6.00 ± 0.94a | 4.53 ± 0.39c | 4.88 ± 2.36b | 4.72 ± 2.22b | 5.08 ± 3.37b | 620 | Almond, walnut | [44] |
| B21 | Phenethyl alcohol | 1031.40 ± 1.84d | 1304.41 ± 2.20c | 1912.83 ± 1.85a | 1602.6 ± 1.61b | 1177.65 ± 1.08c | 86 | Floral, rose, honey | [25] |
| Total | 7347.04 | 5952.42 | 6096.95 | 2305.51 | 6287 | ||||
| Acids | |||||||||
| C1 | Acetic acid | 681.43 ± 7.87a | 42.07 ± 1.67c | 65.20 ± 2.90b | 24.41 ± 10.72d | 59.81 ± 11.17b | 200000 | Acetic | [42] |
| C2 | Isobutyric acid | 13.28 ± 1.26a | 2.19 ± 0.33e | 6.01 ± 1.02b | 5.37 ± 1.76c | 4.29 ± 1.67d | 8100 | Acetic | [42] |
| C3 | Butyric acid | 9.40 ± 1.73a | 3.79 ± 2.24c | ND | 5.68 ± 1.67b | 4.05 ± 1.22c | 173 | Sour, cheese, fatty | [44] |
| C4 | 2-Methylbutyric acid | 35.64 ± 3.10a | 3.20 ± 0.42e | 5.21 ± 0.40d | 8.04 ± 4.10c | 10.77 ± 6.05b | 50 | Fatty, rancid, cheesy | [44] |
| C5 | Decanoic acid | 134.08 ± 7.77a | 9.41 ± 1.39d | 69.03 ± 0.23c | 68.63 ± 1.67c | 82.61 ± 2.24b | 1400 | Butter, cheese | [25] |
| C6 | Hexanoic acid | 322.52 ± 1.81a | 44.06 ± 4.27d | 77.41 ± 12.72c | 44.36 ± 4.12d | 110.57 ± 4.15b | 420 | Cheese, fatty | [25] |
| C7 | 9-decenoic acid | 231.50 ± 7.4a | ND | 33.44 ± 4.38c | 67.35 ± 7.02b | 63.29 ± 6.00b | / | Acetic acid smell | |
| C8 | Octanoic acid | 659.75 ± 8.49a | 21.03 ± 1.03d | 83.24 ± 38.38c | 33.08 ± 6.61d | 297.78 ± 6.59b | 500 | Rancid, cheese, fatty | [25] |
| C9 | 3-Hydroxylauric acid | 1.43 ± 0.46a | 0.54 ± 0.02c | 0.76 ± 0.98b | 0.50 ± 0.48c | 0.58 ± 0.41c | / | Sour smell, fatty | |
| C10 | Heptanoic acid | 2.21 ± 0.23a | ND | ND | ND | 1.48 ± 0.22b | / | / | |
| C11 | 17-octadecynoic acid | ND | 1.22 ± 0.91a | 1.05 ± 0.72b | 0.82 ± 0.11c | 0.32 ± 0.06d | / | / | |
| C12 | Trans-3-decanoic acid | ND | 4.53 ± 2.02a | ND | ND | ND | / | / | |
| C13 | Pentanoic acid | ND | 20.84 ± 0.72a | ND | ND | ND | / | / | |
| Total | 2091.24 | 152.88 | 341.35 | 258.24 | 635.46 | ||||
| Terpenes | |||||||||
| D1 | Linalool | 12.11 ± 2.7c | 15.57 ± 0.40b | 12.27 ± 1.95c | 18.72 ± 0.03a | 15.58 ± 2.16b | 615 | Floral, musk | [25] |
| D2 | α-Terpineol | ND | 3.50 ± 0.58d | 5.98 ± 0.09c | 7.67 ± 2.87a | 6.74 ± 1.99b | 250 | Floral | [25] |
| D3 | Citronellol | ND | 24.15 ± 3.66d | 28.56 ± 6.29c | 56.53 ± 9.45a | 38.17 ± 8.76b | 30 | Lemon, citrus, rose | [25] |
| D4 | Nerol | ND | 1.71 ± 0.60b | 1.02 ± 2.36c | 2.47 ± 2.36a | 2.69 ± 1.34a | 400 | Rose, lemon | [42] |
| D5 | Geraniol | 9.29 ± 1.98c | 14.22 ± 0.64b | 5.40 ± 0.33d | 9.53 ± 5.40c | 15.51 ± 6.98a | 30 | Peach, floral, musk | [42] |
| D6 | Nerolidol | 1.94 ± 1.94bc | 1.58 ± 0.93d | 2.93 ± 1.95b | 5.71 ± 1.95a | 2.09 ± 0.60c | 700 | Apple, rose | [42] |
| D7 | cis-3-Hexen-1-ol | 15.11 ± 6.56c | 12.79 ± 1.10d | 17.96 ± 7.11b | 18.87 ± 5.26a | 8.48 ± 0.02e | 400 | Plant, fruity, aromatic | [25] |
| D8 | β-damascenone | 2.03 ± 0.12c | 1.45 ± 0.3d | 2.67 ± 0.03b | 2.88 ± 5.90b | 11.15 ± 0.82a | 0.5 | Rose, floral | [25] |
| D9 | Myrcene | 1.18 ± 0.45d | 1.94 ± 1.23b | 0.57 ± 0.09e | 1.38 ± 0.47c | 2.52 ± 0.45a | / | / | |
| Total | 41.66 | 76.91 | 77.36 | 123.76 | 102.93 | ||||
| Others | |||||||||
| E1 | Benzaldehyde | 1.29 ± 0.21c | 3.27 ± 0.26a | 1.28 ± 0.20c | ND | 2.12 ± 0.14b | 0.35 | Bitter almond | [43] |
| E2 | Decanal | 3.94 ± 0.35a | 2.27 ± 2.27b | ND | 2.28 ± 0.59b | ND | 1000 | Fruity, floral | [43] |
| E3 | Methylheptenone | 4.05 ± 14.14a | 0.91 ± 0.27c | ND | 1.22 ± 3.99b | 0.86 ± 0.19c | / | Rose, plum | |
| E4 | Damastone | 17.56 ± 4.43e | 22.38 ± 2.58c | 20.73 ± 2.37d | 34.27 ± 7.99b | 37.59 ± 3.22a | 0.05 | / | [44] |
| E5 | 3-Hydroxy-2-butanone | ND | 4.17 ± 1.41a | ND | 2.75 ± 1.59b | 1.37 ± 0.62c | 150,000 | Peach, plum | [43] |
| E6 | Geranyl acetone | ND | ND | 1.03 ± 0.73b | 2.11 ± 0.89a | 1.10 ± 0.53b | 60 | Floral | [44] |
| E7 | Styrene | 6.06 ± 0.54a | 1.79 ± 0.05d | 2.02 ± 0.77c | 2.91 ± 0.36b | ND | / | Pungent | |
| Total | 32.9 | 34.79 | 25.06 | 45.54 | 43.04 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Ma, Y.; Sam, F.E.; Gao, P.; Liang, L.; Peng, S.; Li, M. The Impact of Indigenous Non-Saccharomyces Yeasts Inoculated Fermentations on ‘Semillon’ Icewine. Fermentation 2022, 8, 413. https://doi.org/10.3390/fermentation8080413
Wang J, Ma Y, Sam FE, Gao P, Liang L, Peng S, Li M. The Impact of Indigenous Non-Saccharomyces Yeasts Inoculated Fermentations on ‘Semillon’ Icewine. Fermentation. 2022; 8(8):413. https://doi.org/10.3390/fermentation8080413
Chicago/Turabian StyleWang, Jing, Yuwen Ma, Faisal Eudes Sam, Pingping Gao, Lihong Liang, Shuai Peng, and Min Li. 2022. "The Impact of Indigenous Non-Saccharomyces Yeasts Inoculated Fermentations on ‘Semillon’ Icewine" Fermentation 8, no. 8: 413. https://doi.org/10.3390/fermentation8080413