
Biodiversity and Oenological Property Analysis of Non-Saccharomyces Yeasts Isolated from Korla Fragrant Pears (Pyrus sinkiangensis Yu)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Spontaneous Fermentation and Sample Collection
2.2. Illumina High-Throughput Sequencing
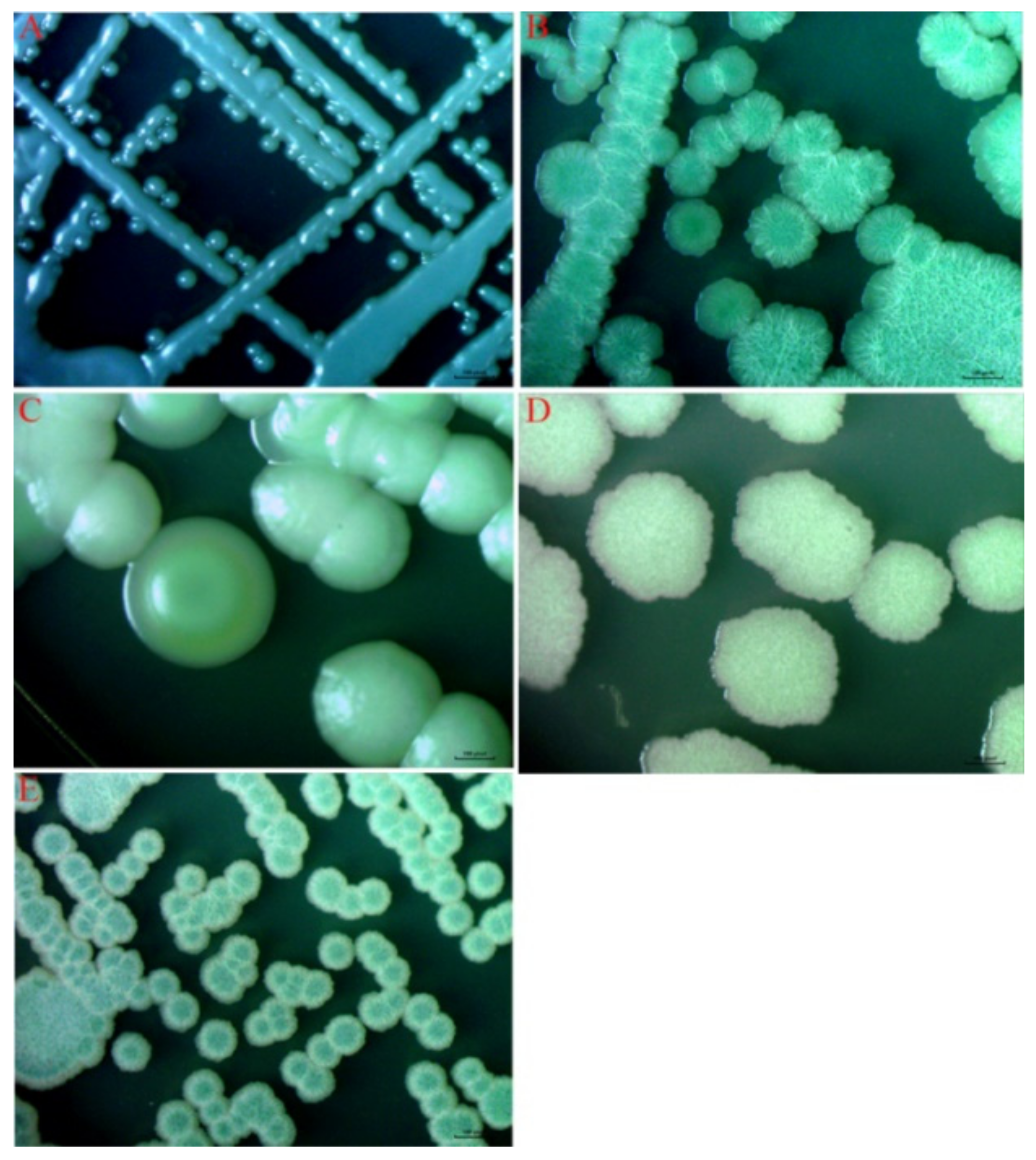
2.3. Culturable Yeasts Isolation and Identification
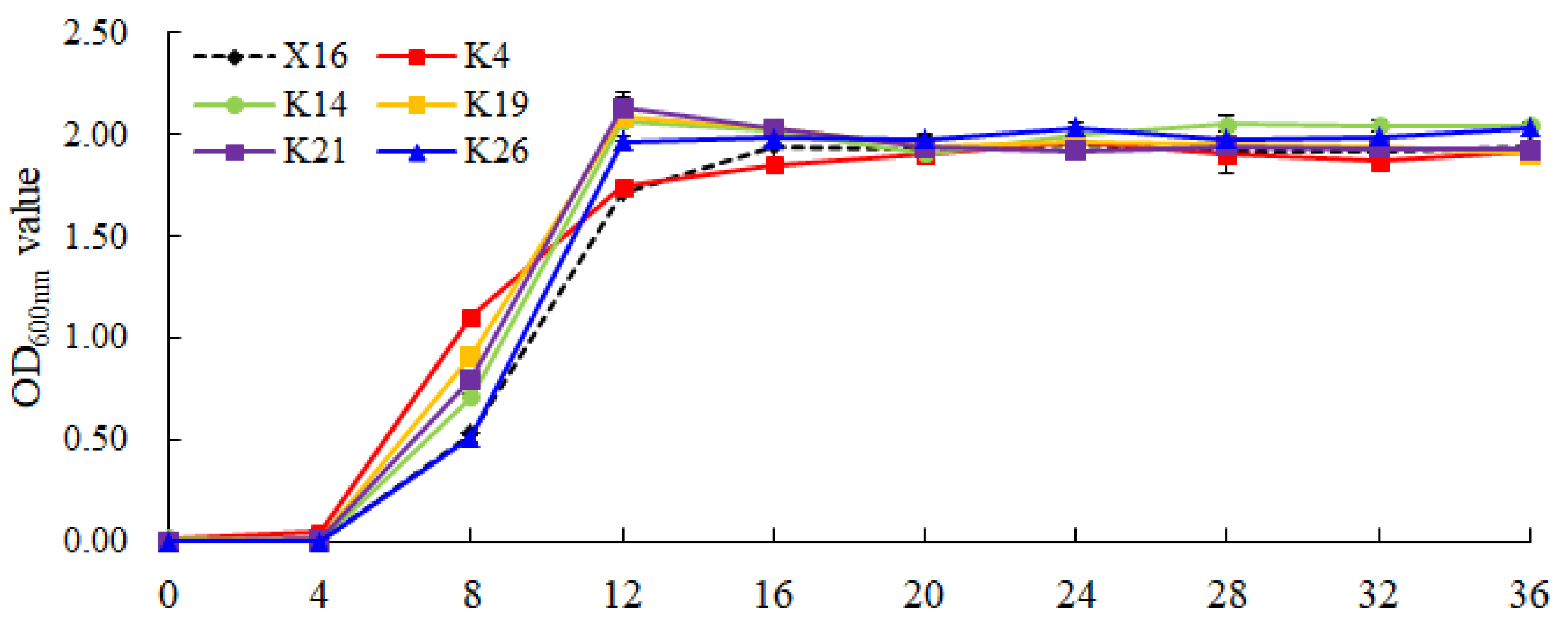
2.4. Growth Curve Determination
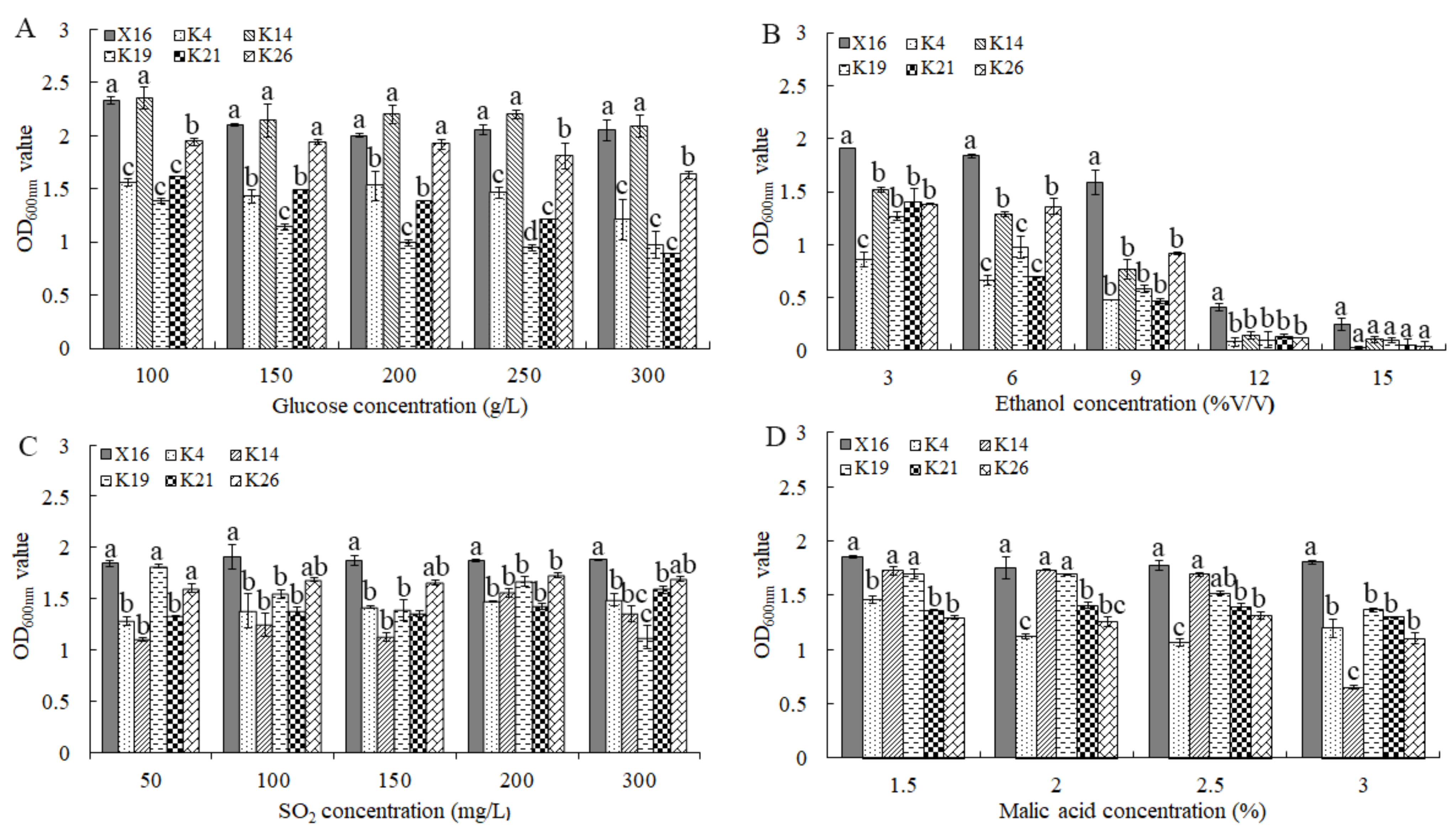
2.5. Oenological Condition Tolerance Analysis
2.6. Determination of the Hydrogen Sulfide (H2S) Production Performance and β-D-Glucosidase Activity of the Selected Yeasts
2.7. Laboratory-Scale Fermentation of KFP Fruit Wine
2.8. Aromatic Composition Analysis of KFP Fruit Wine
2.9. Statistical Analysis
3. Results and Analysis
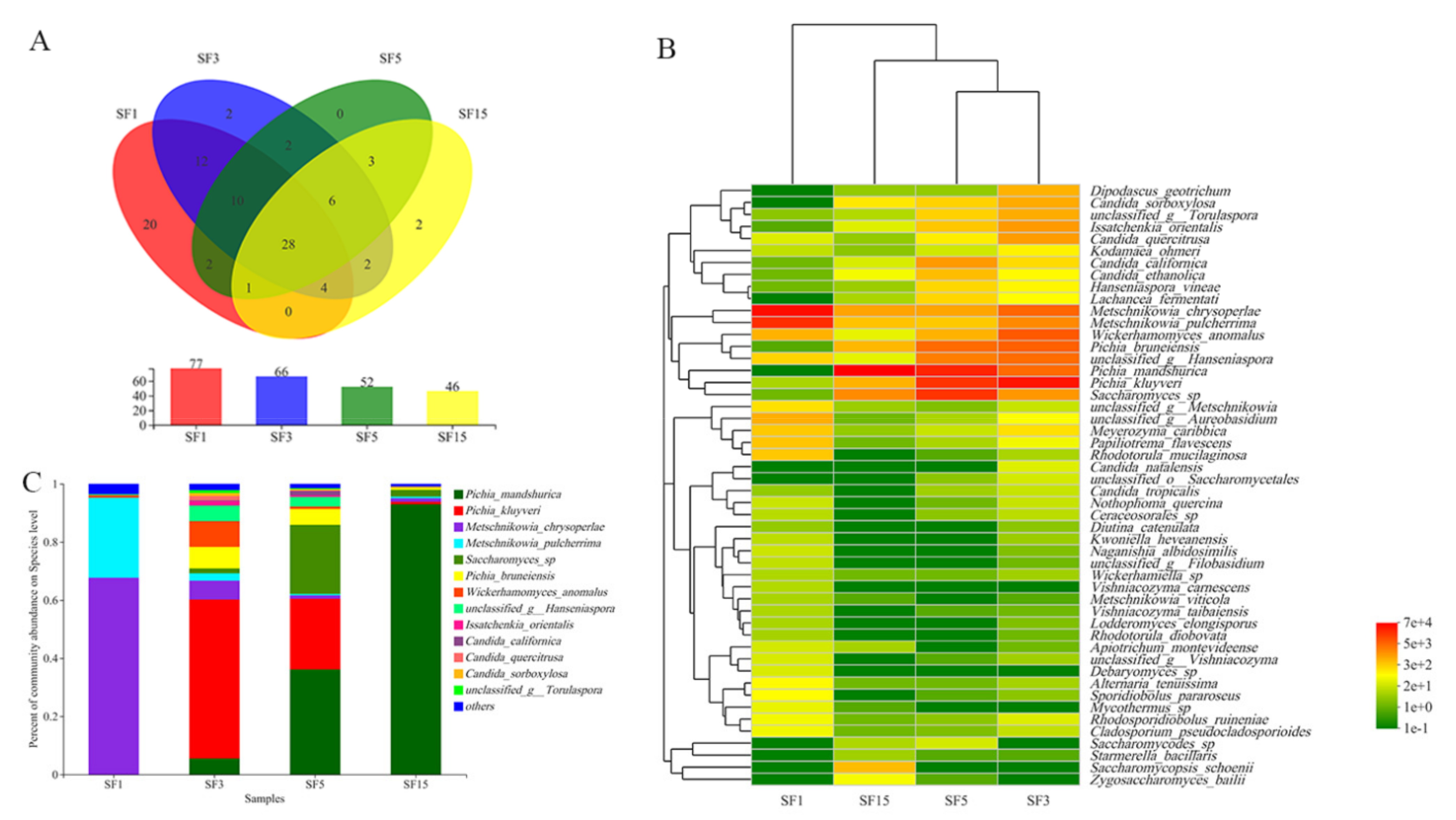
3.1. Non-Saccharomyces Biodiversity Determined Using a High-Throughput Sequencing Approach
3.2. Determination of Diversity of Yeasts Using a Culture-Dependent Approach
3.3. Growth and Carbon Source Utilization Characteristics of the Selected Yeasts
3.4. Properties of Oenological Condition Tolerance of the Selected Yeasts
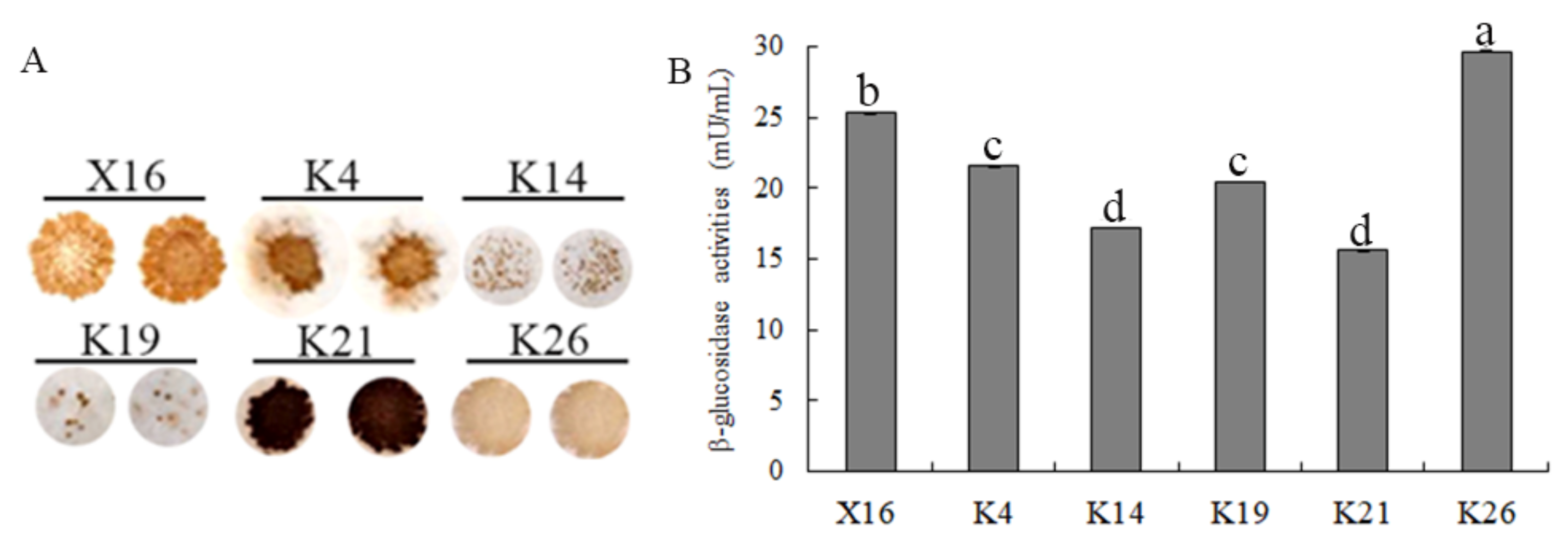
3.5. H2S and β-Glucosidase Production Capacities of the Selected Yeasts
3.6. Winemaking Properties of the Selected Yeasts
3.6.1. Basic Physicochemical Parameters of KFP Wine
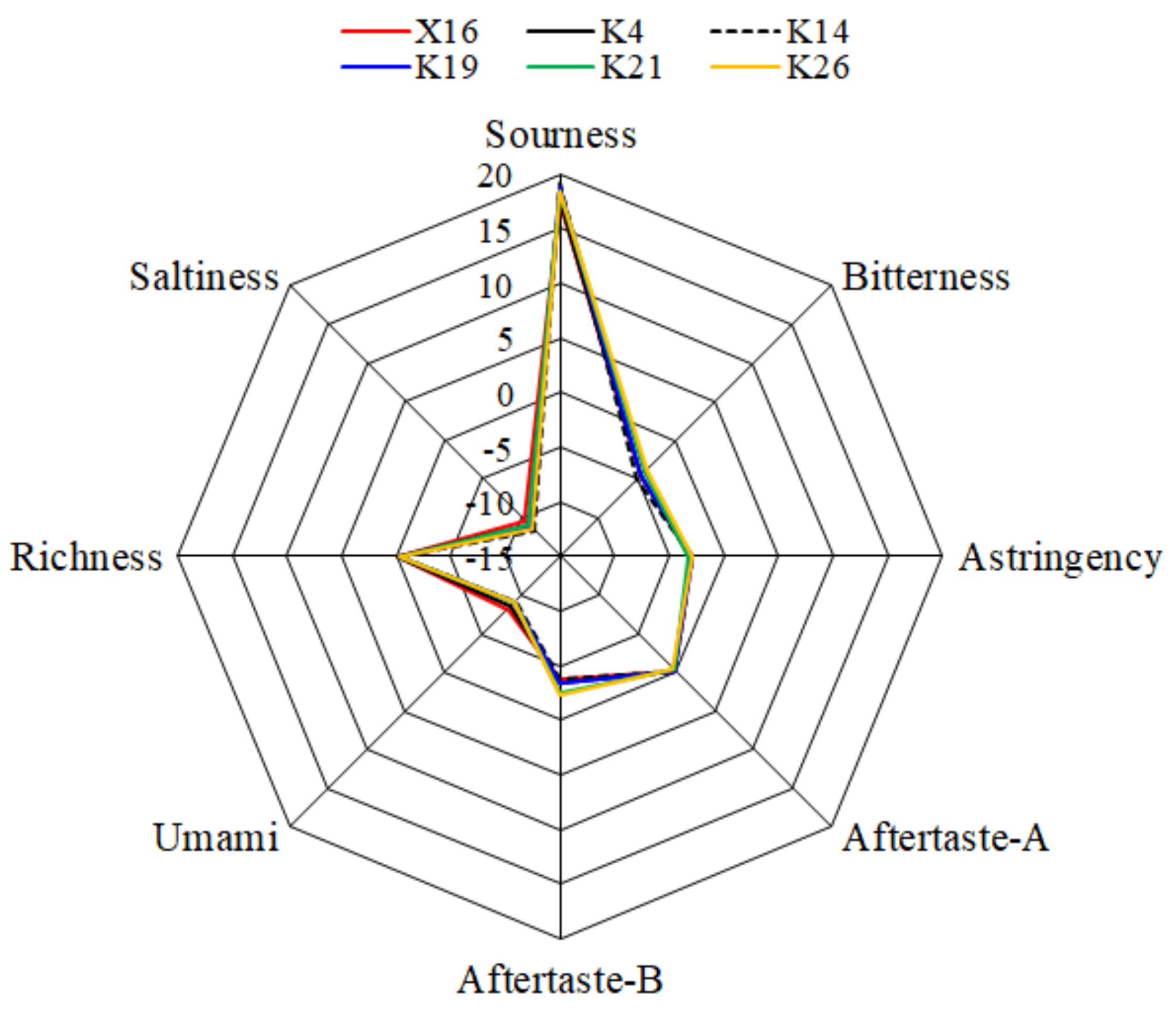
3.6.2. Electronic Sensory Characteristics of KFP Wine
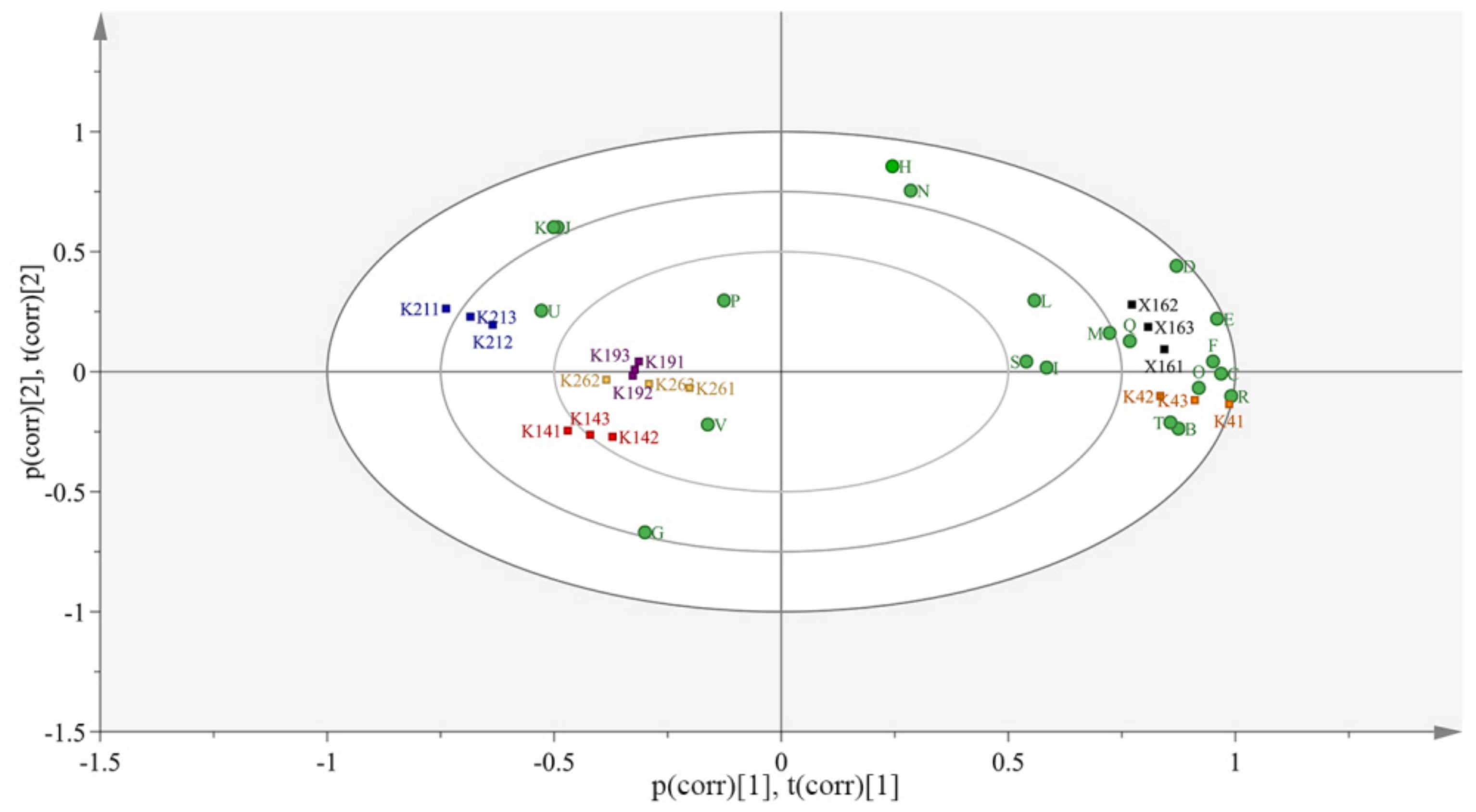
3.6.3. Volatile Aroma Characteristics of KFP Wine
Esters
Alcohols
Acids
Aldoketones
Other Compounds
Aroma Contribution of the Main Compounds in KFP Wine
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, X.Z.; Li, Y.F.; Yu, Z.H.; Hardie, W.J.; Huang, M.Z. Biodiversity of non-Saccharomyces yeasts during natural fermentation of Rosa roxburghii. Acta Microbiol. Sin. 2020, 60, 1696–1708. [Google Scholar]
- Morata, A.; Escott, C.; Bañuelos, M.A.; Loira, I.; del Fresno, J.M.; González, C.; Suárez-Lepe, J.A. Contribution of non-Saccharomyces yeasts to wine freshness: A review. Biomolecules 2020, 10, 34. [Google Scholar] [CrossRef] [PubMed]
- Borren, E.; Tian, B. The important contribution of non-Saccharomyces yeasts to the aroma complexity of wine: A review. Foods 2021, 10, 13. [Google Scholar] [CrossRef] [PubMed]
- Basso, R.F.; Alcarde, A.R.; Portugal, C.B. Could non-Saccharomyces yeasts contribute on innovative brewing fermentations? Food Res. Int. 2016, 86, 112–120. [Google Scholar] [CrossRef]
- Liu, X.Z.; Li, Y.F.; Zhou, J.C.; Huang, M.Z. Effects of co-inoculation and sequential inoculation of Wickerhamomyces anomalus and Saccharomyces cerevisiae on the physicochemical properties and aromatic characteristics of longan (Dimocarpus longan Lour.) wine. Qual. Assur. Saf. Crops 2021, 13, 56–66. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Q.; Niu, H.; Zhang, H.; Lan, H.P.; Zeng, Y.; Jia, F.G. Prediction method for nutritional quality of Korla pear during storage. Int. J. Agr. Biol. Eng. 2021, 14, 247–254. [Google Scholar] [CrossRef]
- Wang, Y.; Su, S.P.; Chen, G.G.; Mao, H.J.; Jiang, Y. Relationship between cuticular waxes and storage quality parameters of Korla pear under different storage methods. J. Plant Growth Regul. 2021, 40, 1152–1165. [Google Scholar] [CrossRef]
- Wang, Y.; Mao, H.; Lv, Y.; Chen, G.G.; Jiang, Y. Comparative analysis of total wax content, chemical composition and crystal morphology of cuticular wax in Korla pear under different relative humidity of storage. Food Chem. 2021, 339, 128097. [Google Scholar] [CrossRef]
- Mamat, A.; Tusong, K.; Xu, J.; Yan, P.; Mei, C.; Wang, J.X. Integrated transcriptomic and proteomic analysis reveals the complex molecular mechanisms underlying stone cell formation in Korla pear. Sci. Rep. 2021, 11, 7688. [Google Scholar] [CrossRef]
- Fu, L.; Zhang, H.; Ye, H. Studies on the yeast fermentation performance of XL1 and XL2 isolated from Korla pear. Food Sci. 2003, 24, 61–64. [Google Scholar]
- Loira, I.; Morata, A.; Comuzzo, P.; Callejo, M.J.; González, C.; Calderón, F.; Suárez-Lepe, J.A. Use of Schizosaccharomyces pombe and Torulaspora delbrueckii strains in mixed and sequential fermentations to improve red wine sensory quality. Food Res. Int. 2015, 76, 325–333. [Google Scholar] [CrossRef]
- Liu, X.Z.; Li, Y.F.; Yu, Z.H.; Hardie, W.J.; Huang, M.Z. Screening and characterisation of β-glucosidase production strains from Rosa roxburghii Tratt. Int. J. Food Eng. 2021, 17, 1–9. [Google Scholar] [CrossRef]
- Liu, X.Z.; Li, Y.F.; Zhao, H.B.; Yu, Z.H.; Huang, M.Z. Oenological property analysis of selected Hanseniaspora uvarum isolated from Rosa roxburghii Tratt. Int. J. Food Eng. 2020, 17, 445–454. [Google Scholar] [CrossRef]
- Song, R.R.; Duan, W.P.; Zhu, X.; Liu, Q.; Yang, X.S.; Han, S.Y. Effects of sequential inoculation with Torulaspora delbrueckii and Saccharomyces cerevisiae on aroma profile of dry red wine. Food Ferment. Ind. 2019, 45, 1–9. [Google Scholar]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and future of non-Saccharomyces yeasts: From spoilage microorganisms to biotechnological tools for improving wine aroma complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef]
- Tufariello, M.; Fragasso, M.; Pico, J.; Panighel, A.; Castellarin, S.D.; Flamini, R.; Grieco, F. Influence of non-Saccharomyces on wine chemistry: A focus on aroma-related compounds. Molecules 2021, 26, 644. [Google Scholar] [CrossRef]
- Hu, J.; Vinothkanna, A.; Wu, M.; Ekumah, J.N.; Akpabli-Tsigbe, N.D.K.; Ma, Y.K. Tracking the dynamic changes of a flavor, phenolic profile, and antioxidant properties of Lactiplantibacillus plantarum-and Saccharomyces cerevisiae-fermented mulberry wine. Food Sci. Nutr. 2021, 9, 6294–6306. [Google Scholar] [CrossRef]
- Kwon, S.J.; Choi, J.S.; Yeo, S.H.; Par, H.D. Fermentation characteristics of persimmon wine by the mixed culture of Pichia anomala JK04 and Saccharomyces cerevisiae Fermivin cells. Korean J. Food Preserv. 2015, 22, 768–777. [Google Scholar] [CrossRef]
- Chen, D.; Yap, Z.Y.; Liu, S.Q. Evaluation of the performance of Torulaspora delbrueckii, Williopsis saturnus, and Kluyveromyces lactis in lychee wine fermentation. Int. J. Food Microbiol. 2015, 206, 45–50. [Google Scholar] [CrossRef]
- Xu, Y.; Zhao, G.A.; Wang, L.P. Controlled formation of volatile components in cider making using a combination of Saccharomyces cerevisiae and Hanseniaspora valbyensis yeast species. J. Ind. Microbiol. Biot. 2006, 33, 192–196. [Google Scholar] [CrossRef]
- Soon, W.W.; Hariharan, M.; Snyder, M.P. Highthroughput sequencing for biology and medicine. Mol. Syst. Biol. 2013, 9, 640. [Google Scholar] [CrossRef]
- Zhao, L.Y.; Li, Y.; Jiang, L.W.; Deng, F.M. Determination of fungal community diversity in fresh and traditional Chinese fermented pepper by pyrosequencing. FEMS Microbiol. Lett. 2016, 363, fnw273. [Google Scholar] [CrossRef]
- Wei, Y.J.; Wu, Y.; Yan, Y.Z.; Zou, W.; Xue, J.; Ma, W.R.; Wang, W.; Tian, G.; Wang, L.Y. Highthroughput sequencing of microbial community diversity in soil, grapes, leaves, grape juice and wine of grapevine from China. PLoS ONE 2018, 13, e0193097. [Google Scholar]
- Auesukaree, C. Molecular mechanisms of the yeast adaptive response and tolerance to stresses encountered during ethanol fermentation. J. Biosci. Bioeng. 2017, 124, 133–142. [Google Scholar] [CrossRef]
- Hu, K.; Qin, Y.; Tao, Y.S.; Zhu, X.L.; Peng, G.T.; Ullah, N. Potential of glycosidase from non-Saccharomyces isolates for enhancement of wine aroma. J. Food Sci. 2016, 81, M935–M943. [Google Scholar] [CrossRef]
- Margolles, A.; de los Reyes-Gavilan, C.G. Purification and functional characterization of a novel α-l-arabinofuranosidase from Bifidobacterium longum B667. Appl. Environ. Microb. 2003, 69, 5096–5103. [Google Scholar] [CrossRef]
- Gallego, M.V.; Pinaga, F.; Ramón, D.; Vallés, S. Purification and characterization of an α-L-Rhamnosidase from Aspergillus terreus of interest in winemaking. J. Food Sci. 2001, 66, 204–209. [Google Scholar] [CrossRef]
- Padilla, B.; Gil, J.V.; Manzanares, P. Challenges of the non-conventional yeast Wickerhamomyces anomalus in winemaking. Fermentation 2018, 4, 68. [Google Scholar] [CrossRef]
- Balmaseda, A.; Bordons, A.; Reguant, C.; Bautista-Gallego, J. Non-Saccharomyces in wine: Effect upon Oenococcus oeni and malolactic fermentation. Front. Microbiol. 2018, 9, 534. [Google Scholar] [CrossRef]
- Varela, C. The impact of non-Saccharomyces yeasts in the production of alcoholic beverages. Appl. Microbiol. Biotechnol. 2016, 100, 9861–9874. [Google Scholar] [CrossRef]
- Saerens, S.; Thevelein, J.; Delvaux, F. Ethyl ester production during brewery fermentation, a review. Cerevisia 2008, 33, 82–90. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Colony Colour | Colony Topography |
|---|---|---|
| A | Steel gray | Knob-like, convex, smooth |
| B | White on the surface and cyan in the center | Convex, wrinkled surface, irregular edge |
| C | Slightly yellow to cream | Convex, wrinkled, and opaque, irregularKnob-like, convex, smooth |
| D | white | Convex, wrinkled surface, irregular edge, opaque surface |
| E | White on the surface and cyan in the center | Convex, wrinkled surface, irregular edge, opaque surface |
| Strains | Glucose | Fructose | Sucrose | Maltose | Galactose | Mannose | Arabinose |
|---|---|---|---|---|---|---|---|
| X16 | + | + | + | + | + | + | − |
| K4 | + | + | − | + | + | + | − |
| K14 | + | + | + | + | − | − | − |
| K19 | + | + | + | − | + | + | − |
| K21 | + | − | − | + | + | + | − |
| K26 | + | + | + | + | + | + | − |
| Strains | Alcohol Content (% v/v) | Residual Sugar (g/L) | pH | Total Acidity (g/L) | Volatile Acidity (g/L) |
|---|---|---|---|---|---|
| X16 | 13.66 ± 0.19 a | 8.42 ± 0.30 a | 3.84 ± 0.02 a | 4.42 ± 0.10 a | 0.34 ± 0.02 b |
| K4 | 12.63 ± 0.18 c | 6.88 ± 0.30 c | 3.89 ± 0.02 a | 3.60 ± 0.10 b | 0.78 ± 0.10 a |
| K14 | 13.09 ± 0.15 b | 7.40 ± 0.40 bc | 3.81 ± 0.04 a | 4.34 ± 0.10 a | 0.64 ± 0.03 a |
| K19 | 13.95 ± 0.15 a | 8.02 ± 0.20 ab | 3.84 ± 0.02 a | 4.34 ± 0.10 a | 0.28 ± 0.03 b |
| K21 | 12.75 ± 0.12 bc | 7.58 ± 0.30 bc | 3.84 ± 0.05 a | 3.96 ± 0.02 b | 0.77 ± 0.02 a |
| K26 | 12.91 ± 0.21 bc | 7.19 ± 0.60 bc | 3.89 ± 0.01 a | 3.84 ± 0.10 b | 0.79 ± 0.10 a |
| No. | Compounds | CAS | Groups | |||||
|---|---|---|---|---|---|---|---|---|
| X16 | K4 | K14 | K19 | K21 | K26 | |||
| 1 | Ethyl acetate | 141-78-6 | 17.82 ± 0.89 b | 18.8 ± 1.79 b | 16.27 ± 1.87 b | 27.47 ± 2.64 a | 17.00 ± 4.16 b | 27.41 ± 0.18 a |
| 2 | Ethyl butyrate | 105-54-4 | 0.96 ± 0.08 a | 1.00 ± 0.00 a | 0.53 ± 0.05 b | ND | ND | 0.52 ± 0.00 b |
| 3 | Ethyl octanoate | 106-32-1 | 47.74 ± 1.29 b | 62.29 ± 0.88 a | 17.90 ± 2.19 c | 20.62 ± 0.14 c | 16.92 ± 3.28 c | 20.37 ± 0.58 c |
| 4 | Ethyl caprate | 110-38-3 | 93.12 ± 13.25 a | 67.21 ± 0.73 b | 15.11 ± 0.35 e | 44.02 ± 4.81 c | 30.52 ± 1.35 d | 32.80 ± 1.95 d |
| 5 | Ethyl laurate | 106-33-2 | 1.67 ± 0.00 c | 12.63 ± 0.62 a | 0.37 ± 0.01 d | 2.39 ± 0.19 b | 0.20 ± 0.00 e | 0.33 ± 0.00 d |
| 6 | Ethyl hexanoate | 123-66-0 | 20.91 ± 9.21 a | 19.12 ± 0.07 a | 5.24 ± 0.13 bc | 4.72 ± 0.19 c | 3.16 ± 0.23 d | 6.04 ± 0.68 b |
| 7 | Ethyl isovalerate | 108-64-5 | 0.18 ± 0.00 | ND | ND | ND | ND | ND |
| 8 | Ethyl pelargonate | 123-29-5 | 1.06 ± 1.49 abc | 1.49 ± 0.08 b | 2.74 ± 0.54 a | 1.13 ± 0.04 c | 1.46 ± 0.08 b | 1.27 ± 0.00 c |
| 9 | Ethyl propionate | 105-37-3 | ND | ND | 0.19 ± 0.00 | ND | ND | ND |
| 10 | Hexyl acetate | 142-92-7 | 1.25 ± 0.24 a | 0.27 ± 0.09 bc | 0.11 ± 0.01 c | 0.41 ± 0.02 b | 0.94 ± 0.28 a | ND |
| 11 | Isobutyl acetate | 110-19-0 | 0.07 ± 0.00 b | 0.34 ± 0.08 a | ND | ND | 0.13 ± 0.05 b | 0.02 ± 0.00 b |
| 12 | Isoamyl acetate | 123-92-2 | ND | ND | ND | ND | 4.80 ± 0.00 | ND |
| 13 | Methyl acetate | 79-20-9 | 0.23 ± 0.01 a | 0.07 ± 0.00 b | ND | ND | ND | 0.03 ± 0.00 b |
| 14 | Phenethyl acetate | 103-45-7 | ND | ND | ND | 1.06 ± 0.68 b | 29.66 ± 1.46 a | ND |
| 15 | Methyl caprylate | 111-11-5 | 0.28 ± 0.10 | ND | ND | ND | ND | ND |
| 16 | Vinyl formate | 692-45-5 | 0.64 ± 0.03 b | 0.69 ± 0.04 b | 1.15 ± 0.33 a | 0.28 ± 0.12 c | 0.26 ± 0.00 c | 0.61 ± 0.00 b |
| ΣEsters | 185.93 ± 26.59 a | 183.91 ± 4.38 a | 59.61 ± 5.48 c | 102.10 ± 8.83 b | 105.05 ± 10.89 b | 89.40 ± 3.39 b | ||
| 17 | 1-Butanol | 71-36-3 | 0.01 ± 0.00 a | 0.06 ± 0.06 a | 0.06 ± 0.03 a | ND | ND | 0.04 ± 0.00 a |
| 18 | (R,R)-2,3-Butanediol | 24347-58-8 | 13.89 ± 0.00 a | 3.57 ± 0.50 c | 3.71 ± 0.72 c | 3.39 ± 0.01 c | 1.68 ± 0.17 | 7.33 ± 0.04 b |
| 19 | cis-4-Decen-1-ol | 57074-37-0 | 4.86 ± 1.73 ab | 5.51 ± 0.46 a | 3.79 ± 1.03 b | 5.85 ± 0.38 a | 1.36 ± 0.36 c | 6.01 ± 0.84 a |
| 20 | 1-Hexanol | 111-27-3 | 6.22 ± 1.19 a | 5.37 ± 0.02 a | 3.72 ± 0.42 b | 0.66 ± 0.12 d | 3.54 ± 0.20 b | 2.46 ± 0.19 c |
| 21 | (Z)-3-Hexen-1-ol | 928-96-1 | ND | ND | 0.23 ± 0.00 | ND | ND | ND |
| 22 | Isoamylol | 123-51-3 | 18.11 ± 2.34 ab | 15.46 ± 2.13 b | 2.57 ± 0.46 d | 5.64 ± 0.46 c | 23.37 ± 3.48 a | 5.46 ± 0.42 c |
| 23 | Isobutanol | 78-83-1 | 16.08 ± 4.05 a | 20.41 ± 1.44 a | 6.74 ± 0.70 cd | 8.56 ± 1.02 c | 6.70 ± 0.07 d | 12.26 ± 1.49 b |
| 24 | Methyl alcohol | 67-56-1 | 2.07 ± 0.56 a | 1.21 ± 0.40 a | 1.01 ± 0.56 a | 0.04 ± 0.00 c | 0.28 ± 0.33 bc | 0.38 ± 0.27 b |
| 25 | 1-Nonanol | 143-08-8 | 2.06 ± 0.31 b | 0.43 ± 0.10 d | 0.23 ± 0.00 e | 3.01 ± 0.17 a | 0.91 ± 0.24 c | 3.15 ± 0.00 a |
| 26 | 3,6-Nonadien-1-ol, (E,Z)- | 56805-23-3 | 4.02 ± 0.03 a | 2.74 ± 0.00 c | 2.31 ± 0.05 d | 3.55 ± 0.31 b | 3.36 ± 0.00 b | 2.38 ± 0.53 cd |
| 27 | 1-Octanol | 111-87-5 | 5.23 ± 0.86 a | 5.31 ± 0.87 a | 0.17 ± 0.09 d | 4.42 ± 0.74 ab | 0.34 ± 0.03 c | 3.63 ± 0.45 b |
| 28 | 1-Pentanol | 71-41-0 | 174.59 ± 9.53 a | 193.72 ± 11.75 a | 93.97 ± 6.80 b | 89.05 ± 1.23 b | 63.57 ± 7.07 c | 102.83 ± 13.46 b |
| 29 | Phenethyl alcohol | 60-12-8 | 18.56 ± 3.05 a | 22.14 ± 1.97 a | 11.01 ± 0.58 b | 24.60 ± 4.05 a | 10.97 ± 1.24 b | 12.64 ± 1.28 b |
| 30 | 1-Propanol | 71-23-8 | 1.09 ± 0.52 abc | 1.36 ± 0.31 a | 0.58 ± 0.01 c | 0.79 ± 0.00 b | 0.17 ± 0.09 d | 0.81 ± 0.23 b |
| ∑Alcohols | 266.79 ± 24.17 a | 277.29 ± 20.01 a | 130.10 ± 11.45 b | 149.56 ± 8.49 b | 116.25 ± 13.28 c | 159.38 ± 19.20 b | ||
| 31 | Acetic acid | 64-19-7 | 6.91 ± 2.11 c | 2.79 ± 0.26 d | 6.28 ± 1.54 c | 7.48 ± 0.23 c | 9.49 ± 0.16 b | 14.24 ± 2.46 a |
| 32 | Gamma-Linolenic acid | 506-26-3 | ND | ND | ND | 2.32 ± 0.00 | ND | ND |
| 33 | Hexanoic acid | 142-62-1 | ND | ND | 0.37 ± 0.00 | ND | ND | ND |
| 34 | Octanoic acid | 124-07-2 | 3.97 ± 0.79 a | 0.14 ± 0.02 c | 4.14 ± 1.39 a | 3.64 ± 0.12 a | 0.60 ± 0.11 b | 3.39 ± 0.10 a |
| ∑Acids | 10.88 ± 2.90 bc | 2.93 ± 0.26 d | 10.79 ± 2.93 bc | 13.44 ± 0.35 b | 10.09 ± 0.27 c | 17.63 ± 2.56 a | ||
| 35 | Glycolaldehyde | 141-46-8 | ND | 0.06 ± 0.00 | ND | ND | ND | ND |
| 36 | 4-Hydroxy-2-butanone | 590-90-9 | ND | ND | ND | 0.03 ± 0.00 | ND | ND |
| ∑Aldoketones | ND | 0.06 ± 0.00 a | ND | 0.03 ± 0.00 a | ND | ND | ||
| 37 | Carbamic acid, monoammonium salt | 1111-78-0 | ND | 3.79 ± 0.89 a | 1.18 ± 0.00 d | 1.60 ± 0.00 c | 1.45 ± 0.12 c | 2.34 ± 0.38 b |
| 38 | 2,4-Di-tert-butylphenol | 96-76-4 | 17.01 ± 0.13 c | 9.96 ± 1.42 d | 16.87 ± 0.51 c | 34.75 ± 4.92 a | 35.84 ± 3.88 a | 19.45 ± 0.34 b |
| 39 | 3,3-dimethyl-1,5-dioxaspiro [5.5]undecane | 707-29-9 | ND | ND | ND | ND | 0.30 ± 0.00 b | 0.49 ± 0.00 a |
| 40 | Ethanol, 2-[2-(ethenyloxy)ethoxy]- | 929-37-3 | ND | ND | 1.99 ± 0.00 | ND | ND | ND |
| 41 | Ethylbenzene | 100-41-4 | ND | 1.52 ± 0.04 a | 0.33 ± 0.00 c | 0.27 ± 0.00 d | 0.31 ± 0.04 cd | 0.97 ± 0.00 b |
| 42 | 2-methyl-1,5-dioxaspiro [5.5]undecane | 6413-26-9 | 1.71 ± 0.29 f | 8.92 ± 0.04 e | 23.73 ± 1.18 a | 11.87 ± 0.90 d | 14.21 ± 0.35 c | 19.52 ± 0.47 b |
| 43 | 2-Phenyl-1-propene | 98-83-9 | 0.64 ± 0.03 a | 0.61 ± 0.10 a | 0.25 ± 0.00 c | ND | 0.36 ± 0.09 c | 0.56 ± 0.02 b |
| 44 | Propane, 1-ethoxy- | 628-32-0 | ND | ND | ND | 1.00 ± 0.02 | ND | ND |
| 45 | 3-Tridecene | 61883-33-8 | ND | ND | ND | ND | 1.58 ± 0.00 | ND |
| ∑Other compounds | 19.36 ±0.45 d | 24.80 ± 2.49 c | 44.35 ± 1.69 b | 49.49 ± 5.84 ab | 54.05 ± 4.48 a | 43.33 ± 1.21 b | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Huang, Y.; Long, H.; Yu, Z.; Huang, M.; Liu, X. Biodiversity and Oenological Property Analysis of Non-Saccharomyces Yeasts Isolated from Korla Fragrant Pears (Pyrus sinkiangensis Yu). Fermentation 2022, 8, 388. https://doi.org/10.3390/fermentation8080388
Li Y, Huang Y, Long H, Yu Z, Huang M, Liu X. Biodiversity and Oenological Property Analysis of Non-Saccharomyces Yeasts Isolated from Korla Fragrant Pears (Pyrus sinkiangensis Yu). Fermentation. 2022; 8(8):388. https://doi.org/10.3390/fermentation8080388
Chicago/Turabian StyleLi, Yinfeng, Yuanmin Huang, Hua Long, Zhihai Yu, Mingzheng Huang, and Xiaozhu Liu. 2022. "Biodiversity and Oenological Property Analysis of Non-Saccharomyces Yeasts Isolated from Korla Fragrant Pears (Pyrus sinkiangensis Yu)" Fermentation 8, no. 8: 388. https://doi.org/10.3390/fermentation8080388