
Exploring the Core Microbiota of Four Different Traditional Fermented Beverages from the Colombian Andes

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
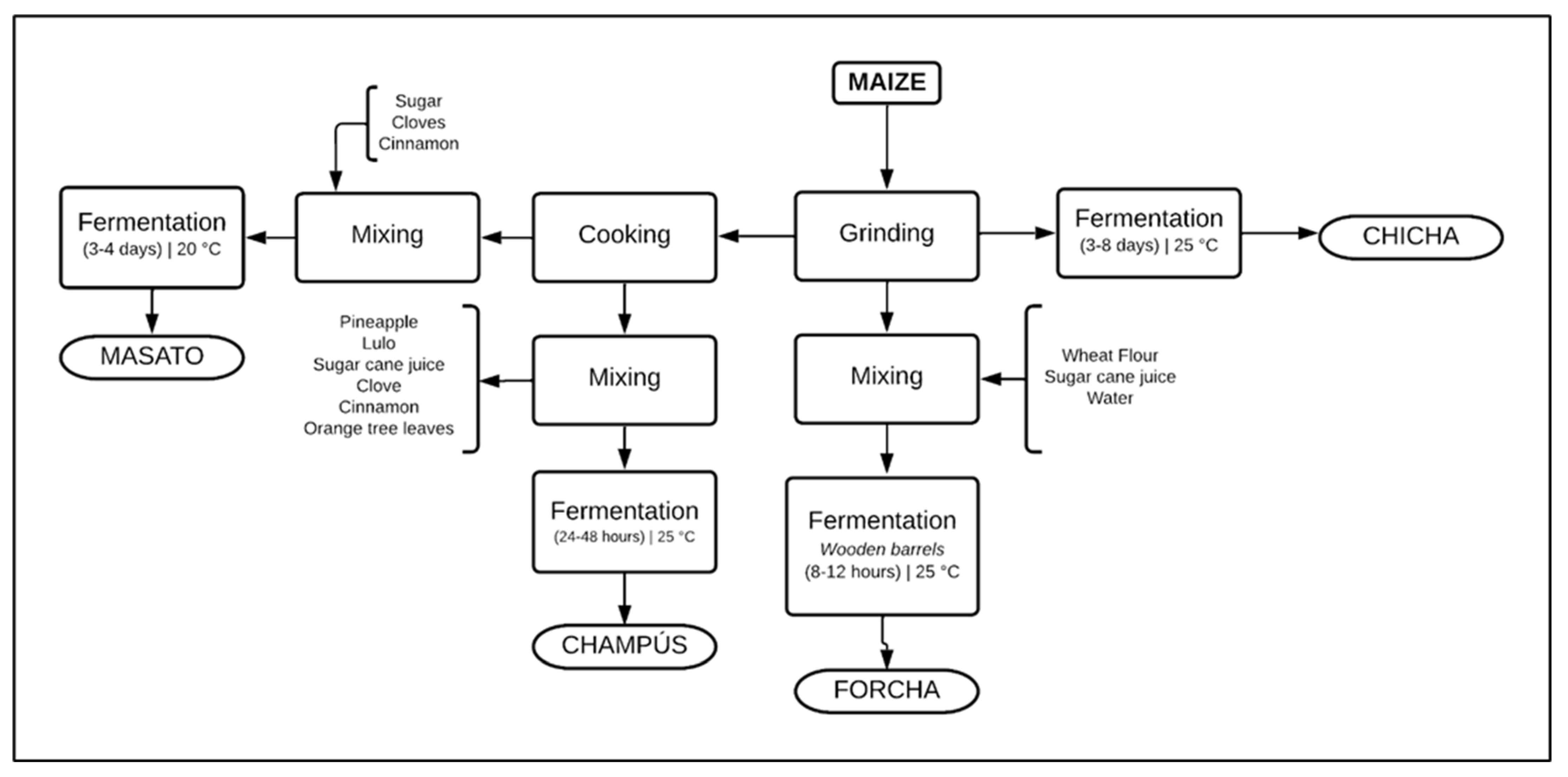
2.1. Samples
2.2. Physicochemical Characterization
2.3. Antioxidant Capacity Assays
2.3.1. ABTS Radical Cation (ABTS•+) Scavenging Activity Assay
2.3.2. DPPH Radical Scavenging Ability Assay
2.3.3. Total Phenol Index
2.4. Organic Acid, Ethanol, and Sugar Content
2.5. Microbiological Analyses
2.6. Genomic DNA Extraction
DNA Sample Quality and Quantity Specification
2.7. Sequence Analysis and Species Identification
2.8. Alpha Diversity Analysis
2.9. Statistical Analysis
3. Results
3.1. Sequence Analysis
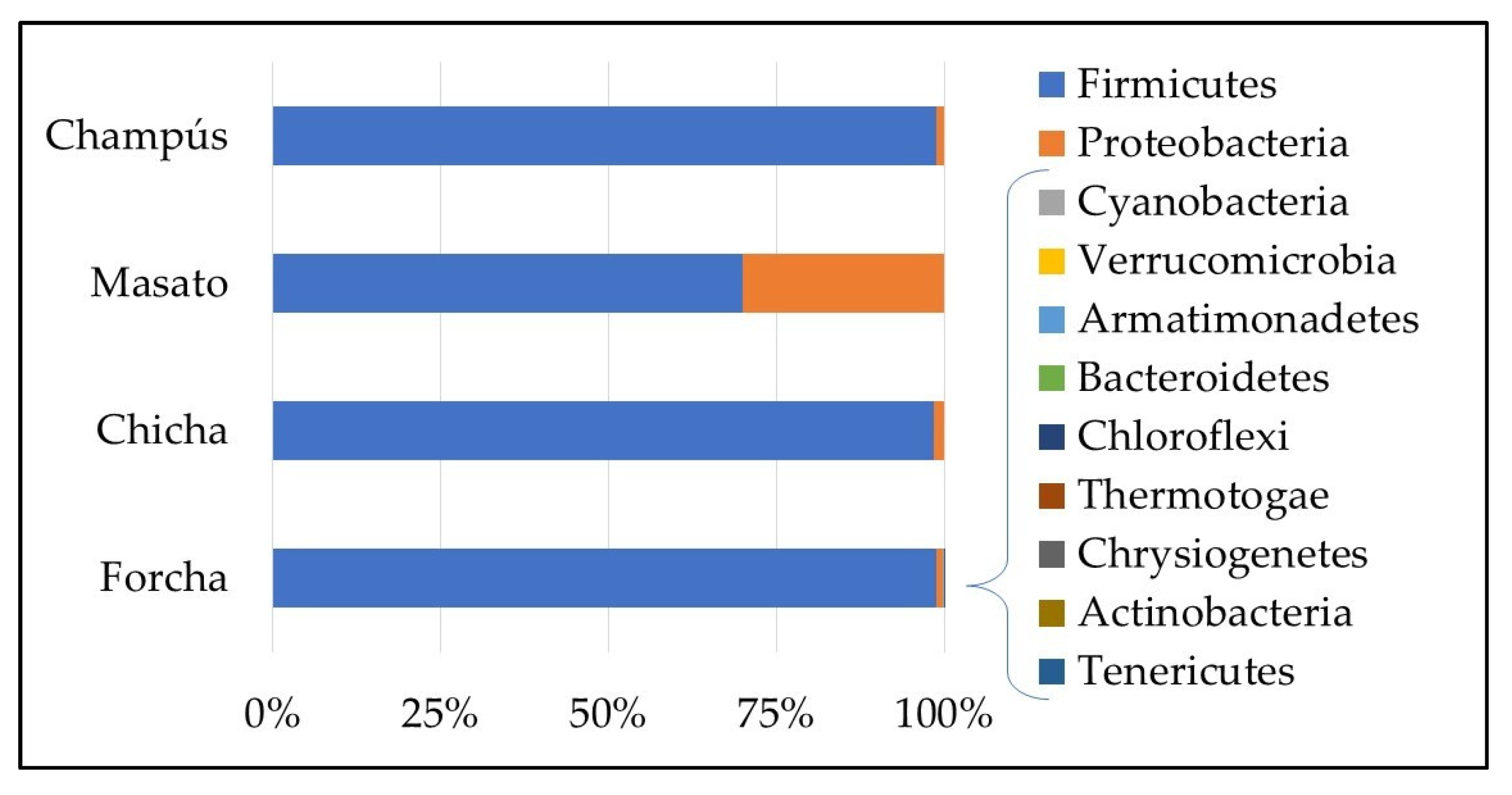
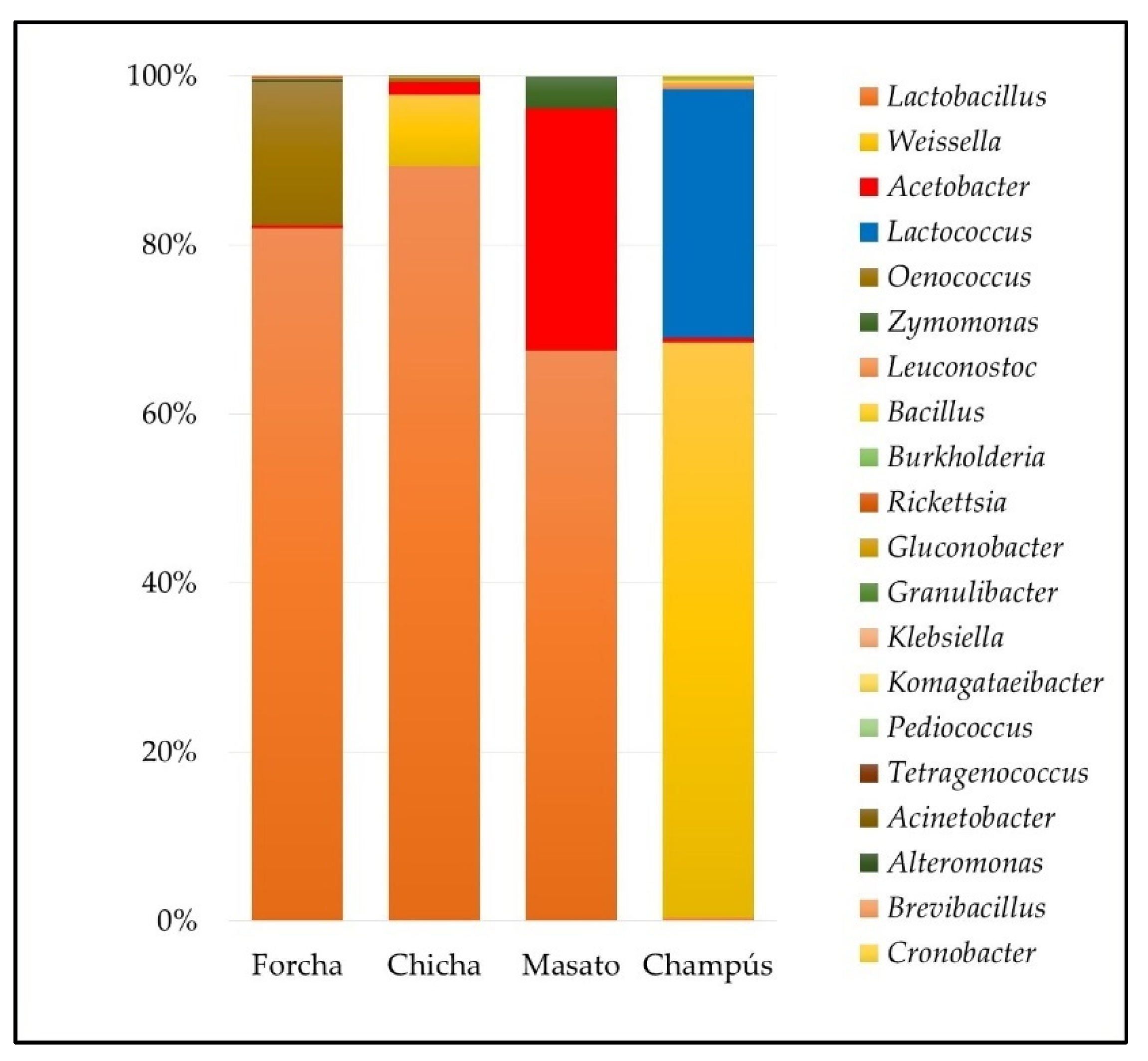
3.2. Bacterial Communities of the Different Fermented Beverages
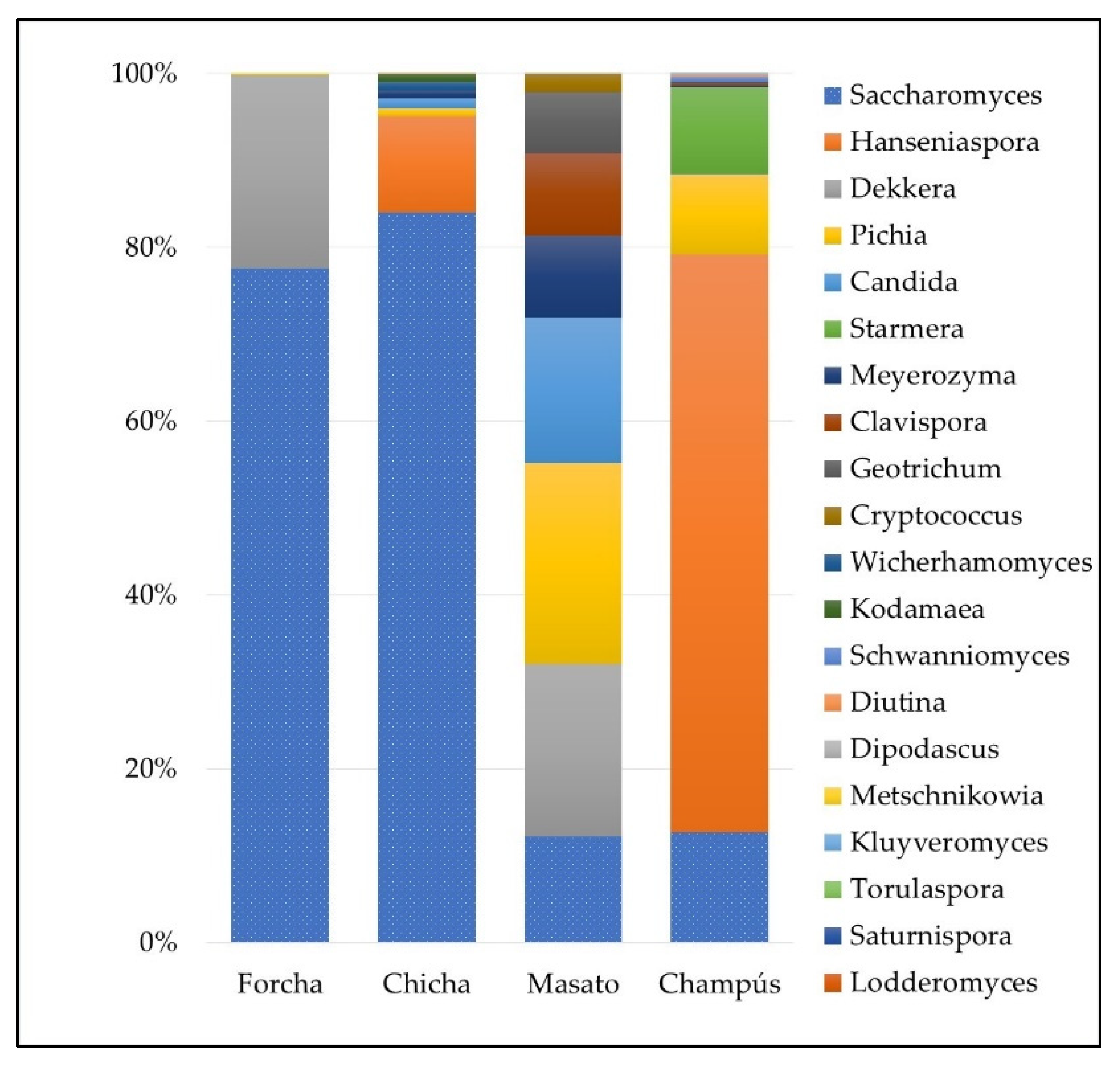
3.3. Fungal Communities of the Different Fermented Beverages
3.4. Species Richness and Diversity Index
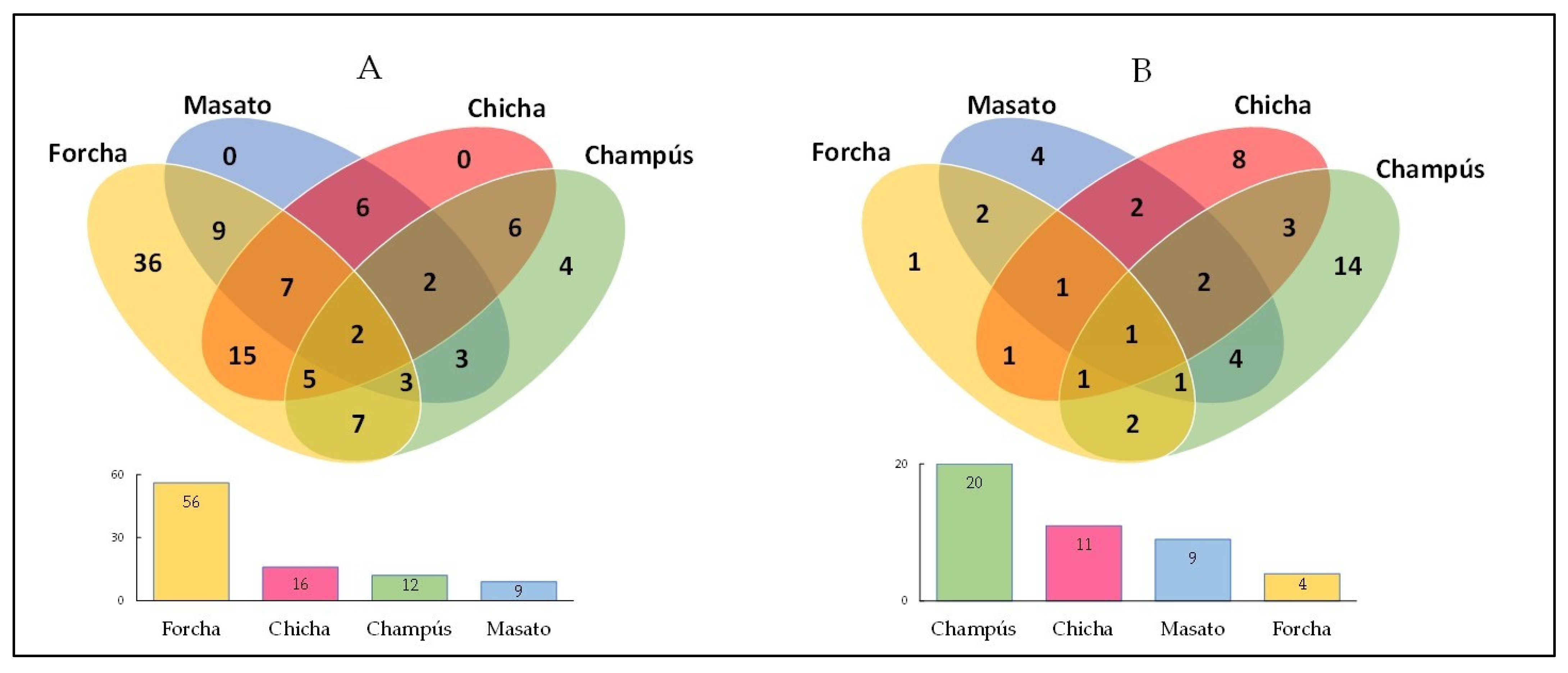
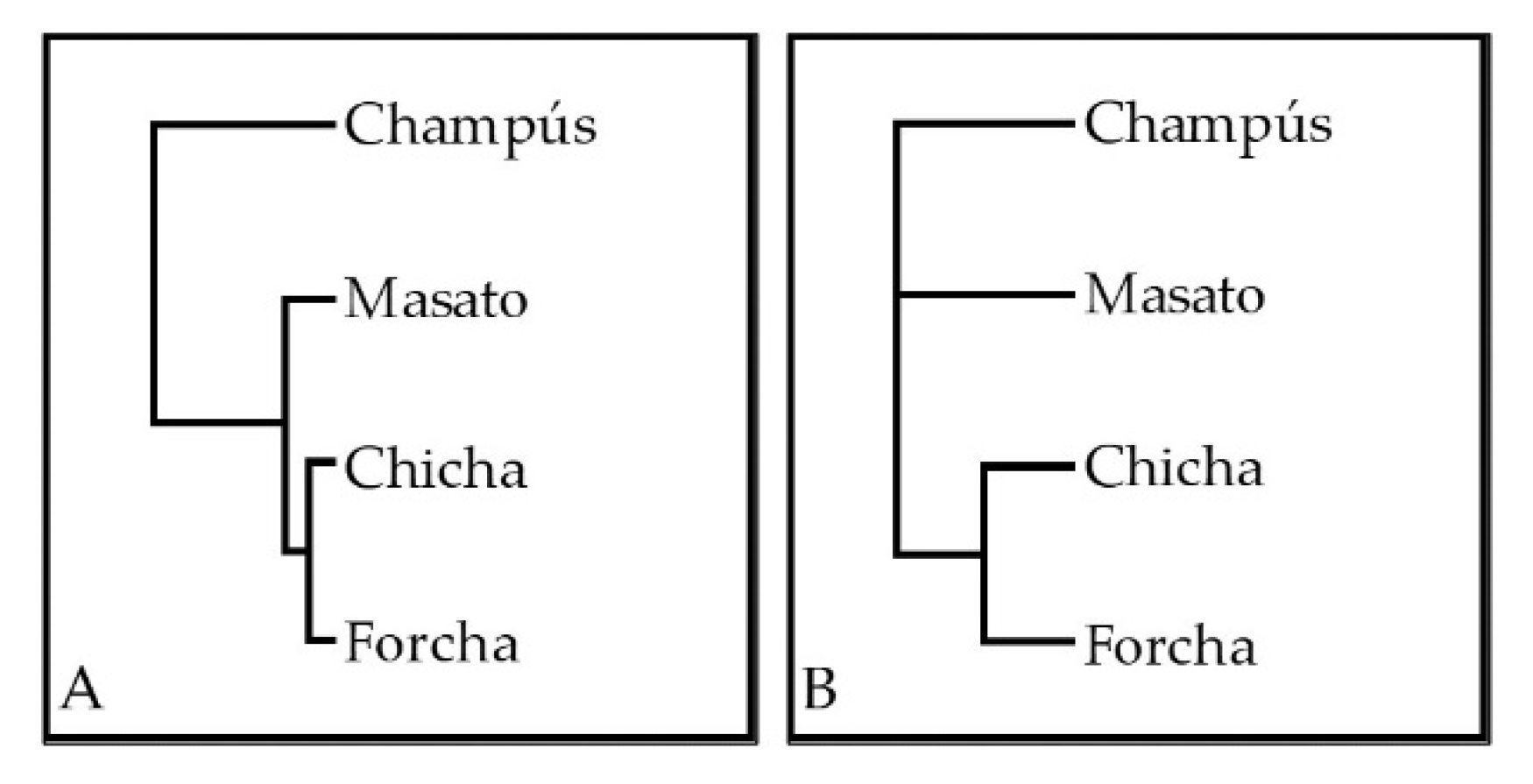
3.5. Similarities and Differences among the Microbial Communities of the Fermented Beverages
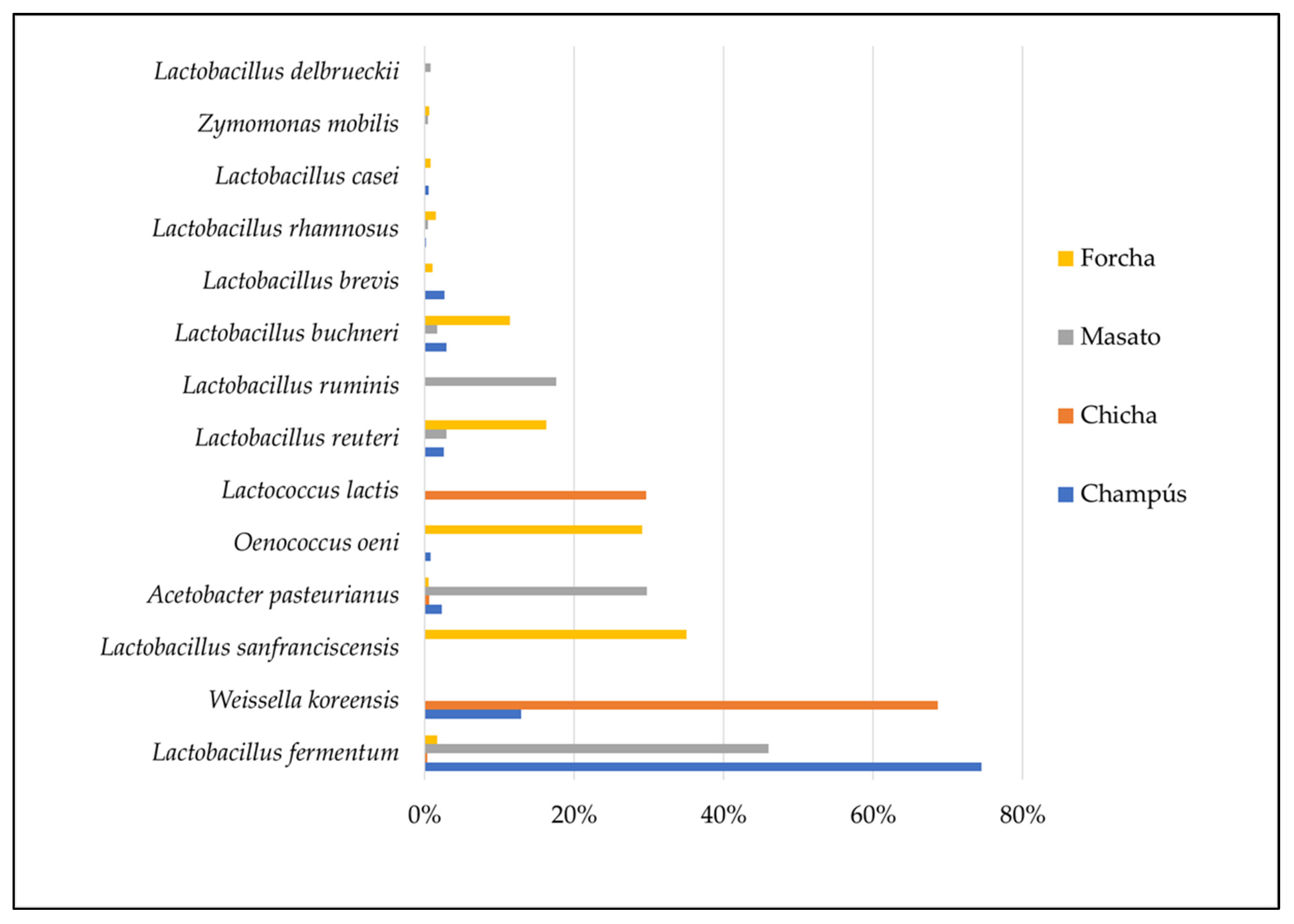
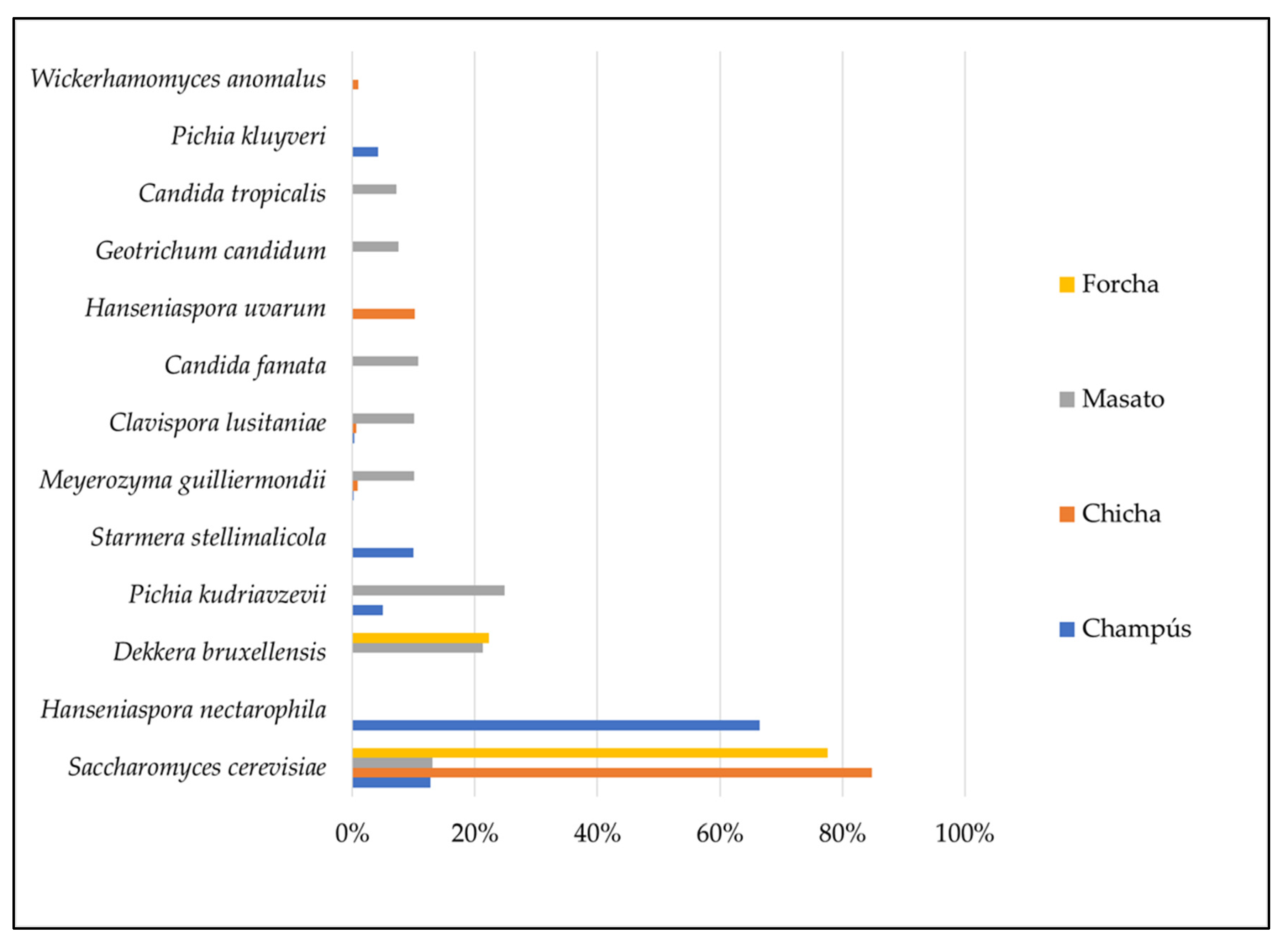
3.6. Dominant Microbiota Identification
3.7. Chemical Characteristics of the Beverages
Anti-Radical Capacity and Total Phenol Index
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Altay, F.; Karbancioglu-Güler, F.; Daskaya-Dikmen, C.; Heperkan, D. A review on traditional Turkish fermented non-alcoholic beverages: Microbiota, fermentation process and quality characteristics. Int. J. Food Microbiol. 2013, 167, 44–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swain, M.R.; Anandharaj, M.; Ray, R.C.; Praveen Rani, R. Fermented Fruits and Vegetables of Asia: A Potential Source of Probiotics. Biotechnol. Res. Int. 2014, 2014, 250424. [Google Scholar] [CrossRef] [PubMed]
- Ray, M.; Ghosh, K.; Singh, S.; Mondal, K.C. Folk to functional: An explorative overview of rice-based fermented foods and beverages in India. J. Ethn. Foods 2016, 3, 5–18. [Google Scholar] [CrossRef] [Green Version]
- Todorov, S.D.; Holzapfel, W.H. Traditional Cereal Fermented Foods as Sources of Functional Microorganisms; Elsevier Ltd.: Amsterdam, The Netherlands, 2014; ISBN 9781782420248. [Google Scholar]
- Aloys, N.; Angeline, N. Traditional fermented foods and beverages in Burundi. Food Res. Int. 2009, 42, 588–594. [Google Scholar] [CrossRef]
- Satish Kumar, R.; Kanmani, P.; Yuvaraj, N.; Paari, K.A.; Pattukumar, V.; Arul, V. Traditional Indian fermented foods: A rich source of lactic acid bacteria. Int. J. Food Sci. Nutr. 2013, 64, 415–428. [Google Scholar] [CrossRef]
- Robledo-Márquez, K.; Ramírez, V.; González-Córdova, A.F.; Ramírez-Rodríguez, Y.; García-Ortega, L.; Trujillo, J. Research opportunities: Traditional fermented beverages in Mexico. Cultural, microbiological, chemical, and functional aspects. Food Res. Int. 2021, 147, 110482. [Google Scholar] [CrossRef] [PubMed]
- Chaves-López, C.; Serio, A.; Grande-Tovar, C.D.; Cuervo-Mulet, R.; Delgado-Ospina, J.; Paparella, A. Traditional fermented foods and beverages from a microbiological and nutritional perspective: The Colombian Heritage. Compr. Rev. Food Sci. Food Saf. 2014, 13, 1031–1048. [Google Scholar] [CrossRef] [Green Version]
- Chaves-López, C.; Serio, A.; Paparella, A.; Martuscelli, M.; Corsetti, A.; Tofalo, R.; Suzzi, G. Impact of microbial cultures on proteolysis and release of bioactive peptides in fermented milk. Food Microbiol. 2014, 42, 117–121. [Google Scholar] [CrossRef]
- Chen, R.; He, Y.; Li, X.; Ramos, J.; Li, M.; Liu, L. Fermented maize beverages as ritual offerings: Investigating elite drinking during Classic Maya period at Copan, Honduras. J. Anthropol. Archaeol. 2022, 65, 101373. [Google Scholar] [CrossRef]
- Vinton, S.D.; Perry, L.; Reinhard, K.J.; Santoro, C.M.; Teixeira-Santos, I. Impact of Empire Expansion on Household Diet: The Inka in Northern Chile’s Atacama Desert. PLoS ONE 2009, 4, e8069. [Google Scholar] [CrossRef]
- My Colombian Cocina. Available online: https://www.mycolombiancocina.com/ (accessed on 1 December 2022).
- Quintero-Angel, M.; Mendoza-Salazar, D.M.; Martínez-Girón, J. Food fears and risk of loss of food heritage: A little-explored effect of food modernity and times of pandemic. Int. J. Gastron. Food Sci. 2022, 28, 100499. [Google Scholar] [CrossRef]
- Osorio-Cadavid, E.; Chaves-López, C.; Tofalo, R.; Paparella, A.; Suzzi, G. Detection and identification of wild yeasts in Champús, a fermented Colombian maize beverage. Food Microbiol. 2008, 25, 771–777. [Google Scholar] [CrossRef] [PubMed]
- Chaves-López, C.; Rossi, C.; Maggio, F.; Paparella, A.; Serio, A. Changes Occurring in Spontaneous Maize Fermentation: An Overview. Fermentation 2020, 6, 36. [Google Scholar] [CrossRef] [Green Version]
- Botta, C.; Cocolin, L. Microbial dynamics and biodiversity in table olive fermentation: Culture-dependent and -independent approaches. Front. Microbiol. 2012, 3, 245. [Google Scholar] [CrossRef] [Green Version]
- Supasil, R.; Suttisansanee, U.; Santivarangkna, C.; Tangsuphoom, N.; Khemthong, C.; Chupeerach, C.; On-nom, N. Improvement of Sourdough and Bread Qualities by Fermented Water of Asian Pears and Assam Tea Leaves with Co-Cultures of Lactiplantibacillus plantarum and Saccharomyces cerevisiae. Foods 2022, 11, 2071. [Google Scholar] [CrossRef]
- Chaves-López, C.; Serio, A.; Mazzarrino, G.; Martuscelli, M.; Scarpone, E.; Paparella, A. Control of household mycoflora in fermented sausages using phenolic fractions from olive mill wastewaters. Int. J. Food Microbiol. 2015, 207, 49–56. [Google Scholar] [CrossRef]
- Ilyasov, I.R.; Beloborodov, V.L.; Selivanova, I.A.; Terekhov, R.P. ABTS/PP Decolorization Assay of Antioxidant Capacity Reaction Pathways. Int. J. Mol. Sci. 2020, 21, 1131. [Google Scholar] [CrossRef] [Green Version]
- Villa-Rodríguez, J.A.; Molina-Corral, F.J.; Ayala-Zavala, J.F.; Olivas, G.I.; González-Aguilar, G.A. Effect of maturity stage on the content of fatty acids and antioxidant activity of “Hass” avocado. Food Res. Int. 2011, 44, 1231–1237. [Google Scholar] [CrossRef]
- Ioannone, F.; Di Mattia, C.D.; De Gregorio, M.; Sergi, M.; Serafini, M.; Sacchetti, G. Flavanols, proanthocyanidins and antioxidant activity changes during cocoa (Theobroma cacao L.) roasting as affected by temperature and time of processing. Food Chem. 2015, 174, 263–269. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-Learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Abarenkov, K.; Zirk, A.; Pöhönen, R.; Ivanov, F.; Nilsson, R.H.; Kõljalg, U. UNITE QIIME release for Fungi 2. Version 10.05.2021. UNITE Community 2021, 7, 1264763. [Google Scholar] [CrossRef]
- Chaves-López, C.; Serio, A.; Delgado-Ospina, J.; Rossi, C.; Grande-Tovar, C.D.; Paparella, A. Exploring the bacterial microbiota of colombian fermented maize dough “Masa agria” (Maiz añejo). Front. Microbiol. 2016, 7, 1168. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Elizaquível, P.; Pérez-Cataluña, A.; Yépez, A.; Aristimuño, C.; Jiménez, E.; Cocconcelli, P.S.; Vignolo, G.; Aznar, R. Pyrosequencing vs. culture-dependent approaches to analyze lactic acid bacteria associated to chicha, a traditional maize-based fermented beverage from Northwestern Argentina. Int. J. Food Microbiol. 2015, 198, 9–18. [Google Scholar] [CrossRef]
- Yurkov, A.M.; Kemler, M.; Begerow, D. Assessment of yeast diversity in soils under different management regimes. Fungal Ecol. 2012, 5, 24–35. [Google Scholar] [CrossRef]
- Vargas Escobar, E.A.; Baena García, D.; Vargas Sánchez, J.E. Análisis de estabilidad y adaptabilidad de híbridos de maíz de alta calidad proteica en diferentes zonas agroecológicas de Colombia. Acta Agronómica 2016, 65, 72–79. [Google Scholar] [CrossRef]
- Igual, M.; Ramires, S.; Mosquera, L.H.; Martínez-Navarrete, N. Optimization of spray drying conditions for lulo (Solanum quitoense L.) pulp. Powder Technol. 2014, 256, 233–238. [Google Scholar] [CrossRef]
- Chaves-López, C.; Serio, A.; Osorio-Cadavid, E.; Paparella, A.; Suzzi, G. Volatile compounds produced in wine by Colombian wild Saccharomyces cerevisiae strains. Ann. Microbiol. 2009, 59, 733–740. [Google Scholar] [CrossRef]
- Sieuwerts, S.; de Bok, F.A.M.; Hugenholtz, J.; van Hylckama Vlieg, J.E. Unraveling microbial interactions in food fermentations: From classical to genomics approaches. Appl. Environ. Microbiol. 2008, 74, 4997–5007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minervini, F.; De Angelis, M.; Di Cagno, R.; Gobbetti, M. Ecological parameters influencing microbial diversity and stability of traditional sourdough. Int. J. Food Microbiol. 2014, 171, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Minervini, F.; Dinardo, F.R.; De Angelis, M.; Gobbetti, M. Tap water is one of the drivers that establish and assembly the lactic acid bacterium biota during sourdough preparation. Sci. Rep. 2019, 9, 570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Armendáriz, B.; Cardoso-Ugarte, G.A. Traditional fermented beverages in Mexico: Biotechnological, nutritional, and functional approaches. Food Res. Int. 2020, 136, 109307. [Google Scholar] [CrossRef] [PubMed]
- Guerra, L.S.; Cevallos-Cevallos, J.M.; Weckx, S.; Ruales, J. Traditional Fermented Foods from Ecuador: A Review with a Focus on Microbial Diversity. Foods 2022, 11, 1854. [Google Scholar] [CrossRef]
- Eisenbach, L.; Geissler, A.J.; Ehrmann, M.A.; Vogel, R.F. Comparative genomics of Lactobacillus sakei supports the development of starter strain combinations. Microbiol. Res. 2019, 221, 1–9. [Google Scholar] [CrossRef]
- Lau, S.W.; Chong, A.Q.; Chin, N.L.; Talib, R.A.; Basha, R.K. Sourdough Microbiome Comparison and Benefits. Microorganisms 2021, 9, 1355. [Google Scholar] [CrossRef]
- Gänzle, M.G.; Zheng, J. Lifestyles of sourdough lactobacilli—Do they matter for microbial ecology and bread quality? Int. J. Food Microbiol. 2019, 302, 15–23. [Google Scholar] [CrossRef]
- Lim, K.; Koh, J. Chapter 5—Fermented foods and probiotic beverages in Korea. In Probiotic Beverages; Panda, S.K., Kellershohn, J., Russell, I.B.T.-P.B., Eds.; Academic Press: London, UK, 2021; pp. 61–80. ISBN 978-0-12-818588-9. [Google Scholar]
- Lorentzen, M.P.G.; Lucas, P.M. Distribution of Oenococcus oeni populations in natural habitats. Appl. Microbiol. Biotechnol. 2019, 103, 2937–2945. [Google Scholar] [CrossRef] [Green Version]
- Mwizerwa, H.; George Ooko Abong, G.O.; Mbugua, S.K.; Okoth, M.W.; Gacheru, P.; Muiru, M.; Obura, B.; Viljoen, B. Profiling of Microbial Content and Growth in Fermented Maize Based Products from Western Kenya. Curr. Res. Nutr. Food Sci. 2018, 6, 509–519. [Google Scholar] [CrossRef]
- Maske, B.L.; de Melo Pereira, G.V.; Vale, A.D.S.; de Carvalho Neto, D.P.; Karp, S.G.; Viesser, J.A.; De Dea Lindner, J.; Pagnoncelli, M.G.; Soccol, V.T.; Soccol, C.R. A review on enzyme-producing lactobacilli associated with the human digestive process: From metabolism to application. Enzyme Microb. Technol. 2021, 149, 109836. [Google Scholar] [CrossRef]
- Park, Y.E.; Kim, M.S.; Shim, K.W.; Kim, Y.-I.; Chu, J.; Kim, B.-K.; Choi, I.S.; Kim, J.Y. Effects of Lactobacillus plantarum Q180 on Postprandial Lipid Levels and Intestinal Environment: A Double-Blind, Randomized, Placebo-Controlled, Parallel Trial. Nutrients 2020, 12, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.S.; Kim, S.H. Anti-inflammatory and Anti-osteoporotic Potential of Lactobacillus plantarum A41 and L. fermentum SRK414 as Probiotics. Probiotics Antimicrob. Proteins 2020, 12, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Azhar, N.S.; Md Zin, N.H.; Hamid, T. Lactococcus lactis strain A5 producing nisin-like bacteriocin active against Gram positive and negative bacteria. Trop. life Sci. Res. 2017, 28, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Woo, C.; Jung, S.; Fugaban, J.I.I.; Bucheli, J.E.V.; Holzapfel, W.H.; Todorov, S.D. Bacteriocin production by Leuconostoc citreum ST110LD isolated from organic farm soil, a promising biopreservative. J. Appl. Microbiol. 2021, 131, 1226–1239. [Google Scholar] [CrossRef] [PubMed]
- Xiraphi, N.; Georgalaki, M.; Rantsiou, K.; Cocolin, L.; Tsakalidou, E.; Drosinos, E.H. Purification and characterization of a bacteriocin produced by Leuconostoc mesenteroides E131. Meat Sci. 2008, 80, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Yeong, M.S.; Hee, M.S.; Choon, C.H. Characterization of High-Ornithine-Producing Weissella koreensis DB1 Isolated from Kimchi and Its Application in Rice Bran Fermentation as a Starter Culture. Foods 2020, 9, 1545. [Google Scholar] [CrossRef]
- Bassi, D.; Orrù, L.; Cabanillas Vasquez, J.; Cocconcelli, P.S.; Fontana, C. Peruvian chicha: A Focus on the Microbial Populations of This Ancient Maize-Based Fermented Beverage. Microorganisms 2020, 8, 93. [Google Scholar] [CrossRef] [Green Version]
- Fusco, V.; Quero, G.M.; Cho, G.-S.; Kabisch, J.; Meske, D.; Neve, H.; Bockelmann, W.; Franz, C.M.A.P. The genus Weissella: Taxonomy, ecology and biotechnological potential. Front. Microbiol. 2015, 6, 155. [Google Scholar] [CrossRef] [Green Version]
- Fessard, A.; Remize, F. Why Are Weissella spp. Not Used as Commercial Starter Cultures for Food Fermentation? Fermentation 2017, 3, 38. [Google Scholar] [CrossRef]
- Ayivi, R.D.; Gyawali, R.; Krastanov, A.; Aljaloud, S.O.; Worku, M.; Tahergorabi, R.; Silva, R.C.; Ibrahim, S.A. Lactic Acid Bacteria: Food Safety and Human Health Applications. Dairy 2020, 1, 202–232. [Google Scholar] [CrossRef]
- Papalexandratou, Z.; Vrancken, G.; De Bruyne, K.; Vandamme, P.; De Vuyst, L. Spontaneous organic cocoa bean box fermentations in Brazil are characterized by a restricted species diversity of lactic acid bacteria and acetic acid bacteria. Food Microbiol. 2011, 28, 1326–1338. [Google Scholar] [CrossRef] [PubMed]
- Elhalis, H.; Cox, J.; Frank, D.; Zhao, J. The crucial role of yeasts in the wet fermentation of coffee beans and quality. Int. J. Food Microbiol. 2020, 333, 108796. [Google Scholar] [CrossRef]
- Xu, H.; Hong, J.H.; Kim, D.; Jin, Y.H.; Pawluk, A.M.; Mah, J.-H. Evaluation of Bioactive Compounds and Antioxidative Activity of Fermented Green Tea Produced via One- and Two-Step Fermentation. Antioxidants 2022, 11, 1425. [Google Scholar] [CrossRef]
- De Roos, J.; De Vuyst, L. Acetic acid bacteria in fermented foods and beverages. Curr. Opin. Biotechnol. 2018, 49, 115–119. [Google Scholar] [CrossRef]
- Da Silva, G.A.R.; Oliveira, S.S.d.S.; Lima, S.F.; do Nascimento, R.P.; Baptista, A.R.d.S.; Fiaux, S.B. The industrial versatility of Gluconobacter oxydans: Current applications and future perspectives. World J. Microbiol. Biotechnol. 2022, 38, 134. [Google Scholar] [CrossRef]
- Leupold, S.; Hubmann, G.; Litsios, A.; Meinema, A.C.; Takhaveev, V.; Papagiannakis, A.; Niebel, B.; Janssens, G.; Siegel, D.; Heinemann, M. Saccharomyces cerevisiae goes through distinct metabolic phases during its replicative lifespan. Elife 2019, 8, e41046. [Google Scholar] [CrossRef]
- Jespersen, L. Occurrence and taxonomic characteristics of strains of Saccharomyces cerevisiae predominant in African indigenous fermented foods and beverages. FEMS Yeast Res. 2003, 3, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, M.; Pérez-Través, L.; Sangorrín, M.; Barrio, E.; Querol, A.; Lopes, C. Saccharomyces uvarum is responsible for the traditional fermentation of apple chicha in Patagonia. FEMS Yeast Res. 2017, 17, fow109. [Google Scholar] [CrossRef] [Green Version]
- Mendoza, L.M.; Neef, A.; Vignolo, G.; Belloch, C. Yeast diversity during the fermentation of Andean chicha: A comparison of high-throughput sequencing and culture-dependent approaches. Food Microbiol. 2017, 67, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Piló, F.B.; Carvajal-Barriga, E.J.; Guamán-Burneo, M.C.; Portero-Barahona, P.; Dias, A.M.M.; Freitas, L.F.D.d.; Gomes, F.d.C.O.; Rosa, C.A. Saccharomyces cerevisiae populations and other yeasts associated with indigenous beers (chicha) of Ecuador. Braz. J. Microbiol. 2018, 49, 808–815. [Google Scholar] [CrossRef] [PubMed]
- Grijalva-Vallejos, N.; Aranda, A.; Matallana, E. Evaluation of yeasts from Ecuadorian chicha by their performance as starters for alcoholic fermentations in the food industry. Int. J. Food Microbiol. 2020, 317, 108462. [Google Scholar] [CrossRef] [PubMed]
- Čadež, N.; Pagnocca, F.C.; Raspor, P.; Rosa, C.A. Hanseniaspora nectarophila sp. nov., a yeast species isolated from ephemeral flowers. Int. J. Syst. Evol. Microbiol. 2014, 64, 2364–2369. [Google Scholar] [CrossRef]
- Kamilari, E.; Mina, M.; Karallis, C.; Tsaltas, D. Metataxonomic Analysis of Grape Microbiota During Wine Fermentation Reveals the Distinction of Cyprus Regional terroirs. Front. Microbiol. 2021, 12, 726483. [Google Scholar] [CrossRef]
- Jatmiko, Y.D.; Lopes, M.d.B.; Barton, M.D. Molecular Identification of Yeasts Isolated from Dadih by RFLP-PCR and Assessment on Their Ability in Utilizing Lactate. Microbiol. Indones. 2012, 6, 5. [Google Scholar] [CrossRef] [Green Version]
- Da Cunha, T.; Ferraz, L.P.; Wehr, P.P.; Kupper, K.C. Antifungal activity and action mechanisms of yeasts isolates from citrus against Penicillium italicum. Int. J. Food Microbiol. 2018, 276, 20–27. [Google Scholar] [CrossRef] [Green Version]
- N’guessan, K.F.; Brou, K.; Jacques, N.; Casaregola, S.; Dje, K.M. Identification of yeasts during alcoholic fermentation of tchapalo, a traditional sorghum beer from Côte d’Ivoire. Antonie Van Leeuwenhoek 2011, 99, 855–864. [Google Scholar] [CrossRef]
- Katongole, J.N. The Microbial Succession in Indigenous Fermented Maize Products; University of Free State: Bloemfontein, South Africa, 2008. [Google Scholar]
- Oro, L.; Canonico, L.; Marinelli, V.; Ciani, M.; Comitini, F. Occurrence of Brettanomyces bruxellensis on Grape Berries and in Related Winemaking Cellar. Front. Microbiol. 2019, 10, 415. [Google Scholar] [CrossRef] [Green Version]
- Oladeji, B.S. Effects of Fermentation and Roasting on Natural Antioxidants in Maize. Eur. J. Agric. Food Sci. 2022, 4, 95–100. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Svensson, L.; Sekwati-Monang, B.; Lutz, D.L.; Schieber, A.; Gänzle, M.G. Phenolic Acids and Flavonoids in Nonfermented and Fermented Red Sorghum (Sorghum bicolor (L.) Moench). J. Agric. Food Chem. 2010, 58, 9214–9220. [Google Scholar] [CrossRef] [PubMed]
- Gänzle, M.G. Enzymatic and bacterial conversions during sourdough fermentation. Food Microbiol. 2014, 37, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Gancel, A.-L.; Alter, P.; Dhuique-Mayer, C.; Ruales, J.; Vaillant, F. Identifying Carotenoids and Phenolic Compounds In Naranjilla (Solanum quitoense Lam. Var. Puyo Hybrid), an Andean Fruit. J. Agric. Food Chem. 2008, 56, 11890–11899. [Google Scholar] [CrossRef]
- Kaplan, N.M.; Palmer, B.F. Nutritional and Health Benefits of Beer. Am. J. Med. Sci. 2000, 320, 320–326. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fermented Beverage | ||||
|---|---|---|---|---|
| Chicha | Forcha | Champús | Masato | |
| Lactic acid bacteria (Log CFU mL−1) | 2.4 ± 0.3 a | 6.10 ± 0.4 c | 3.8 ± 0.1 ab | 5.2 ± 0.2 bc |
| Acetic bacteria (Log CFU mL−1) | 2.3 ± 0.2 a | 2.1 ± 0.1 a | <1.0 | <1.0 |
| Yeast (Log CFU mL−1) | 5.1 ± 0.3 b | 4.2 ± 0.4 ab | 6.8 ± 0.2 b | 2.1 ± 0.2 a |
| Coliforms | <1.0 | <1.0 | <1.0 | <1.0 |
| Fermented Beverages | Observed Species | Chao1 | Shannon | |||
|---|---|---|---|---|---|---|
| Bacteria | Fungi | Bacteria | Fungi | Bacteria | Fungi | |
| Chicha | 16 | 11 | 21 | 13 | 1.42 | 0.89 |
| Forcha | 56 | 4 | 72 | 4 | 2.41 | 0.77 |
| Champús | 12 | 20 | 14 | 21 | 1.04 | 1.66 |
| Masato | 9 | 9 | 14 | 11 | 1.88 | 2.85 |
| Parameters | Fermented Beverage | |||
|---|---|---|---|---|
| Chicha | Forcha | Champús | Masato | |
| pH | 3.50 ± 0.08 a | 3.76 ± 0.75 ab | 3.92 ± 0.15 ab | 4.35 ± 0.15 b |
| Lactic acid (g/L) | 16.7 ± 1.7 c | 8.8 ± 3.0 bc | 5.5 ± 2.8 ab | 0.3 ± 0.1 a |
| Ash (%) | 2.17 ± 0.82 b | 0.46 ± 0.21 a | 0.96 ± 0.25 a | 0.53 ± 0.01 a |
| Protein (%) | 5.30 ± 0.7 b | 1.41 ± 0.3 a | 1.12 ± 0.1 a | 3.40 ± 0.6 ab |
| Saccharose (g/L) | 4.8 ± 2.9 a | 28.1 ± 10.3 b | 127.2 ± 18.8 c | 117.1 ± 1.8 c |
| Glucose (g/L) | 2.9 ± 2.4 a | 23.0 ± 11.5 b | 24.0 ± 16.4 b | 3.2 ± 0.1 a |
| Fructose (g/L) | 14.1 ± 7.2 b | 2.8 ± 1.1 a | 21.1 ± 14.0 b | 1.0 ± 0.1 a |
| Soluble solids (%) | 8.5 ± 1.5 a | 26.3 ± 7.6 c | 16.0 ± 4.0 bc | 12.5 ± 2.5 ab |
| Ethanol (%) | 9.9 ± 1.2 c | 2.9 ± 1.1 bc | 0.5 ± 0.2 a | 0.2 ± 0.1 a |
| Beverage | DPPH (µM TE/L) | ABTS (µM TE/L) | TPI (mg GAE/L) |
|---|---|---|---|
| Chicha | 15.24 ± 3.76 b | 15.48 ± 1.05 b | 546 ± 33 b |
| Forcha | 3.08 ± 1.28 a | 0.55 ± 0.42 a | 0.87 ± 0.33 ac |
| Champús | 1.99 ± 0.45 a | 13.13 ± 9.19 b | 258 ± 76 bc |
| Masato | 2.20 ± 0.89 a | 0.38 ± 0.20 a | 0.43 ± 0.15 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgado-Ospina, J.; Puerta-Polanco, L.F.; Grande-Tovar, C.D.; Cuervo, R.A.; Navia-Porras, D.P.; Poveda-Perdomo, L.G.; Fernández-Daza, F.F.; Chaves-López, C. Exploring the Core Microbiota of Four Different Traditional Fermented Beverages from the Colombian Andes. Fermentation 2022, 8, 733. https://doi.org/10.3390/fermentation8120733
Delgado-Ospina J, Puerta-Polanco LF, Grande-Tovar CD, Cuervo RA, Navia-Porras DP, Poveda-Perdomo LG, Fernández-Daza FF, Chaves-López C. Exploring the Core Microbiota of Four Different Traditional Fermented Beverages from the Colombian Andes. Fermentation. 2022; 8(12):733. https://doi.org/10.3390/fermentation8120733
Chicago/Turabian StyleDelgado-Ospina, Johannes, Luisa Fernanda Puerta-Polanco, Carlos David Grande-Tovar, Raúl Alberto Cuervo, Diana Paola Navia-Porras, Luis Gabriel Poveda-Perdomo, Fabián Felipe Fernández-Daza, and Clemencia Chaves-López. 2022. "Exploring the Core Microbiota of Four Different Traditional Fermented Beverages from the Colombian Andes" Fermentation 8, no. 12: 733. https://doi.org/10.3390/fermentation8120733