
Application of Cool Fermentation Temperatures to Encourage Non-Saccharomyces Yeasts to Yield Lower Ethanol Concentrations in Wines

Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Starter Cultures
2.2. Fermentations
2.3. Analytical Analyses
2.4. Sensory Analyses
2.5. Statistical Analyses
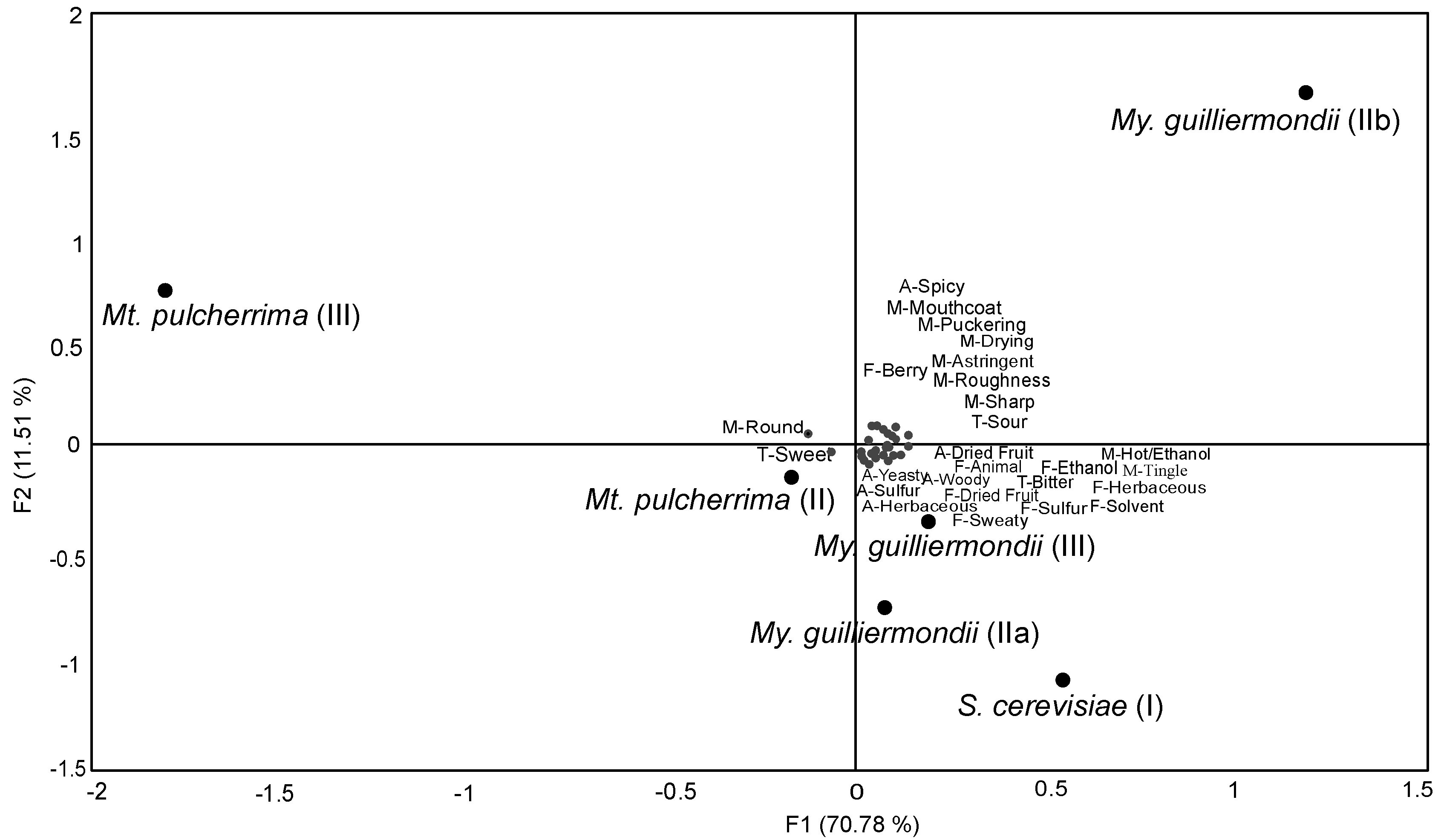
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gonzalez, R.; Quirós, M.; Morales, P. Yeast respiration of sugars by non-Saccharomyces yeast species: A promising and barely explored approach to lowering alcohol content of wines. Trends Food Sci. Technol. 2013, 29, 55–61. [Google Scholar] [CrossRef]
- Cioch-Skoneczny, M.; Satora, P.; Skoneczny, S.; Pater, A. Determination of the oenological properties of yeast strains isolated from spontaneously fermented grape musts obtained from cool climate grape varieties. Eur. Food Res. Technol. 2020, 246, 2299–2307. [Google Scholar] [CrossRef]
- Gobbi, M.; De Vero, L.; Solieri, L.; Comitini, F.; Oro, L.; Giudici, P.; Ciani, M. Fermentative aptitude of non-Saccharomyces wine yeast for reduction in the ethanol content in wine. Eur. Food Res. Technol. 2014, 239, 41–48. [Google Scholar] [CrossRef]
- Morales, P.; Rojas, V.; Quirós, M.; Gonzalez, R. The impact of oxygen on the final alcohol content of wine fermented by a mixed starter culture. Appl. Microbiol. Biotechnol. 2015, 99, 3993–4003. [Google Scholar] [CrossRef]
- Contreras, A.; Hidalgo, C.; Henschke, P.A.; Chambers, P.J.; Curtin, C.; Varela, C. Evaluation of non-Saccharomyces yeasts for the reduction of alcohol content in wine. Appl. Environ. Microbiol. 2014, 80, 1670–1678. [Google Scholar] [CrossRef]
- Contreras, A.; Curtin, C.; Varela, C. Yeast population dynamics reveal a potential ‘collaboration’ between Metschnikowia pulcherrima and Saccharomyces uvarum for the production of reduced alcohol wines during Shiraz fermentation. Appl. Microbiol. Biotechnol. 2015, 99, 1885–1895. [Google Scholar] [CrossRef]
- Contreras, A.; Hidalgo, C.; Schmidt, S.; Henschke, P.A.; Curtin, C.; Varela, C. The application of non-Saccharomyces yeast in fermentations with limited aeration as a strategy for the production of wine with reduced alcohol content. Int. J. Food Microbiol. 2015, 205, 7–15. [Google Scholar] [CrossRef]
- Englezos, V.; Rantsiou, K.; Cravero, F.; Torchio, F.; Ortiz-Julien, A.; Gerbi, V.; Rolle, L.; Cocolin, L. Starmerella bacillaris and Saccharomyces cerevisiae mixed fermentations to reduce ethanol content in wine. Appl. Microbiol. Biotechnol. 2016, 100, 5515–5526. [Google Scholar] [CrossRef]
- Rodrigues, A.J.; Raimbourg, T.; Gonzalez, R.; Morales, P. Environmental factors influencing the efficacy of different yeast strains for alcohol level reduction in wine by respiration. LWT Food Sci. Technol. 2016, 65, 1038–1043. [Google Scholar] [CrossRef]
- García, M.; Esteve-Zarzoso, B.; Cabellos, J.M.; Arroyo, T. Sequential non-Saccharomyces and Saccharomyces cerevisiae fermentations to reduce the alcohol content in wine. Fermentation 2020, 6, 60. [Google Scholar] [CrossRef]
- Hranilovic, A.; Gambetta, J.M.; Jeffery, D.W.; Grbin, P.R.; Jiranek, V. Lower-alcohol wines produced by Metschnikowia pulcherrima and Saccharomyces cerevisiae co-fermentations: The effect of sequential inoculation timing. Int. J. Food Microbiol. 2020, 329, 108651. [Google Scholar] [CrossRef] [PubMed]
- Aplin, J.J.; Edwards, C.G. Impacts of non-Saccharomyces species and aeration on sequential inoculation with Saccharomyces cerevisiae to produce lower alcohol Merlot wines. J. Sci. Food Agric. 2020, 101, 1715–1719. [Google Scholar] [CrossRef]
- Aplin, J.J.; Paup, V.D.; Ross, C.F.; Edwards, C.G. Chemical and sensory profiles of Merlot wines produced by sequential inoculation of Metschnikowia pulcherrima or Meyerozyma guilliermondii. Fermentation 2021, 7, 126. [Google Scholar] [CrossRef]
- Casalta, E.; Aguera, E.; Picou, C.; Rodriguez-Bencomo, J.J.; Salmon, J.M.; Sablayrolles, J.M. A comparison of laboratory and pilot-scale fermentations in winemaking conditions. Appl. Microbiol. Biotechnol. 2010, 87, 1665–1673. [Google Scholar] [CrossRef] [PubMed]
- Viana, T.; Loureiro-Dias, M.C.; Prista, C. Efficient fermentation of an improved synthetic grape must by enological and laboratory strains of Saccharomyces cerevisiae. AMB Express 2014, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- Aplin, J.J.; White, K.P.; Edwards, C.G. Growth and metabolism of non-Saccharomyces yeasts isolated from Washington state vineyards in media and high sugar grape musts. Food Microbiol. 2019, 77, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Rossouw, D.; Jolly, N.; Jacobson, D.; Bauer, F.F. The effect of scale on gene expression: Commercial versus laboratory wine fermentations. Appl. Microbiol. Biotechnol. 2012, 93, 1207–1219. [Google Scholar] [CrossRef]
- Canonico, L.; Comitini, F.; Ciani, M. Metschnikowia pulcherrima selected strain for ethanol reduction in wine: Influence of cell immobilization and aeration condition. Foods 2019, 8, 378. [Google Scholar] [CrossRef]
- Schmid, F.; Schadt, J.; Jiranek, V.; Block, D.E. Formation of temperature gradients in large- and small-scale red wine fermentations during cap management. Aust. J. Grape Wine Res. 2009, 15, 249–255. [Google Scholar] [CrossRef]
- Gao, C.; Fleet, G.H. The effects of temperature and pH on the ethanol tolerance of the wine yeasts, Saccharomyces cerevisiae, Candida stellata and Kloeckera apiculata. J. Appl. Bacteriol. 1988, 65, 405–409. [Google Scholar] [CrossRef]
- Erten, H. Relations between elevated temperatures and fermentation behaviour of Kloeckera apiculata and Saccharomyces cerevisiae associated with winemaking in mixed cultures. World J. Microbiol. Biotechnol. 2002, 18, 377–382. [Google Scholar] [CrossRef]
- Zohre, D.E.; Erten, H. The influence of Kloeckera apiculata and Candida pulcherrima yeasts on wine fermentation. Process Biochem. 2002, 38, 319–324. [Google Scholar] [CrossRef]
- Zott, K.; Miot-Sertier, C.; Claisse, O.; Lonvaud-Funel, A.; Masneuf-Pomarede, I. Dynamics and diversity of non-Saccharomyces yeasts during the early stages in winemaking. Int. J. Food Microbiol. 2008, 125, 197–203. [Google Scholar] [CrossRef]
- Andorrà, I.; Landi, S.; Mas, A.; Esteve-Zarzoso, B.; Guillamón, J.M. Effect of fermentation temperature on microbial population evolution using culture-independent and dependent techniques. Food Res. Int. 2010, 43, 773–779. [Google Scholar] [CrossRef]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef]
- Tanguler, H. Influence of temperatures and termentation behaviour of mixed cultures of Williopsis saturnus var. saturnus and Saccharomyces cerevisiae associated with winemaking. Food Sci. Technol. Res. 2013, 19, 781–793. [Google Scholar] [CrossRef]
- Maturano, Y.P.; Mestre, M.V.; Kuchen, B.; Toro, M.E.; Mercado, L.A.; Vazquez, F.; Combina, M. Optimization of fermentation-relevant factors: A strategy to reduce ethanol in red wine by sequential culture of native yeasts. Int. J. Food Microbiol. 2019, 289, 40–48. [Google Scholar] [CrossRef]
- Bourret, T.B.; Grove, G.G.; Vandemark, G.J.; Henick-Kling, T.; Glawe, D.A. Diversity and molecular determination of wild yeasts in a central Washington State vineyard. N. Am. Fungi 2013, 8, 1–32. [Google Scholar] [CrossRef]
- Eyéghé-Bickong, H.A.; Alexandersson, E.O.; Gouws, L.M.; Young, P.R.; Vivier, M.A. Optimisation of an HPLC method for the simultaneous quantification of the major sugars and organic acids in grapevine berries. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2012, 885–886, 43–49. [Google Scholar] [CrossRef]
- Heard, G.M.; Fleet, G.H. The effects of temperature and pH on the growth of yeast species during the fermentation of grape juice. J. Appl. Bacteriol. 1988, 65, 23–28. [Google Scholar] [CrossRef]
- Ciani, M.; Comitini, F.; Mannazzu, I.; Domizio, P. Controlled mixed culture fermentation: A new perspective on the use of non-Saccharomyces yeasts in winemaking. FEMS Yeast Res. 2010, 10, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Abeln, F.; Chuck, C.J. The role of temperature, pH and nutrition in process development of the unique oleaginous yeast Metschnikowia pulcherrima. J. Chem. Technol. Biotechnol. 2020, 95, 1163–1172. [Google Scholar] [CrossRef]
- Beltran, G.; Novo, M.; Guillamón, J.M.; Mas, A.; Rozès, N. Effect of fermentation temperature and culture media on the yeast lipid composition and wine volatile compounds. Int. J. Food Microbiol. 2008, 121, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Fleet, G.H.; Heard, G.M. Yeast: Growth During Fermentation. In Wine Microbiology and Biotechnology; Fleet, G.H., Ed.; Harwood Academic Publishers: Chur, Switzerland, 1993; pp. 27–54. [Google Scholar]
- Varela, C.; Sengler, F.; Solomon, M.; Curtin, C. Volatile flavour profile of reduced alcohol wines fermented with the non-conventional yeast species Metschnikowia pulcherrima and Saccharomyces uvarum. Food Chem. 2016, 209, 57–64. [Google Scholar] [CrossRef]
- Varela, C.; Barker, A.; Tran, T.; Borneman, A.; Curtin, C. Sensory profile and volatile aroma composition of reduced alcohol Merlot wines fermented with Metschnikowia pulcherrima and Saccharomyces uvarum. Int. J. Food Microbiol. 2017, 252, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Quirós, M.; Rojas, V.; Gonzalez, R.; Morales, P. Selection of non-Saccharomyces yeast strains for reducing alcohol levels in wine by sugar respiration. Int. J. Food Microbiol. 2014, 181, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Medina, K.; Boido, E.; Dellacassa, E.; Carrau, F. Growth of non-Saccharomyces yeasts affects nutrient availability for Saccharomyces cerevisiae during wine fermentation. Int. J. Food Microbiol. 2012, 157, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Oro, L.; Ciani, M.; Comitini, F. Antimicrobial activity of Metschnikowia pulcherrima on wine yeasts. J. Appl. Microbiol. 2014, 116, 1209–1217. [Google Scholar] [CrossRef] [PubMed]
- Ciani, M.; Beco, L.; Comitini, F. Fermentation behaviour and metabolic interactions of multistarter wine yeast fermentations. Int. J. Food Microbiol. 2006, 108, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.K.; Castura, J.C.; Ross, C.F. Temporal check-all-that-apply characterization of Syrah wine. J. Food Sci. 2016, 81, 1521–1529. [Google Scholar] [CrossRef]
- Villamor, R.R.; Ross, C.F. Wine matrix compounds affect perception of wine aromas. Ann. Rev. Food Sci. Technol. 2013, 4, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Seguinot, P.; Ortiz-Julien, A.; Camarasa, C. Impact of nutrient availability on the fermentation and production of aroma compounds under sequential inoculation with M. pulcherrima and S. cerevisiae. Front. Microbiol. 2020, 11, 305. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Pickering, G.J. Ethanol difference thresholds in wine and the influence of mode of evaluation and wine style. Am. J. Enol. Vitic. 2008, 59, 146–152. [Google Scholar]
- Chambers, D.H.; Allison, A.-M.A.; Chambers, E. Training effects on performance of descriptive panelists. J. Sens. Stud. 2004, 19, 486–499. [Google Scholar] [CrossRef]
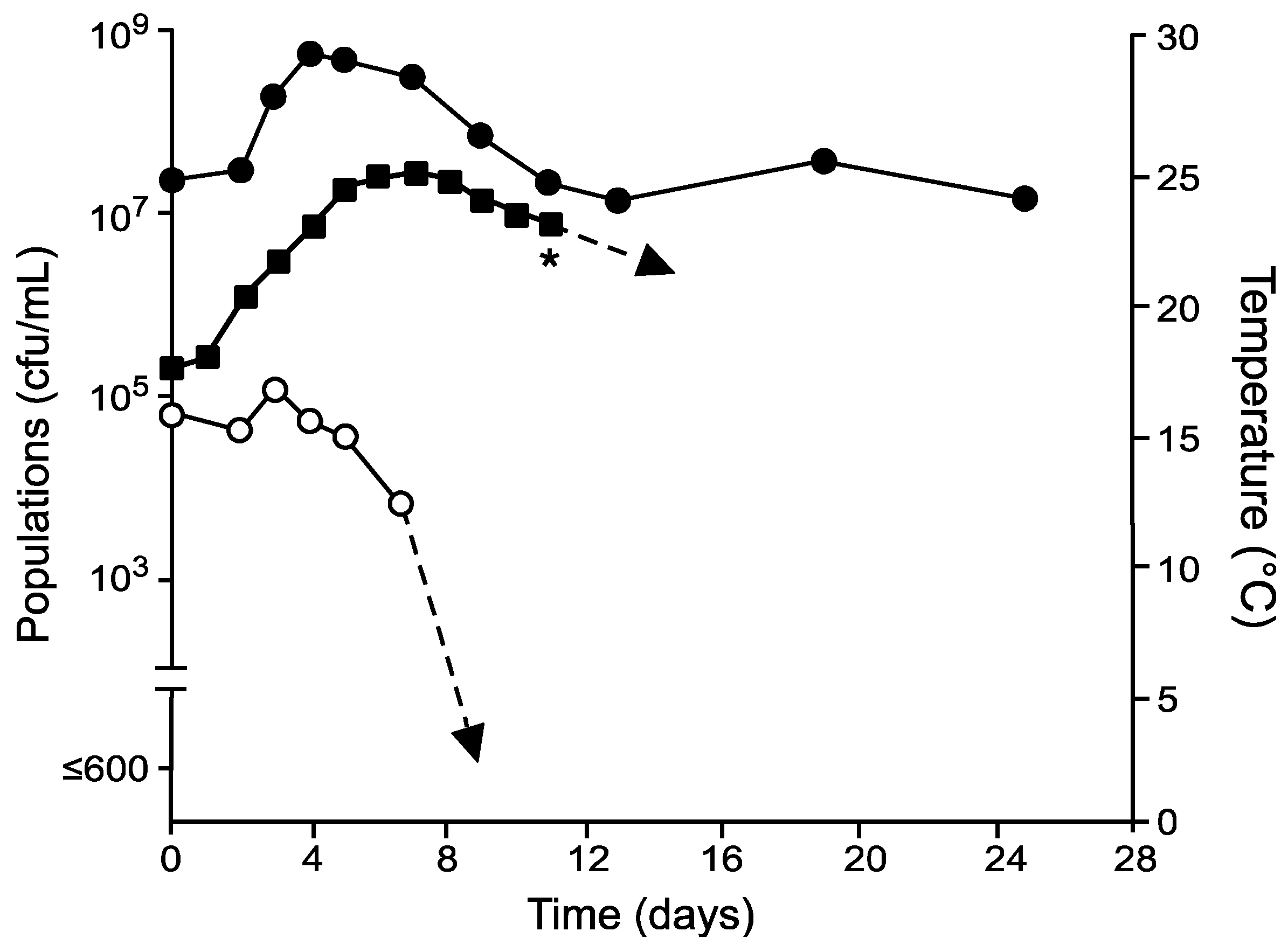
 ), total non-Saccharomyces yeasts (
), total non-Saccharomyces yeasts (  ), and temperatures (
), and temperatures (  ) in grape musts inoculated on day 0 with S. cerevisiae following fermentation temperature regime I. * Denotes time when the wine was pressed (approximately 0° Brix) prior to storage at 21 °C.
), total non-Saccharomyces yeasts ( ), and temperatures ( ) in grape musts inoculated on day 0 with S. cerevisiae following fermentation temperature regime I. * Denotes time when the wine was pressed (approximately 0° Brix) prior to storage at 21 °C.
) in grape musts inoculated on day 0 with S. cerevisiae following fermentation temperature regime I. * Denotes time when the wine was pressed (approximately 0° Brix) prior to storage at 21 °C.
), total non-Saccharomyces yeasts ( ), and temperatures ( ) in grape musts inoculated on day 0 with S. cerevisiae following fermentation temperature regime I. * Denotes time when the wine was pressed (approximately 0° Brix) prior to storage at 21 °C.
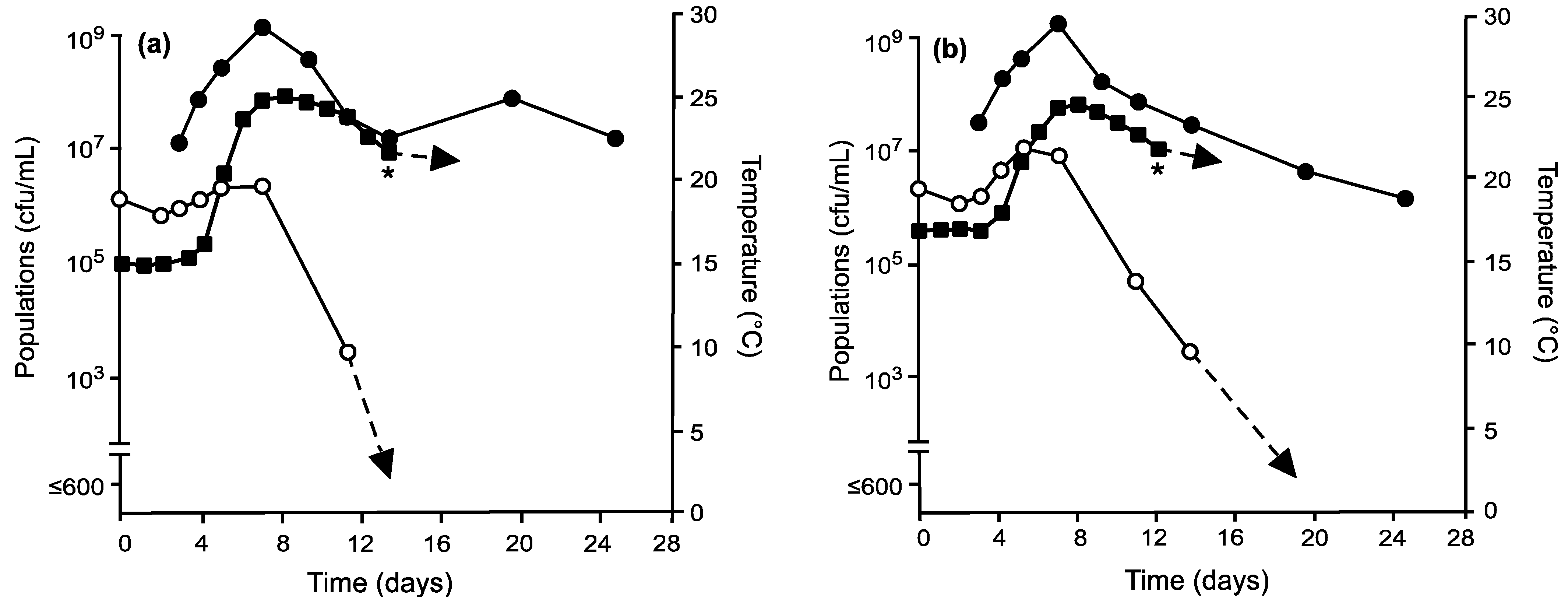
 ), total non-Saccharomyces yeasts (
), total non-Saccharomyces yeasts (  ), and fermentation temperatures (
), and fermentation temperatures (  ) in grape musts inoculated on day 0 with Mt. pulcherrima prior to inoculation with S. cerevisiae on day 3 and following fermentation temperature regime II (a) or III (b). * Denotes time when the wine was pressed (approximately 0° Brix) prior to storage at 21 °C.
), total non-Saccharomyces yeasts ( ), and fermentation temperatures ( ) in grape musts inoculated on day 0 with Mt. pulcherrima prior to inoculation with S. cerevisiae on day 3 and following fermentation temperature regime II (a) or III (b). * Denotes time when the wine was pressed (approximately 0° Brix) prior to storage at 21 °C.
) in grape musts inoculated on day 0 with Mt. pulcherrima prior to inoculation with S. cerevisiae on day 3 and following fermentation temperature regime II (a) or III (b). * Denotes time when the wine was pressed (approximately 0° Brix) prior to storage at 21 °C.
), total non-Saccharomyces yeasts ( ), and fermentation temperatures ( ) in grape musts inoculated on day 0 with Mt. pulcherrima prior to inoculation with S. cerevisiae on day 3 and following fermentation temperature regime II (a) or III (b). * Denotes time when the wine was pressed (approximately 0° Brix) prior to storage at 21 °C.
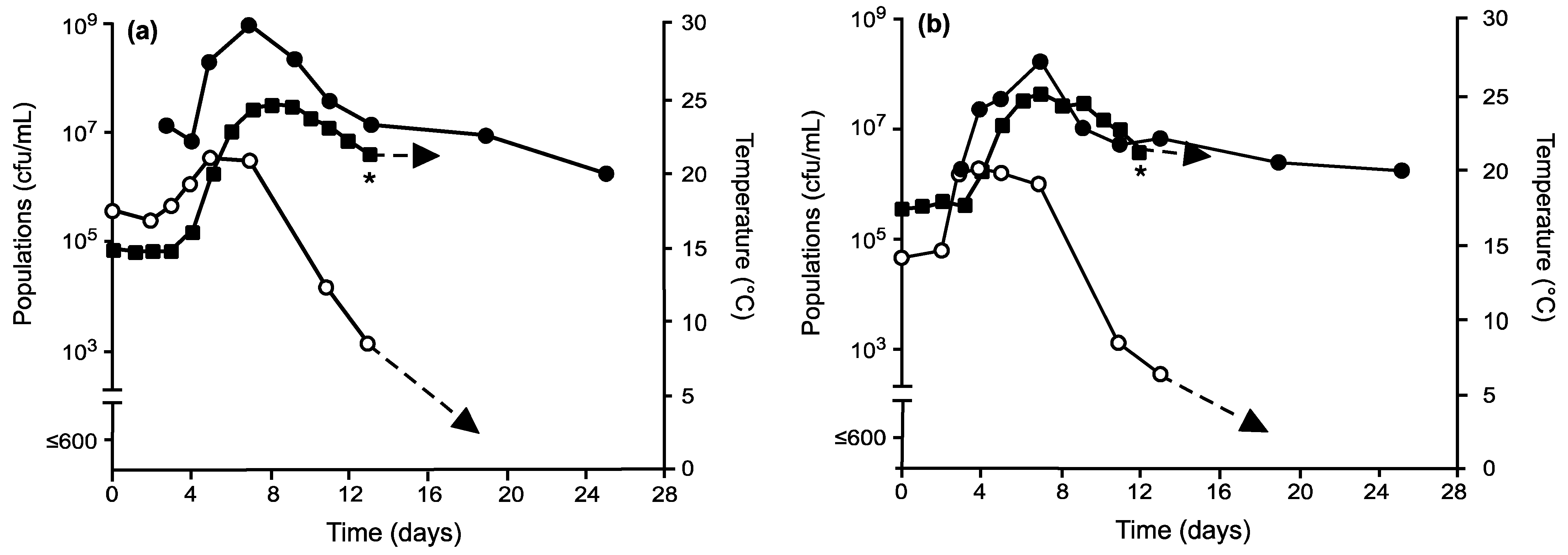
 ), total non-Saccharomyces
yeasts (
), total non-Saccharomyces
yeasts (  ), and temperatures
(
), and temperatures
(  ) in grape musts
inoculated on day 0 with My. guilliermondii prior to inoculation with S.
cerevisiae on day 3 and following fermentation temperature regime II (a)
or III (b). * Denotes time when the wine was pressed (approximately 0°
Brix) prior to storage at 21 °C.
), total non-Saccharomyces
yeasts ( ), and temperatures
( ) in grape musts
inoculated on day 0 with My. guilliermondii prior to inoculation with S.
cerevisiae on day 3 and following fermentation temperature regime II (a)
or III (b). * Denotes time when the wine was pressed (approximately 0°
Brix) prior to storage at 21 °C.
) in grape musts
inoculated on day 0 with My. guilliermondii prior to inoculation with S.
cerevisiae on day 3 and following fermentation temperature regime II (a)
or III (b). * Denotes time when the wine was pressed (approximately 0°
Brix) prior to storage at 21 °C.
), total non-Saccharomyces
yeasts ( ), and temperatures
( ) in grape musts
inoculated on day 0 with My. guilliermondii prior to inoculation with S.
cerevisiae on day 3 and following fermentation temperature regime II (a)
or III (b). * Denotes time when the wine was pressed (approximately 0°
Brix) prior to storage at 21 °C.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yeast Strain (Temperature Regime) | Glu + Fru (g/L) | Ethanol (% v/v) | Glycerol (g/L) | Succinic Acid (g/L) | Acetic Acid (g/L) |
|---|---|---|---|---|---|
| S. cerevisiae (I) | nd | 15.0 a | 9.96 a | 2.19 a | 0.704 a |
| Mt. pulcherrima (II) | nd | 14.8 ab | 9.41 bc | 1.89 c | 0.569 ab |
| Mt. pulcherrima (III) | nd | 14.2 c | 9.27 c | 1.99 b | 0.526 b |
| My. guilliermondii (II) | nd | 14.7 b | 9.34 bc | 2.01 b | 0.486 b |
| My. guilliermondii (III) | nd | 14.6 b | 9.52 b | 2.02 b | 0.483 b |
| Source of Variation (Degrees Freedom) | ||||
|---|---|---|---|---|
| Compound | Yeast (2) | Temperature (1) | Replicate (1) | Yeast * Temperature (1) |
| Ethanol | 10.6 *** | 19.1 *** | 0.01 | 12.0 ** |
| Glycerol | 31.3 *** | 0.05 | 0.00 | 5.78 * |
| Succinic acid | 21.1 *** | 3.48 | 8.51 ** | 1.72 |
| Acetic acid | 4.50 * | 0.16 | 0.07 | 1.25 |
| Treatment | ||||||
|---|---|---|---|---|---|---|
| Attribute | S. cerevisiae (I) | My. guilliermondii (IIa) * | My. guilliermondii (IIb) * | My. guilliermondii (III) | Mt. pulcherrima (II) | Mt. pulcherrima (III) |
| Aroma | ||||||
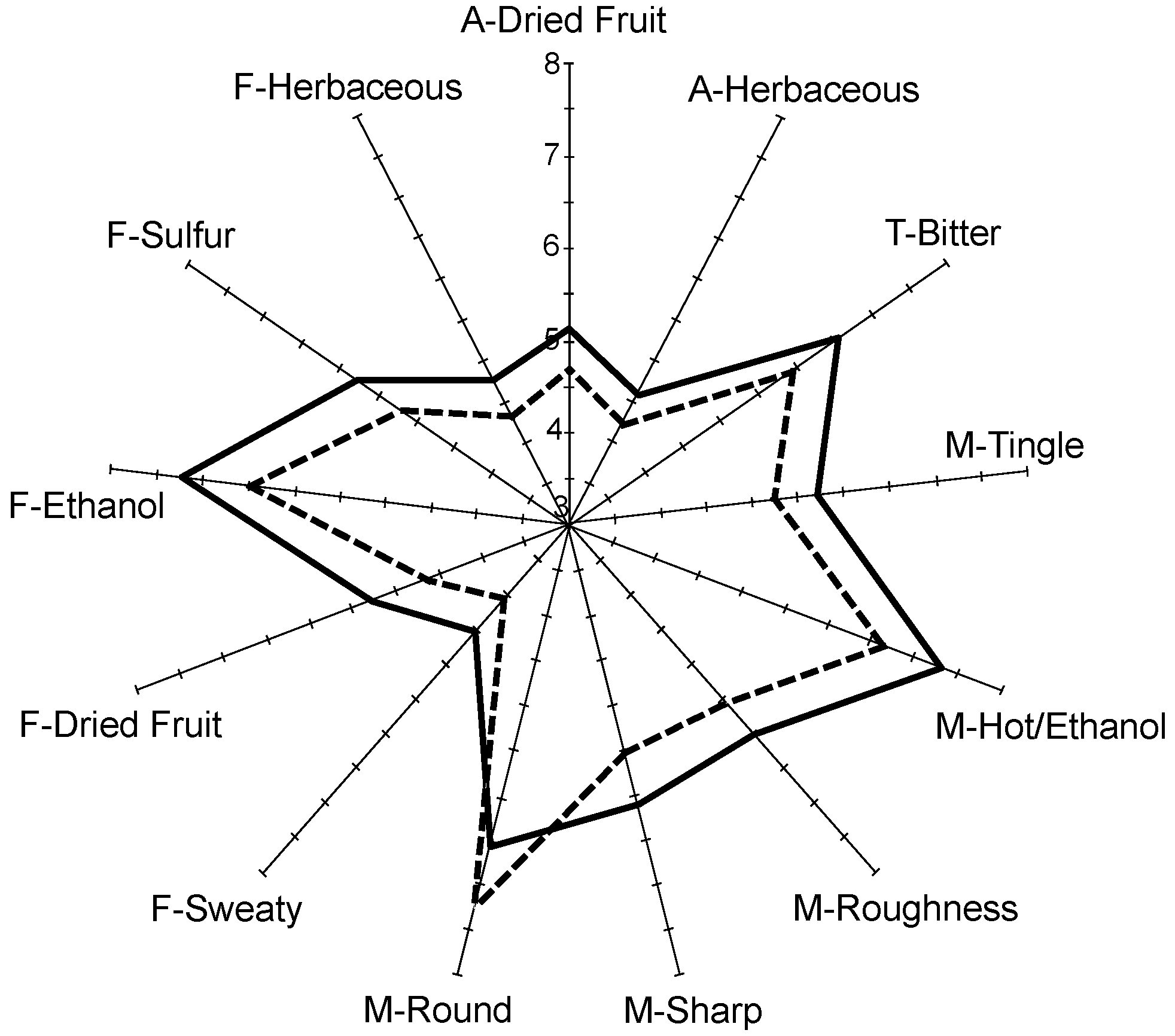
| ‘Fruity’ | 5.6 a | 5.7 a | 5.7 a | 5.7 a | 5.8 a | 5.9 a |
| ‘Floral’ | 5.2 a | 5.5 a | 5.5 a | 5.6 a | 5.4 a | 5.6 a |
| ‘Berry’ | 5.4 a | 5.5 a | 5.4 a | 5.5 a | 5.3 a | 5.6 a |
| ‘Chocolate’ | 3.8 a | 3.9 a | 4.0 a | 3.9 a | 3.8 a | 3.9 a |
| ‘Sweaty’ | 4.5 a | 4.4 a | 4.4 a | 4.2 a | 4.4 a | 4.1 a |
| ‘Dried Fruit’ | 5.1 a | 4.9 ab | 5.1 ab | 5.1 a | 5.1 a | 4.7 b |
| ‘Ethanol’ | 6.9 a | 6.7 a | 6.9 a | 6.9 a | 6.7 a | 6.6 a |
| ‘Sulfur’ | 5.6 ab | 5.8 a | 5.4 a | 5.5 ab | 5.6 ab | 5.3 b |
| ‘Solvent’ | 5.7 a | 5.7 a | 5.8 a | 5.7 a | 5.6 a | 5.5 a |
| ‘Buttery’ | 4.1 a | 4.0 a | 4.2 a | 4.1 a | 4.2 a | 4.0 a |
| ‘Woody’ | 4.7 ab | 4.4 ab | 4.5 ab | 4.5 ab | 4.7 a | 4.3 b |
| ‘Animal’ | 4.4 a | 4.4 a | 4.5 a | 4.3 a | 4.3 a | 4.2 a |
| ‘Herbaceous’ | 4.6 a | 4.4 ab | 4.3 ab | 4.4 ab | 4.5 ab | 4.2 b |
| ‘Spicy’ | 3.9 b | 3.7 b | 4.2 a | 3.8 b | 3.8 b | 3.7 b |
| ‘Yeasty’ | 4.6 ab | 4.5 ab | 4.5 ab | 4.5 ab | 4.8 a | 4.3 b |
| Taste | ||||||
| ‘Sweet’ | 6.2 ab | 6.0 abc | 5.9 bc | 6.0 abc | 5.8 c | 6.3 a |
| ‘Bitter’ | 6.6 a | 6.5 a | 6.5 a | 6.5 a | 6.2 ab | 5.9 b |
| ‘Sour’ | 6.8 abc | 6.9 ab | 7.1 a | 7.0 ab | 6.7 bc | 6.4 c |
| Flavor | ||||||
| ‘Fruity’ | 5.7 a | 5.6 a | 5.5 a | 5.7 a | 5.5 a | 5.6 a |
| ‘Floral’ | 5.3 a | 5.5 a | 5.5 a | 5.5 a | 5.3 a | 5.3 a |
| ‘Berry’ | 5.2 ab | 5.1b | 5.4 ab | 5.5 a | 5.2 ab | 5.1 b |
| ‘Chocolate’ | 3.7 a | 3.6 a | 3.8 a | 3.8 a | 3.7 a | 3.7 a |
| ‘Sweaty’ | 4.5 a | 4.4 ab | 4.3 ab | 4.3 ab | 4.2 ab | 4.1 b |
| ‘Dried Fruit’ | 5.3 a | 4.8 bc | 5.9 abc | 5.1 ab | 5.0 abc | 4.7 c |
| ‘Ethanol’ | 7.2 a | 7.1 a | 7.2 a | 7.0 a | 7.2 a | 6.5 b |
| ‘Sulfur’ | 5.8 a | 5.8 a | 5.7 a | 5.7 a | 5.6 ab | 5.2 b |
| ‘Solvent’ | 6.0 ab | 6.1 a | 6.1 a | 6.1 a | 5.9 ab | 5.6 b |
| ‘Buttery’ | 4.1 a | 4.0 a | 4.1 a | 4.2 a | 4.2 a | 4.0 a |
| ‘Woody’ | 4.5 a | 4.5 a | 4.5 a | 4.5 a | 4.4 a | 4.5 a |
| ‘Animal’ | 4.5 ab | 4.7 a | 4.5 ab | 4.6 ab | 4.4 ab | 4.2 b |
| ‘Herbaceous’ | 4.8 a | 4.6 ab | 4.8 a | 4.6 ab | 4.5 ab | 4.3 b |
| ‘Spicy’ | 3.9 a | 4.0 a | 4.1 a | 3.9 a | 3.8 a | 3.8 a |
| ‘Yeasty’ | 4.7 a | 4.6 a | 4.7 a | 4.5 a | 4.5 a | 4.4 a |
| Mouthfeel | ||||||
| ‘Tingle’ | 5.7 a | 5.5 ab | 5.8 a | 5.6 ab | 5.5 ab | 5.2b |
| ‘Viscosity’ | 6.7 a | 6.8 a | 6.7 a | 6.6 a | 6.8 a | 6.7 a |
| ‘Weight’ | 6.1 a | 6.3 a | 6.2 a | 6.1 a | 6.3 a | 6.2 a |
| ‘Hot/Ethanol’ | 7.3 ab | 7.3 ab | 7.5 a | 7.3 ab | 7.0 bc | 6.7 c |
| ‘Roughness’ | 6.0 ab | 5.9 bc | 6.3 a | 5.8 bc | 6.0 ab | 5.6 c |
| ‘Astringent’ | 6.8 ab | 6.8 ab | 7.1 a | 6.8 ab | 6.6 b | 6.5 b |
| ‘Drying’ | 6.3 ab | 6.2 b | 6.6 a | 6.1 b | 6.2 b | 6.0 b |
| ‘Puckering’ | 6.0 bc | 6.0 bc | 6.5 a | 6.2 b | 6.1 bc | 5.8 c |
| ‘Sharp’ | 6.1 ab | 6.0 b | 6.4 a | 6.0 b | 6.0 bc | 5.5 c |
| ‘Mouthcoat’ | 6.0 b | 6.1 ab | 6.5 a | 6.2 ab | 6.3 ab | 6.0 b |
| ‘Round’ | 6.6 b | 6.9 ab | 6.7 b | 6.8 ab | 7.0 ab | 7.3 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Edwards, C.G.; Aplin, J.J. Application of Cool Fermentation Temperatures to Encourage Non-Saccharomyces Yeasts to Yield Lower Ethanol Concentrations in Wines. Fermentation 2022, 8, 421. https://doi.org/10.3390/fermentation8090421
Edwards CG, Aplin JJ. Application of Cool Fermentation Temperatures to Encourage Non-Saccharomyces Yeasts to Yield Lower Ethanol Concentrations in Wines. Fermentation. 2022; 8(9):421. https://doi.org/10.3390/fermentation8090421
Chicago/Turabian StyleEdwards, Charles G., and Jesse J. Aplin. 2022. "Application of Cool Fermentation Temperatures to Encourage Non-Saccharomyces Yeasts to Yield Lower Ethanol Concentrations in Wines" Fermentation 8, no. 9: 421. https://doi.org/10.3390/fermentation8090421