
Underwater Shock Wave-Enhanced Cavitation to Induce Morphological Changes and Cell Permeabilization in Microscopic Fungi
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Subject
2.2. Fungal Growth and Conidia Harvesting
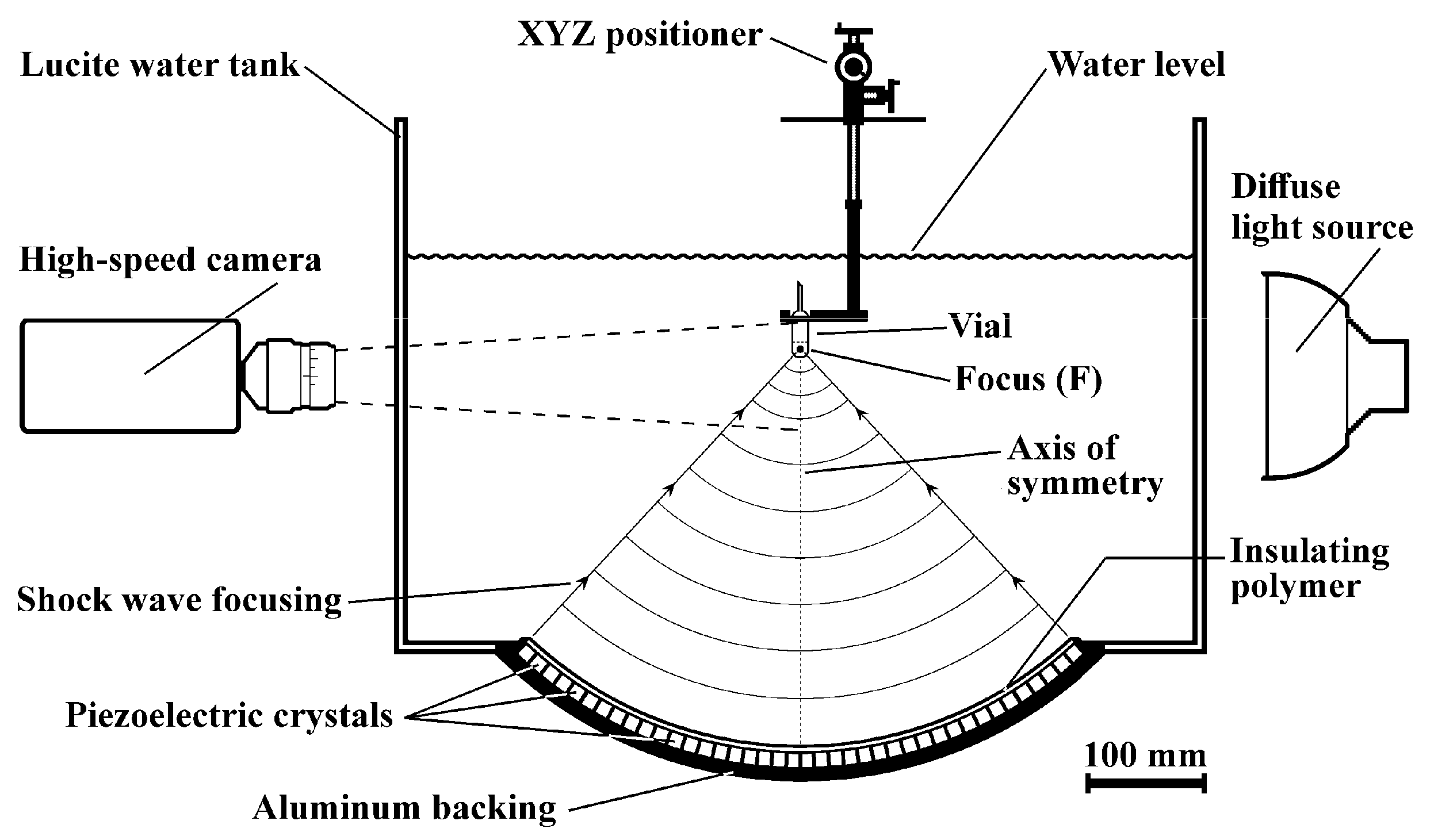
2.3. The Shock Wave Generator
2.4. Shock Wave Exposure
2.5. High-Speed Imaging
2.6. Electron Microscopy Sample Preparation
2.7. Protein Release Analysis
2.8. Electrophoretic Light Scattering
3. Results
3.1. Morphological Changes in A. niger Observed through SEM
3.2. Internal Changes of A. niger Observed through TEM
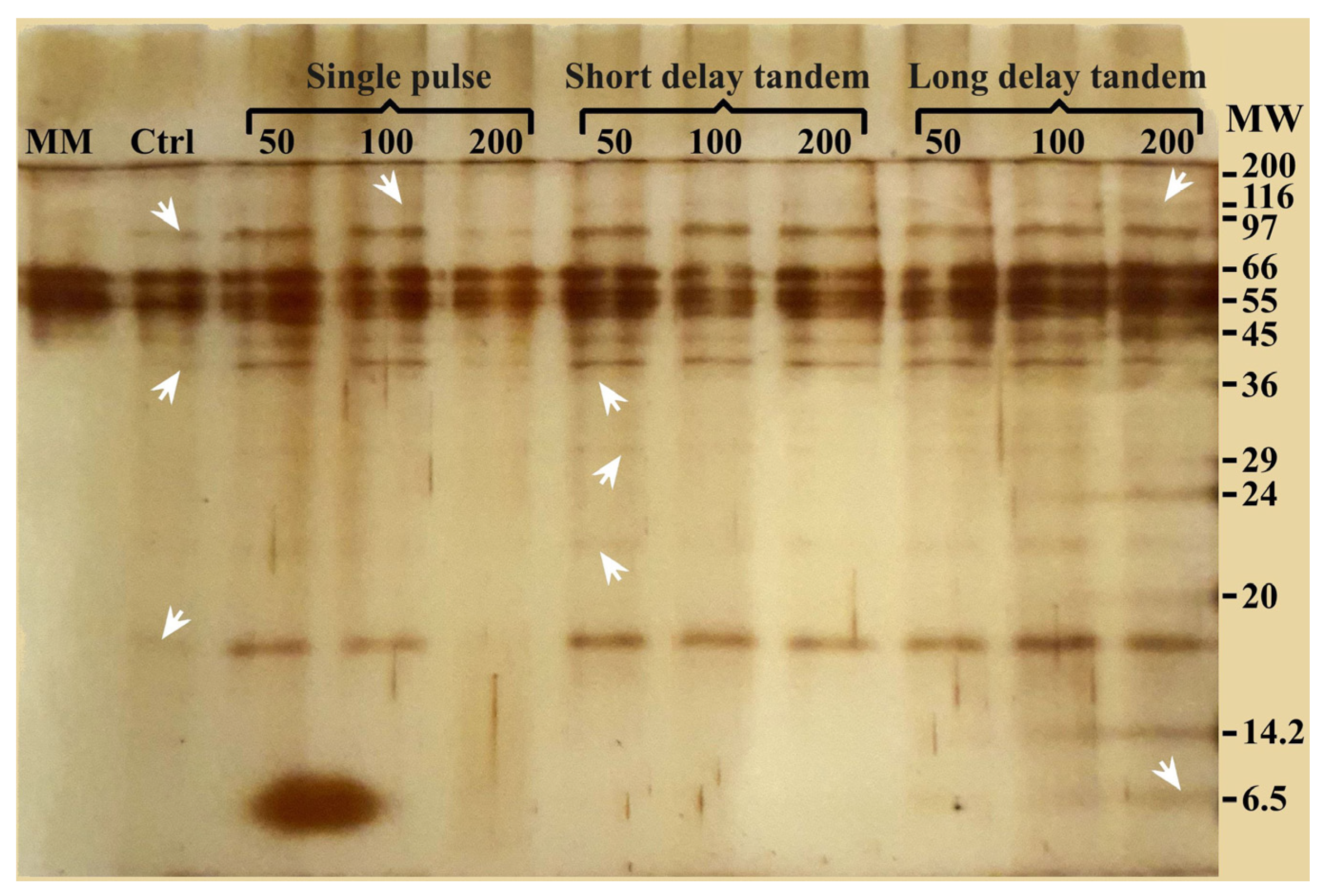
3.3. Analysis of Released Protein
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Field, J.E. The physics of liquid impact, shock wave interactions with cavities, and the implications to shock wave lithotripsy. Phys. Med. Biol. 1991, 36, 1475–1484. [Google Scholar] [CrossRef]
- Chaussy, C.; Brendel, W.; Schmiedt, E. Extracorporeally induced destruction of kidney stones by shock waves. Lancet 1980, 316, 1265–1268. [Google Scholar] [CrossRef]
- Chaussy, C.; Schmiedt, E.; Jocham, B.; Brendel, W.; Forssmann, B.; Walther, V. First clinical experience with extracorporeally induced destruction of kidney stones by shock waves. J. Urol. 1982, 127, 417–420. [Google Scholar] [CrossRef]
- Philipp, A.; Delius, M.; Scheffczyk, C.; Vogel, A.; Lauterborn, W. Interaction of lithotripter-generated shock waves with air bubbles. J. Acoust. Soc. Am. 1993, 93, 2496–2509. [Google Scholar] [CrossRef]
- Lauterborn, W.; Ohl, C.D. The peculiar dynamics of cavitation bubbles. Appl. Sci. Res. 1998, 58, 63–76. [Google Scholar] [CrossRef]
- Brujan, E.A.; Ikeda, T.; Matsumoto, Y. On the pressure of cavitation bubbles. Exp. Thermal Fluid Sci. 2008, 32, 1188–1191. [Google Scholar] [CrossRef]
- Klaseboer, E.; Fong, S.W.; Turangan, C.K.; Khoo, B.C.; Szeri, A.J.; Calvisi, M.L.; Sankin, G.N.; Zhong, P. Interaction of lithotripter shockwaves with single inertial cavitation bubbles. J. Fluid Mech. 2007, 593, 33–56. [Google Scholar] [CrossRef]
- Choi, M.J.; Coleman, A.J.; Saunders, J.E. The influence of fluid properties and pulse amplitude on bubble dynamics in the field of a shock wave lithotripter. J. Phys. Med. Biol. 1993, 38, 1561–1573. [Google Scholar] [CrossRef]
- Prosperetti, A.; Lezzi, A. Bubble dynamics in a compressible liquid. Part 1. First-order theory. J. Fluid Mech. 1986, 168, 457–478. [Google Scholar] [CrossRef]
- Zhong, P.; Lin, H.; Xi, X.; Zhu, S.; Bhogte, E.S. Shock wave-inertial microbubble interaction: Methodology, physical characterization, and bioeffect study. J. Acoust. Soc. Am. 1999, 105, 1997–2009. [Google Scholar] [CrossRef]
- Coleman, A.J.; Saunders, J.E.; Crum, L.A.; Dyson, M. Acoustic cavitation generated by an extracorporeal shock wave lithotripter. Ultrasound Med. Biol. 1987, 13, 69–76. [Google Scholar] [CrossRef]
- Crum, L.A. Cavitation microjets as a contributory mechanism for renal calculi disintegration in ESWL. J. Urol. 1988, 140, 1587–1590. [Google Scholar] [CrossRef]
- Delacrétaz, G.; Rink, K.; Pittomvils, G.; Lafaut, J.P.; Vandeursen, H.; Boving, R. Importance of the implosion of ESWL-induced cavitation bubbles. Ultrasound Med. Biol. 1995, 21, 97–103. [Google Scholar] [CrossRef]
- Zhong, P.; Cocks, F.H.; Cioanta, I.; Preminger, G.M. Controlled, forced collapse of cavitation bubbles for improved stone fragmentation during shock wave lithotripsy. J. Urol. 1997, 158, 2323–2328. [Google Scholar] [CrossRef]
- Cathignol, E.; Tavakkoli, J.; Birer, A.; Arefiev, A. Comparison between the effects of cavitation induced by two different pressure-time shock waveform pulses. IEEE Trans. Ultrason. Ferroelectr. Freq. Control 1998, 45, 788–799. [Google Scholar] [CrossRef]
- Xi, X.; Zhong, P. Improvement of stone fragmentation during shock-wave lithotripsy using a combined EH/PEAA shock-wave generator-in vitro experiments. Ultrasound Med. Biol. 2000, 26, 457–467. [Google Scholar] [CrossRef]
- Loske, A.M.; Prieto, F.E.; Fernández, F.; van Cauwelaert, J. Tandem shock wave cavitation enhancement for extracorporeal lithotripsy. Phys. Med. Biol. 2002, 47, 3945–3957. [Google Scholar] [CrossRef]
- Loske, A.M.; Fernández, F.; Zendejas, H.; Paredes, M.; Castaño-Tostado, E. Dual pulse shock wave lithotripsy: In vitro and in vivo study. J. Urol. 2005, 174, 2388–2392. [Google Scholar] [CrossRef]
- Fernández, F.; Fernández, G.; Loske, A.M. Treatment time reduction using tandem shockwaves for lithotripsy: An in vivo study. J. Endourol. 2009, 23, 1247–1253. [Google Scholar] [CrossRef]
- Ohl, C.D.; Ikink, R. Shock-wave-induced jetting of micron-size bubble. Phys. Rev. Lett. 2003, 90, 214502. [Google Scholar] [CrossRef]
- Kodama, T.; Takayama, K. Dynamic behavior of bubbles during extracorporeal shock-wave lithotripsy. Ultrasound Med. Biol. 1998, 24, 723–738. [Google Scholar] [CrossRef]
- Tzanakis, I.; Eskin, D.G.; Georgoulas, A.; Fytanidis, D.K. Incubation pit analysis and calculation of the hydrodynamic impact pressure from the implosion of an acoustic cavitation bubble. Ultrason. Sonochem. 2014, 21, 866–878. [Google Scholar] [CrossRef]
- Loske, A.M.; Campos-Guillen, J.; Fernández, F.; Castaño-Tostado, E. Enhanced shock wave-assisted transformation of Escherichia coli. Ultrasound Med. Biol. 2011, 37, 502–510. [Google Scholar] [CrossRef]
- Magaña-Ortíz, D.; Coconi-Linares, N.; Ortiz-Vazquez, E.; Fernández, F.; Loske, A.M.; Gómez-Lim, M.A. A novel and highly efficient method for genetic transformation of fungi employing shock waves. Fungal Genet. Biol. 2013, 56, 9–16. [Google Scholar] [CrossRef]
- Molina, G.A.; González-Fuentes, F.; Loske, A.M.; Fernández, F.; Estevez, M. Shock wave-assisted extraction of phenolic acids and flavonoids from Eysenhardtia polystachya heartwood: A novel method and its comparison with conventional methodologies. Ultrason. Sonochem. 2020, 61, 104809. [Google Scholar] [CrossRef]
- Church, C.C. A Theoretical study of cavitation generated by an extracorporeal shock wave lithotripter. J. Acoust. Soc. Am. 1989, 86, 215–227. [Google Scholar] [CrossRef]
- Cairns, T.C.; Barthel, L.; Meyer, V. Something old, something new: Challenges and developments in Aspergillus niger biotechnology. Essays Biochem. 2021, 65, 213–224. [Google Scholar] [CrossRef]
- Meyer, V.; Wu, B.; Ram, A.F.J. Aspergillus as a multi-purpose cell factory: Current status and perspectives. Biotechnol. Lett. 2011, 33, 469–476. [Google Scholar] [CrossRef]
- Rose, S.H.; van Zyl, W.H. Exploitation of Aspergillus niger for the heterologous production of cellulases and hemicellulases. Open Biotechnol. J. 2008, 2, 167–175. [Google Scholar] [CrossRef]
- Fleißner, A.; Dersch, P. Expression and export: Recombinant protein production systems for Aspergillus. Appl. Microbiol. Biot. 2010, 87, 1255–1270. [Google Scholar] [CrossRef]
- Sun, X.; Wu, H.; Zhao, G.; Li, Z.; Wu, X.; Liu, H.; Zheng, Z. Morphological regulation of Aspergillus niger to improve citric acid production by chsC gene silencing. Bioproc. Biosyst. Eng. 2018, 41, 1029–1038. [Google Scholar] [CrossRef]
- van Leeuwe, T.M.; Gerritsen, A.; Arentshorst, M.; Punt, P.J.; Ram, A.F.J. Rab GDP-dissociation inhibitor gdiA is an essential gene required for cell wall chitin deposition in Aspergillus niger. Fungal Genet. Biol. 2020, 136, 103319. [Google Scholar] [CrossRef]
- de Groot, P.W.J.; Brandt, B.W.; Horiuchi, H.; Ram, A.F.J.; de Koster, C.G.; Klis, F.M. Comprehensive genomic analysis of cell wall genes in Aspergillus nidulans. Fungal Genet. Biol. 2009, 46, S72–S81. [Google Scholar] [CrossRef]
- Gow, N.A.R.; Latge, J.P.; Munro, C.A. The fungal cell wall: Structure, biosynthesis, and function. Microbiol. Spectr. 2017, 5, 1–25. [Google Scholar] [CrossRef]
- Ruiz-Herrera, J.; Ortiz-Castellanos, L. Cell wall glucans of fungi. A review. Cell Surface 2019, 5, 100022. [Google Scholar] [CrossRef]
- Olaya-Abril, A.; Jiménez-Munguía, I.; Gómez-Gascón, L.; Rodríguez-Ortega, M.J. Surfomics: Shaving live organisms for a fast proteomic identification of surface proteins. J. Proteom. 2014, 97, 164–176. [Google Scholar] [CrossRef]
- Lichius, A.; Ruiz, D.M.; Zeilinger, S. Genetic Transformation of Filamentous Fungi: Achievements and Challenges. In Grand Challenges in Biology and Biotechnology; Nevalainen, H., Ed.; Springer: Cham, Switzerland, 2020; Volume Grand Challenges in Fungal Biotechnology, pp. 123–164. [Google Scholar] [CrossRef]
- Fang, T.H.; Kang, S.H.; Hong, Z.H.; Wu, C.D. Elasticity and nanomechanical response of Aspergillus niger spores using atomic force microscopy. Micron 2012, 43, 407–411. [Google Scholar] [CrossRef]
- Wargenau, A.; Fleißner, A.; Bolten, C.J.; Rohde, M.; Kampen, I.; Kwade, A. On the origin of the electrostatic surface potential of Aspergillus niger spores in acidic environments. Res. Microbiol. 2011, 162, 1011–1017. [Google Scholar] [CrossRef]
- Pihet, M.; Vandeputte, P.; Tronchin, G.; Renier, G.; Saulnier, P.; Georgeault, S.; Bouchara, J.P. Melanin is an essential component for the integrity of the cell wall of Aspergillus fumigatus conidia. BMC Microbiol. 2009, 9, 177. [Google Scholar] [CrossRef]
- Koshiyama, K.; Kodama, T.; Yano, T.; Fujikawa, S. Structural change in lipid bilayers and water penetration induced by shock waves: Molecular dynamics simulations. Biophys. J. 2006, 91, 2198–2205. [Google Scholar] [CrossRef]
- Kodama, T.; Hamblin, M.R.; Doukas, A.G. Cytoplasmic molecular delivery with shock waves: Importance of impulse. Biophys. J. 2000, 79, 1821–1832. [Google Scholar] [CrossRef]
- Kuraya, E.; Miyafuji, Y.; Takemoto, A.; Itoh, S. The effect of underwater shock waves on steam distillation of Alpinia zerumbet leaves. Trans. Mater. Res. Soc. Jpn. 2014, 39, 447–449. [Google Scholar] [CrossRef]
- López-Marín, L.M.; Millán-Chiu, B.E.; Castaño-González, K.; Aceves, C.; Fernández, F.; Varela-Echavarría, A.; Loske, A.M. Shock wave-induced damage and poration in eukaryotic cell membranes. J. Membr. Biol. 2017, 250, 41–52. [Google Scholar] [CrossRef]
- Liauw, C.M.; Slate, A.J.; Butler, J.A.; Wilson-Nieuwenhuis, J.S.; Deisenroth, T.; Preuss, A.; Whitehead, K.A. The effect of surface hydrophobicity on the attachment of fungal conidia to substrates of polyvinyl acetate and polyvinyl alcohol. J. Polym. Environ. 2020, 28, 1450–1464. [Google Scholar] [CrossRef]
- Arora, M.; Ohl, C.D.; Mørch, K.A. Cavitation inception on microparticles: A self-propelled particle accelerator. Phys. Rev. Lett. 2004, 92, 174501. [Google Scholar] [CrossRef] [PubMed]
- Newton Harvey, E.; Barnes, D.K.; McElroy, W.D.; Whiteley, A.H.; Pease, D.C.; Cooper, K.W. Bubble formation in animals. I. Physical factors. J. Cell. Comp. Physiol. 1944, 24, 1–22. [Google Scholar] [CrossRef]
- Newton Harvey, E.; McElroy, W.D.; Whiteley, A.H. On cavity formation in water. J. Appl. Phys. 1947, 18, 162–172. [Google Scholar] [CrossRef]
- Kfoury, R.; Marzban, B.; Makki, E.; Greenfield, M.L.; Yuan, H. Effect of pressure profile of shock waves on lipid membrane deformation. PLoS ONE 2019, 14, e0212566. [Google Scholar] [CrossRef] [PubMed]
- de Icaza-Herrera, M.; Fernández, F.; Loske, A.M. Combined short and long-delay tandem shock waves to improve shock wave lithotripsy according to the Gilmore–Akulichev theory. Ultrasonics 2015, 58, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Pellegrino, A.; Hallack, A.; Petrinic, N.; Jérusalem, A.; Cleveland, R.O. Response of single cells to shock waves and numerically optimized waveforms for cancer therapy. Biophys. J. 2018, 114, 1433–1439. [Google Scholar] [CrossRef]
- Kodama, T.; Tomita, Y. Cavitation bubble behavior and bubble–shock wave interaction near a gelatin surface as a study of in vivo bubble dynamics. Appl. Phys. B 2000, 70, 139–149. [Google Scholar] [CrossRef]
- Qin, P.; Han, T.; Alfred, C.; Xu, L. Mechanistic understanding the bioeffects of ultrasound-driven microbubbles to enhance macromolecule delivery. J. Control. Release 2018, 272, 169–181. [Google Scholar] [CrossRef] [PubMed]
- van Wamel, A.; Kooiman, K.; Harteveld, M.; Emmer, M.; ten Cate, F.J.; Versluis, M.; de Jong, N. Vibrating microbubbles poking individual cells: Drug transfer into cells via sonoporation. J. Control. Release 2006, 112, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Juan, T.; Guo, X.S.; Di, X.; Dong, Z. Microstreaming velocity field and shear stress created by an oscillating encapsulated microbubble near a cell membrane. Chin. Phys. B 2014, 23, 124302. [Google Scholar] [CrossRef]
- Le Gac, S.; Zwaan, E.; van den Berg, A.; Ohl, C.D. Sonoporation of suspension cells with a single cavitation bubble in a microfluidic confinement. Lab Chip 2007, 7, 1666–1672. [Google Scholar] [CrossRef]
- Marmottant, P.; Hilgenfeldt, S. Controlled vesicle deformation and lysis by single oscillating bubbles. Nature 2003, 423, 153–156. [Google Scholar] [CrossRef]
- Zhao, L.; Schaefer, D.; Marten, M.R. Assessment of elasticity and topography of Aspergillus nidulans spores via atomic force microscopy. Appl. Environ. Microb. 2005, 71, 955–960. [Google Scholar] [CrossRef]
- Hernáez, M.L.; Ximénez-Embún, P.; Martínez-Gomariz, M.; Gutiérrez-Blázquez, M.D.; Nombela, C.; Gil, C. Identification of Candida albicans exposed surface proteins in vivo by a rapid proteomic approach. J. Proteom. 2010, 73, 1404–1409. [Google Scholar] [CrossRef]
- Karkowska-Kuleta, J.; Satala, D.; Bochenska, O.; Rapala-Kozik, M.; Kozik, A. Moonlighting proteins are variably exposed at the cell surfaces of Candida glabrata, Candida parapsilosis and Candida tropicalis under certain growth conditions. BMC Microbiol. 2019, 19, 149. [Google Scholar] [CrossRef] [PubMed]
- Gow, N.A.R.; Lenardon, M.D. Architecture of the dynamic fungal cell wall. Nat. Rev. Microbiol. 2023, 21, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Gil-Bona, A.; Parra-Giraldo, C.M.; Hernáez, M.L.; Reales-Calderon, J.A.; Solis, N.V.; Filler, S.G.; Gil, C. Candida albicans cell shaving uncovers new proteins involved in cell wall integrity, yeast to hypha transition, stress response and host–pathogen interaction. J. Proteom. 2015, 127, 340–351. [Google Scholar] [CrossRef] [PubMed]
- Yin, Q.Y.; de Groot, P.W.; de Koster, C.G.; Klis, F.M. Mass spectrometry-based proteomics of fungal wall glycoproteins. Trends Microbiol. 2008, 16, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Y.; Hua, X.W.; Jia, H.R.; Li, C.; Lin, F.; Chen, Z.; Wu, F.G. Universal cell surface imaging for mammalian, fungal, and bacterial cells. ACS Biomater. Sci. Eng. 2016, 2, 987–997. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Mode | Tandem Events | Number of SWs 1 |
|---|---|---|---|
| Control | - | - | 0 |
| 1 | SP 2 | - | 50 |
| 2 | SP | - | 100 |
| 3 | SP | - | 200 |
| 4 | SDT 3 | 25 | 50 |
| 5 | SDT | 50 | 100 |
| 6 | SDT | 100 | 200 |
| 7 | LDT 4 | 25 | 50 |
| 8 | LDT | 50 | 100 |
| 9 | LDT | 100 | 200 |
| Sample | Mean Z Potential (mV) ± SD 1 | Electrophoretic Mobility (µm × cm/Vs) |
|---|---|---|
| Control without SWs 2 | −20.37 ± 0.94 | −1.44 |
| SP 3 50 SWs | −28.85 ± 0.82 | −2.04 |
| SP 100 SWs | −26.98 ± 0.59 | −1.91 |
| SP 200 SWs | −30.09 ± 0.74 | −2.13 |
| SDT 4 50 SWs | −33.34 ±0.35 | −2.36 |
| SDT 100 SWs | −29.56 ± 1.33 | −2.09 |
| SDT 200 SWs | −32.66 ± 0.81 | −2.31 |
| LDT 5 50 SWs | −27.3 ± 1.0 | −1.93 |
| LDT 100 SWs | −29.57 ± 1.22 | −2.09 |
| LDT 200 SWs | −30.36 ± 0.04 | −2.15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Maldonado, M.A.; Millán-Chiu, B.E.; Fernández, F.; Larrañaga, D.; Gómez-Lim, M.A.; Loske, A.M. Underwater Shock Wave-Enhanced Cavitation to Induce Morphological Changes and Cell Permeabilization in Microscopic Fungi. Fluids 2024, 9, 81. https://doi.org/10.3390/fluids9040081
Martínez-Maldonado MA, Millán-Chiu BE, Fernández F, Larrañaga D, Gómez-Lim MA, Loske AM. Underwater Shock Wave-Enhanced Cavitation to Induce Morphological Changes and Cell Permeabilization in Microscopic Fungi. Fluids. 2024; 9(4):81. https://doi.org/10.3390/fluids9040081
Chicago/Turabian StyleMartínez-Maldonado, Miguel A., Blanca E. Millán-Chiu, Francisco Fernández, Daniel Larrañaga, Miguel A. Gómez-Lim, and Achim M. Loske. 2024. "Underwater Shock Wave-Enhanced Cavitation to Induce Morphological Changes and Cell Permeabilization in Microscopic Fungi" Fluids 9, no. 4: 81. https://doi.org/10.3390/fluids9040081