
Influence of Lactobacillus (LAB) Fermentation on the Enhancement of Branched Chain Amino Acids and Antioxidant Properties in Bran among Wheat By-Products

Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Samples and Reagents
2.2. Solid-State Fermentation (SSF)
2.3. Characterization of WB
2.3.1. Physicochemical Characteristics
2.3.2. Bioactive Components and Antioxidant Properties
2.3.3. Amino Acid Profiles
2.3.4. Morphological Characteristics
2.4. Statistical Analysis
3. Results
3.1. Physicochemical Characteristics
3.2. Amino Acid Profile
3.3. Bioactive Components and Antioxidant Activities
3.4. Morphological Characteristics
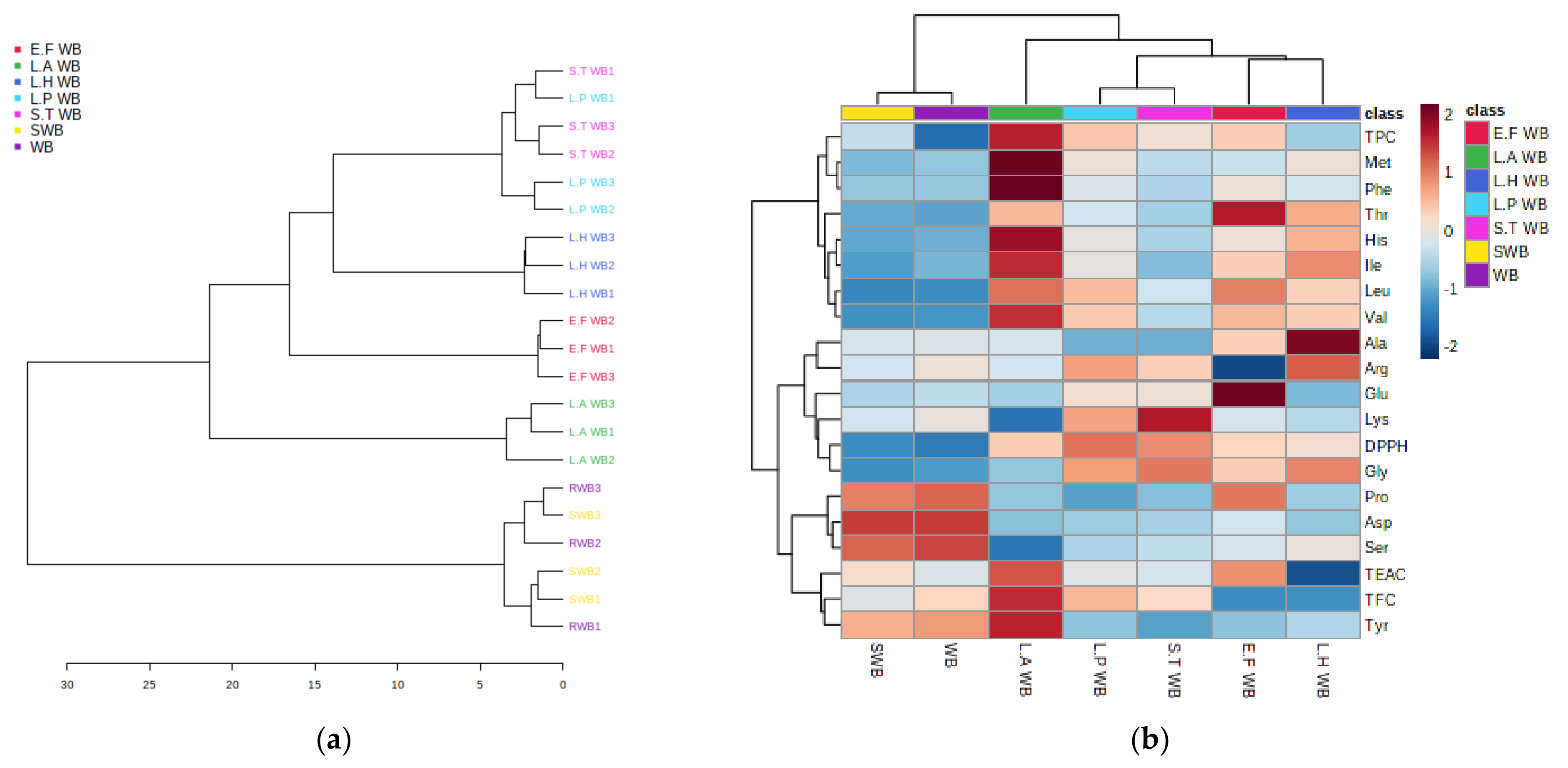
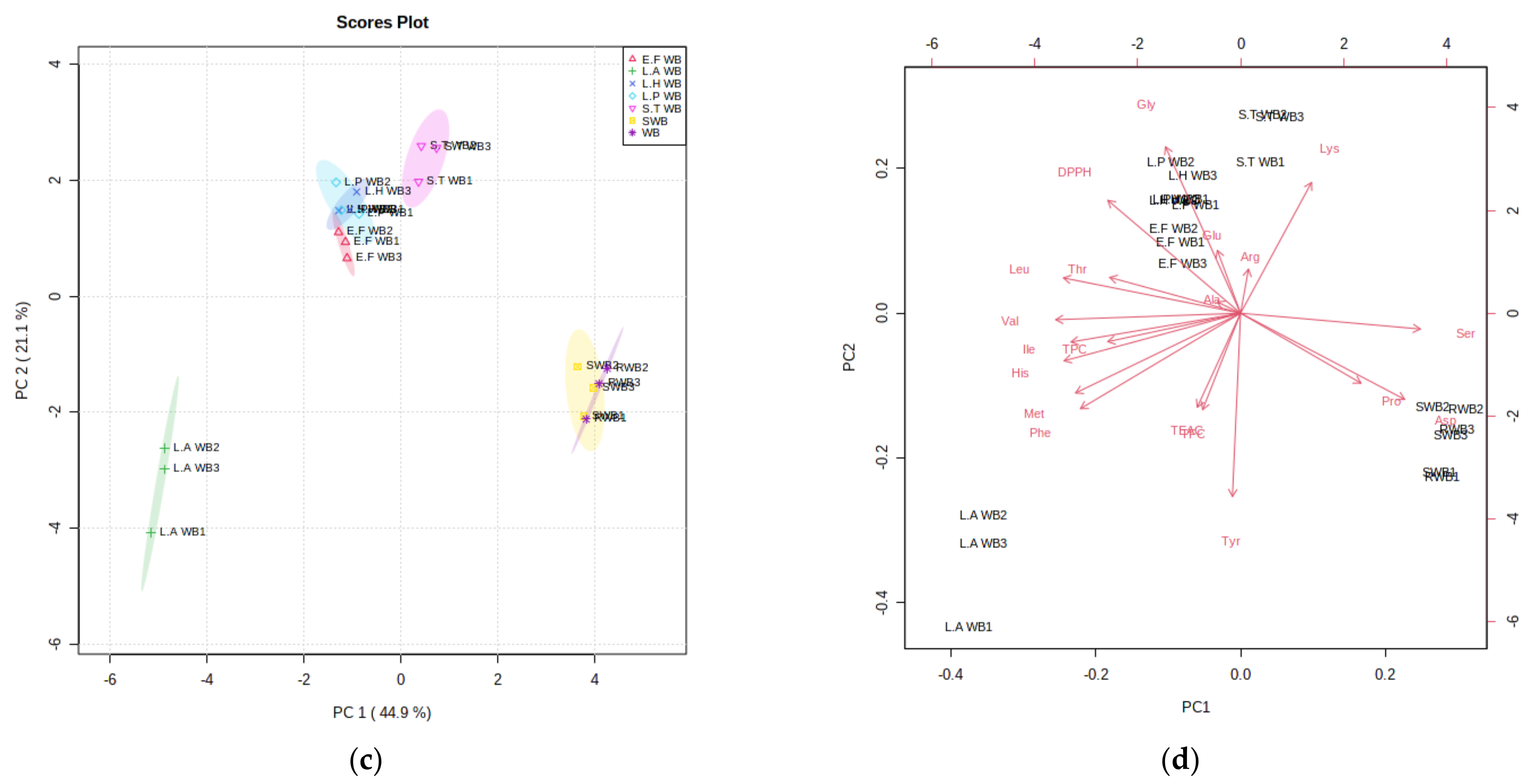
3.5. Statistical Assessment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT. FAO Statistics, Food and Agriculture Organization of the United Nations Rome, Italy. 2020. Available online: https://faostat.fao.org/ (accessed on 15 November 2022).
- Khader, B.F.; Yigezu, Y.A.; Duwayri, M.A.; Niane, A.A.; Shideed, K. Where in the value chain are we losing the most food? The case of wheat in Jordan. Food Secur. 2019, 11, 1009–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comino, E.; Dominici, L.; Perozzi, D. Do-it-yourself approach applied to the valorisation of a wheat milling industry’s by-product for producing bio-based material. J. Clean. Prod. 2021, 318, 128267. [Google Scholar] [CrossRef]
- Onipe, O.O.; Jideani, A.I.; Beswa, D. Composition and functionality of wheat bran and its application in some cereal food products. Int. J. Food Sci. Technol. 2015, 50, 2509–2518. [Google Scholar]
- Safaripour, M.; Hossain, K.G.; Ulven, C.A.; Pourhashem, G. Environmental impact tradeoff considerations for wheat bran-based biocomposite. Sci. Total Environ. 2021, 781, 146588. [Google Scholar] [CrossRef]
- Song, X.; Zhu, W.; Pei, Y.; Ai, Z.; Chen, J. Effects of wheat bran with different colors on the qualities of dry noodles. J. Cereal Sci. 2013, 58, 400–407. [Google Scholar]
- Nazari, J.; Yadegari, N.; Khodam, S.; Almasi-Hashian, A.; Amini, S. Effect of consumption of whole-wheat breads on FBS, HbA1c, and blood lipids in patients with type 2 diabetes. Prev. Nutr. Food Sci. 2021, 26, 269. [Google Scholar]
- Mosharraf, L.; Kadivar, M.; Shahedi, M. Effect of hydrothermaled bran on physicochemical, rheological and microstructural characteristics of Sangak bread. J. Cereal Sci. 2009, 49, 398–404. [Google Scholar] [CrossRef]
- Mao, M.; Wang, P.; Shi, K.; Lu, Z.; Bie, X.; Zhao, H.; Zhang, C.; Lv, F. Effect of solid state fermentation by Enterococcus faecalis M2 on antioxidant and nutritional properties of wheat bran. J. Cereal Sci. 2020, 94, 102997. [Google Scholar] [CrossRef]
- An, E.-M.; Lee, J.-K.; Choi, Y.-S.; Kim, Y.-H.; Shin, H.-S. Physicochemical properties of wheat bran in different areas prepared by a high-pressure homogenizer process. Korean J. Food Sci. Technol. 2014, 46, 404–409. [Google Scholar] [CrossRef] [Green Version]
- Nordlund, E.; Katina, K.; Aura, A.-M.; Poutanen, K. Changes in bran structure by bioprocessing with enzymes and yeast modifies the in vitro digestibility and fermentability of bran protein and dietary fibre complex. J. Cereal Sci. 2013, 58, 200–208. [Google Scholar] [CrossRef]
- Rezaei, S.; Najafi, M.A.; Haddadi, T. Effect of fermentation process, wheat bran size and replacement level on some characteristics of wheat bran, dough, and high-fiber Tafton bread. J. Cereal Sci. 2019, 85, 56–61. [Google Scholar] [CrossRef]
- Tanasković, S.J.; Šekuljica, N.; Jovanović, J.; Gazikalović, I.; Grbavčić, S.; Đorđević, N.; Sekulić, M.V.; Hao, J.; Luković, N.; Knežević-Jugović, Z. Upgrading of valuable food component contents and anti-nutritional factors depletion by solid-state fermentation: A way to valorize wheat bran for nutrition. J. Cereal Sci. 2021, 99, 103159. [Google Scholar] [CrossRef]
- Roasa, J.; De Villa, R.; Mine, Y.; Tsao, R. Phenolics of cereal, pulse and oilseed processing by-products and potential effects of solid-state fermentation on their bioaccessibility, bioavailability and health benefits: A review. Trends Food Sci. Technol. 2021, 116, 954–974. [Google Scholar] [CrossRef]
- Liang, Y.-F.; Long, Z.-X.; Zhang, Y.-J.; Luo, C.-Y.; Yan, L.-T.; Gao, W.-Y.; Li, H. The chemical mechanisms of the enzymes in the branched-chain amino acids biosynthetic pathway and their applications. Biochimie 2021, 184, 72–87. [Google Scholar] [CrossRef] [PubMed]
- Mutaguchi, Y.; Kasuga, K.; Kojima, I. Production of D-branched-chain amino acids by lactic acid bacteria carrying homologs to isoleucine 2-epimerase of Lactobacillus buchneri. Front. Microbiol. 2018, 9, 1540. [Google Scholar] [CrossRef] [Green Version]
- Maki, H.; Yamanaka-Okumura, H.; Katayama, T.; Ozawa, Y.; Hosoda, A.; Kurata, N.; Amemiya, F. Late evening snacks with branched-chain amino acids improve the Fischer ratio with patients liver cirrhosis at fasting in the next morning. Clin. Nutr. ESPEN 2019, 30, 138–144. [Google Scholar] [CrossRef]
- Yamanashi, K.; Kinugawa, S.; Fukushima, A.; Kakutani, N.; Takada, S.; Obata, Y.; Nakano, I.; Yokota, T.; Kitaura, Y.; Shimomura, Y. Branched-chain amino acid supplementation ameliorates angiotensin II-induced skeletal muscle atrophy. Life Sci. 2020, 250, 117593. [Google Scholar]
- El-Kalla, F.S.; Mansor, L.O.; El-Bassat, H.A.; Mishaal, S.; Attia, J.F. The effect of a late-evening protein-containing snack on nitrogen balance in cirrhotic patients. Tanta Med. J. 2014, 42, 47. [Google Scholar]
- Lee, N.H.; Jung, D.S.; Hong, J. Antioxidant properties and protective effects of aerial parts from Cnidium officinale makino on oxidative stress-induced neuronal cell death. Prev. Nutr. Food Sci. 2021, 26, 200. [Google Scholar] [CrossRef]
- OAC (Ed.) Official Methods of Analysis of AOAC International, 21st ed.; AOAC International: Washigton, DC, USA, 2019. [Google Scholar]
- Kim, S.M.; Aung, T.; Kim, M.J. Optimization of germination conditions to enhance the antioxidant activity in chickpea (Cicer arietimum L.) using response surface methodology. Korean J. Food Preserv. 2022, 29, 632–644. [Google Scholar] [CrossRef]
- Kim, M.J.; Kwak, H.S.; Kim, S.S. Effects of salinity on bacterial communities, Maillard reactions, isoflavone composition, antioxidation and antiproliferation in Korean fermented soybean paste (doenjang). Food Chem. 2018, 245, 402–409. [Google Scholar] [PubMed]
- Aung, T.; Kim, B.R.; Kim, M.J. Comparative Flavor Profile of Roasted Germinated Wheat (Triticum aestivum L.) Beverages Served Hot and Cold Using Electronic Sensors Combined with Chemometric Statistical Analysis. Foods 2022, 11, 3099. [Google Scholar] [PubMed]
- Xia, J.; Wishart, D.S. Using MetaboAnalyst 3.0 for comprehensive metabolomics data analysis. Curr. Protoc. Bioinform. 2016, 55, 14.10.11–14.10.91. [Google Scholar] [CrossRef] [PubMed]
- Antognoni, F.; Mandrioli, R.; Potente, G.; Taneyo Saa, D.L.; Gianotti, A. Changes in carotenoids, phenolic acids and antioxidant capacity in bread wheat doughs fermented with different lactic acid bacteria strains. Food Chem. 2019, 292, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Lei, M.; Samina, N.; Chen, L.; Liu, C.; Yin, T.; Yan, X.; Wu, C.; He, H.; Yi, C. Impact of Lactobacillus plantarum 423 fermentation on the antioxidant activity and flavor properties of rice bran and wheat bran. Food Chem. 2020, 330, 127156. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, X.; Xu, H.; Sun, Y.; Zhang, Y.; Wang, Y. Improvement of the nutritional, antioxidant and bioavailability properties of corn gluten-wheat bran mixture fermented with lactic acid bacteria and acid protease. LWT 2021, 144, 111161. [Google Scholar]
- Parthasarathy, A.; Cross, P.J.; Dobson, R.C.J.; Adams, L.E.; Savka, M.A.; Hudson, A.O. A Three-Ring Circus: Metabolism of the Three Proteogenic Aromatic Amino Acids and Their Role in the Health of Plants and Animals. Front. Mol. Biosci. 2018, 5, 29. [Google Scholar] [CrossRef]
- Görbitz, C.; Dalhus, B. Redetermination of L-Leucine at 120K. Acta Crystallogr. Sect. C Cryst. Struct. Commun. 1996, 52, 1754–1756. [Google Scholar]
- Holecek, M. Branched-chain amino acids in health and disease: Metabolism, alterations in blood plasma, and as supplements. Nutr. Metab. 2018, 15, 33. [Google Scholar] [CrossRef] [Green Version]
- Spaggiari, M.; Ricci, A.; Calani, L.; Bresciani, L.; Neviani, E.; Dall’Asta, C.; Lazzi, C.; Galaverna, G. Solid state lactic acid fermentation: A strategy to improve wheat bran functionality. LWT 2020, 118, 108668. [Google Scholar]
- Magalhães, L.M.; Segundo, M.A.; Reis, S.; Lima, J.L. Methodological aspects about in vitro evaluation of antioxidant properties. Anal. Chim. Acta 2008, 613, 1–19. [Google Scholar] [CrossRef]
- Wang, C.-C.; Chang, C.-H.; Chang, S.-C.; Fan, G.-J.; Lin, M.-J.; Yu, B.; Lee, T.-T. In vitro free radicals scavenging activity and antioxidant capacity of solid-state fermented wheat bran and its potential modulation of antioxidative molecular targets in chicken PBMC. Rev. Bras. De Zootec. 2016, 45, 451–457. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Wang, Z.; Yang, S.; Wang, X.; Li, L.; Sun, B.; Ma, S.; Zheng, S. Microstructure observation of multilayers separated from wheat bran. Grain Oil Sci. Technol. 2021, 4, 165–173. [Google Scholar] [CrossRef]
- De Paepe, K.; Verspreet, J.; Rezaei, M.N.; Hidalgo Martinez, S.; Meysman, F.; Van de Walle, D.; Dewettinck, K.; Raes, J.; Courtin, C.; Van de Wiele, T. Isolation of wheat bran-colonizing and metabolizing species from the human fecal microbiota. PeerJ 2019, 7, e6293. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhao, L.; Wang, L.; Liu, H. Microstructure-modified products from stone-milled wheat bran powder improve glycemic response and sustain colonic fermentation. Int. J. Biol. Macromol. 2020, 153, 1193–1201. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Fat *** (g/100 g) | Protein ** (g/100 g) | Ash *** (g/100 g) | L* Value *** | a* Value ** | b* Value ** |
|---|---|---|---|---|---|---|
| RWB | 1.99 ± 0.15 b | 16.13 ± 0.10 a | 4.74 ± 0.07 a | 64.00 ± 0.76 d | 7.50 ± 0.34 ab | 21.60 ± 1.07 a |
| SWB | 2.29 ± 0.27 c | 17.20 ± 0.18 c | 4.85 ± 0.05 ab | 64.33 ± 0.52 d | 7.07 ± 0.21 b | 21.00 ± 0.69 a |
| L.A WB | 2.89 ± 0.08 d | 17.10 ± 0.09 c | 4.91 ± 0.02 bc | 54.42 ± 1.56 a | 7.19 ± 0.21 b | 22.15 ± 0.44 bc |
| L.P WB | 2.99 ± 0.08 d | 16.39 ± 0.35 bc | 4.76 ± 0.00 a | 58.93 ± 0.42 c | 7.68 ± 0.22 ab | 22.87 ± 0.94 bc |
| L.H WB | 3.06 ± 0.10 d | 16.73 ± 0.27 bc | 4.85 ± 0.03 ab | 59.90 ± 2.15 c | 7.93 ± 0.70 bc | 24.13 ± 0.62 d |
| S.T WB | 1.50 ± 0.11 a | 17.14 ± 0.58 c | 4.93 ± 0.02 d | 59.88 ± 0.98 c | 7.86 ± 0.21 bc | 23.40 ± 0.36 c |
| E.F WB | 3.07 ± 0.08 d | 17.29 ± 0.37 c | 4.80 ± 0.05 ab | 56.70 ± 0.75 b | 8.32 ± 0.13 c | 22.84 ± 0.22 ab |
| Amino Acid 1 | RWB | SWB | L.A WB | L.P WB | L.H WB | S.T WB | E.F WB |
|---|---|---|---|---|---|---|---|
| Ala *** | 5.74 ± 0.42 b | 5.58 ± 0.20 b | 5.60 ± 0.23 b | 3.40 ± 0.43 a | 12.16 ± 2.12 c | 3.29 ± 0.22 a | 7.12 ± 0.69 b |
| Arg *** | 15.38 ± 0.45 c | 13.51 ± 0.55 b | 13.52 ± 1.58 b | 19.31 ± 0.33 e | 22.14 ± 1.26 f | 16.90 ± 0.55 c | 3.50 ± 0.12 a |
| Asp *** | 11.00 ± 0.76 c | 10.94 ± 0.63 c | 3.20 ± 0.17 a | 3.70 ± 0.31 a | 3.37 ± 0.37 a | 3.96 ± 0.25 a | 5.02 ± 0.25 b |
| Glu *** | 7.31 ± 0.07 b | 6.90 ± 0.13 b | 6.71 ± 0.18 ab | 8.71 ± 0.58 c | 6.06 ± 0.56 c | 8.58 ± 0.50 c | 14.24 ± 0.57 d |
| Gly *** | 3.33 ± 0.10 a | 3.16 ± 0.07 a | 3.95 ± 0.16 b | 6.07 ± 0.19 d | 6.35 ±0.26 d | 6.48 ± 0.48 d | 5.48± 0.32 c |
| His *** | 3.12 ± 0.13 a | 2.71 ± 0.09 a | 14.59 ± 1.10 e | 6.81 ± 0.19 c | 9.45 ± 0.11 d | 4.69 ± 0.22 b | 7.13 ± 0.36 c |
| Ile *** | 1.29 ± 0.03 a | 1.15 ± 0.04 a | 2.56 ± 0.05 d | 1.73 ± 0.25 b | 2.21 ± 0.09 c | 1.31 ± 0.08 a | 1.92 ± 0.11 b |
| Leu *** | 1.61 ±0.04 a | 1.49 ± 0.07 a | 7.70 ± 0.40 d | 6.25 ± 0.28 c | 5.74 ± 0.32 c | 4.28 ± 0.15 b | 7.48 ± 0.46 d |
| Lys *** | 3.49 ± 0.12 bc | 3.37 ± 0.12 b | 2.65 ± 0.12 a | 3.88 ± 0.43 c | 3.26 ± 0.39 b | 4.42 ± 0.25 d | 3.39 ± 0.19 b |
| Met *** | 0.41 ± 0.08 a | 0.29 ± 0.15 a | 3.57 ± 0.08 d | 1.23 ± 0.07 c | 1.26 ± 0.03 c | 0.76 ± 0.02 b | 0.87 ± 0.07 b |
| Phe *** | 1.36 ± 0.11 a | 1.32 ± 0.21 a | 16.46 ± 0.74 e | 4.30 ± 0.23 c | 3.79 ± 0.22 c | 2.31 ± 0.25 b | 5.16 ± 0.25 d |
| Pro *** | 19.82 ± 0.91 c | 19.02 ± 0.19 c | 11.67 ± 0.45 b | 10.07 ± 0.66 a | 12.16 ± 0.20 b | 11.45 ± 0.94 bc | 19.19 ± 1.45 c |
| Ser *** | 28.90 ± 0.15 e | 27.54 ± 0.83 e | 11.48 ± 0.40 a | 17.38 ± 1.32 b | 20.55 ± 0.31 d | 18.47 ± 1.10 bc | 19.44 ± 1.05 cd |
| Thr *** | 1.86 ± 0.09 a | 1.99 ± 0.42 a | 5.97 ± 0.98 c | 3.97 ± 0.46 b | 6.27 ± 0.60 c | 3.03 ± 0.43 b | 8.90 ± 0.69 e |
| Tyr *** | 2.06 ± 0.22 c | 1.89 ± 0.10 c | 2.92 ± 0.53 d | 0.49 ± 0.12 ab | 0.74 ± 0.01 ab | 0.15 ± 0.06 a | 0.48 ± 0.11 ab |
| Val *** | 2.18 ± 0.16 a | 2.12 ± 0.17 a | 7.21 ± 0.37 d | 5.08 ± 0.55 c | 5.02 ± 0.52 c | 3.57 ± 0.35 b | 5.36 ± 0.18 c |
| Samples | TPC *** (mg GAE/ g) | TFC *** (mg CE/ g) | DPPH *** (µM TE/ g) | TEAC *** (mM TE/ g) |
|---|---|---|---|---|
| RWB | 1.61 ± 0.62 a | 0.88 ± 0.11 bc | 8.08 ± 0.17 a | 8.11 ± 0.33 bc |
| SWB | 2.07 ± 0.27 ab | 0.84 ± 0.06 ab | 8.18 ± 0.28 a | 8.47 ± 0.71 bc |
| L.A WB | 2.80 ± 0.27 c | 1.01 ± 0.08 c | 8.98 ± 0.14 bc | 9.88 ± 2.07 d |
| L.P WB | 2.36 ± 0.16 bc | 0.91 ± 0.06 bc | 9.34 ± 0.12 d | 8.18 ± 0.54 bc |
| L.H WB | 1.98 ± 0.21 ab | 0.71 ± 0.07 a | 8.86 ± 0.08 b | 5.93 ± 1.24 a |
| S.T WB | 2.23 ± 0.19 bc | 0.87 ± 0.12 bc | 9.24 ± 0.10 cd | 8.06 ± 0.07 ab |
| E.F WB | 2.33 ± 0.09 bc | 0.71 ± 0.08 a | 8.94 ± 0.02 b | 9.33 ± 0.42 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aung, T.; Park, S.-S.; Kim, M.-J. Influence of Lactobacillus (LAB) Fermentation on the Enhancement of Branched Chain Amino Acids and Antioxidant Properties in Bran among Wheat By-Products. Fermentation 2022, 8, 732. https://doi.org/10.3390/fermentation8120732
Aung T, Park S-S, Kim M-J. Influence of Lactobacillus (LAB) Fermentation on the Enhancement of Branched Chain Amino Acids and Antioxidant Properties in Bran among Wheat By-Products. Fermentation. 2022; 8(12):732. https://doi.org/10.3390/fermentation8120732
Chicago/Turabian StyleAung, Thinzar, Seung-Soo Park, and Mi-Jeong Kim. 2022. "Influence of Lactobacillus (LAB) Fermentation on the Enhancement of Branched Chain Amino Acids and Antioxidant Properties in Bran among Wheat By-Products" Fermentation 8, no. 12: 732. https://doi.org/10.3390/fermentation8120732