

Short-Stranded Zein Fibers for Muscle Tissue Engineering in Alginate-Based Composite Hydrogels
 ,
, 
Abstract
:1. Introduction
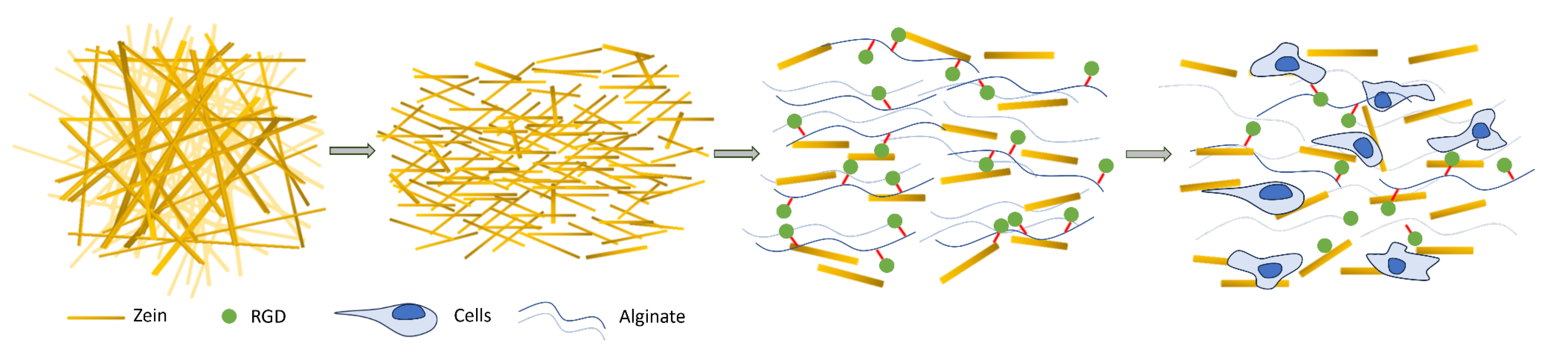
2. Results and Discussion
2.1. Electrospinning of Zein Fiber Solutions
2.2. Zein Fibers Can Be Processed into Cytocompatible Short Stranded Fibers
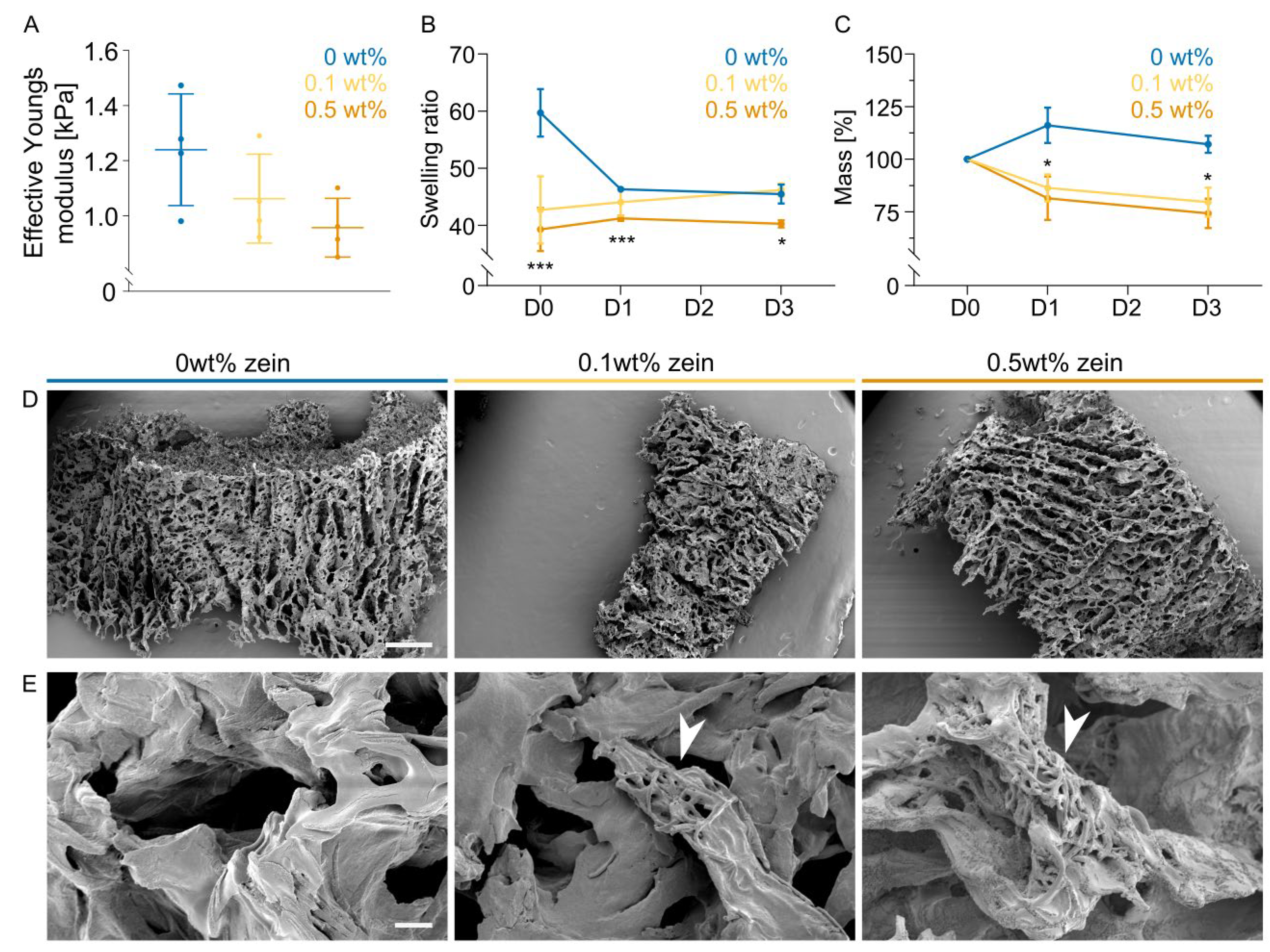
2.3. Zein Fiber Addition Leads to Faster Biomaterial Degradation
2.4. Addition of Zein Fibers Improves Compaction during Cultured Muscle Formation
2.5. Zein Fiber Addition Improves Protein Yield and Metabolic Activity
2.6. Discussion
3. Conclusions
4. Material and Methods
4.1. Preparation and Characterization of Spinning Solutions and Fibers
4.1.1. Solution Preparation
4.1.2. Viscosity Measurements
4.1.3. Conductivity Measurements
4.1.4. Electrospinning
4.1.5. Scanning Electron Microscopy
4.1.6. Ultrasonication
4.1.7. Fluorescently Labeled Zein
4.2. Hydrogel Formation and 3D Cell Culture
4.2.1. Cell Isolation and Purification
4.2.2. Synthesis of RGD-Functionalized Alginate
4.2.3. Gel Degradation Assays
4.2.4. Mechanical Properties
4.2.5. 3D Cell Culture
4.2.6. Protein Measurements
4.2.7. ELISAs
4.2.8. Western Blotting
4.2.9. Tissue Staining
4.2.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Post, M.J. Cultured meat from stem cells: Challenges and prospects. Meat Sci. 2012, 92, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Parodi, A.; Leip, A.; De Boer, I.J.M.; Slegers, P.M.; Ziegler, F.; Temme, E.H. The potential of future foods for sustainable and healthy diets. Nat. Sustain. 2018, 1, 782–789. [Google Scholar] [CrossRef]
- Post, M.J.; Levenberg, S.; Kaplan, D.L.; Genovese, N.; Fu, J.; Bryant, C.J. Scientific, sustainability and regulatory challenges of cultured meat. Nat. Food 2020, 1, 403–415. [Google Scholar] [CrossRef]
- Messmer, T.; Klevernic, I.; Furquim, C.; Ovchinnikova, E.; Dogan, A.; Cruz, H. A serum-free media formulation for cultured meat production supports bovine satellite cell differentiation in the absence of serum starvation. Nat. Food 2022, 3, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Ben-Arye, T.; Shandalov, Y.; Ben-Shaul, S.; Landau, S.; Zagury, Y.; Ianovici, I. Textured soy protein scaffolds enable the generation of three-dimensional bovine skeletal muscle tissue for cell-based meat. Nat. Food 2020, 1, 210–220. [Google Scholar] [CrossRef]
- Stout, A.J.; Mirliani, A.B.; Rittenberg, M.L.; Shub, M.; White, E.C.; Yuen, J.S., Jr.; Kaplan, D.L. Simple and effective serum-free medium for sustained expansion of bovine satellite cells for cell cultured meat. Commun. Biol. 2022, 5, 466. [Google Scholar] [CrossRef]
- Kolkmann, A.M.; Van Essen, A.; Post, M.J.; Moutsatsou, P. Development of a Chemically Defined Medium for in vitro Expansion of Primary Bovine Satellite Cells. Front. Bioeng. Biotechnol. 2022, 10, 895289. [Google Scholar] [CrossRef] [PubMed]
- Mitić, R.; Cantoni, F.; Börlin, C.S.; Post, M.J.; Jackisch, L. A simplified and defined serum-free medium for cultivating fat across species. iScience 2023, 26, 105822. [Google Scholar] [CrossRef] [PubMed]
- Spanier, A.M.; Flores, M.; Toldrá, F.; Aristoy, M.C.; Bett, K.L.; Bystricky, P.; Bland, J.M. Meat flavor: Contribution of proteins and peptides to the flavor of beef. Adv. Exp. Med. Biol. 2004, 542, 33–49. [Google Scholar]
- Zhu, Y.; Hamill, R.M.; Mullen, A.M.; Kelly, A.L.; Gagaoua, M. Molecular mechanisms contributing to the development of beef sensory texture and flavour traits and related biomarkers: Insights from early post-mortem muscle using label-free proteomics. J. Proteom. 2023, 286, 104953. [Google Scholar] [CrossRef]
- Das, M.; Rumsey, J.W.; Bhargava, N.; Stancescu, M.; Hickman, J.J. Skeletal muscle tissue engineering: A maturation model promoting long-term survival of myotubes, structural development of the excitation-contraction coupling apparatus and neonatal myosin heavy chain expression. Biomaterials 2009, 30, 5392–5402. [Google Scholar] [CrossRef] [PubMed]
- Bian, W.; Bursac, N. Engineered skeletal muscle tissue networks with controllable architecture. Biomaterials 2009, 30, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- Ostrovidov, S.; Ramalingam, M.; Bae, H.; Orive, G.; Fujie, T.; Shi, X.; Kaji, H. Latest developments in engineered skeletal muscle tissues for drug discovery and development. Expert Opin. Drug Discov. 2023, 18, 47–63. [Google Scholar] [CrossRef] [PubMed]
- Wollschlaeger, J.O.; Maatz, R.; Albrecht, F.B.; Klatt, A.; Heine, S.; Blaeser, A.; Kluger, P.J. Scaffolds for Cultured Meat on the Basis of Polysaccharide Hydrogels Enriched with Plant-Based Proteins. Gels 2022, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Rowley, J.A.; Madlambayan, G.; Mooney, D.J. Alginate hydrogels as synthetic extracellular matrix materials. Biomaterials 1999, 20, 45–53. [Google Scholar] [CrossRef]
- Handorf, A.M.; Zhou, Y.; Halanski, M.A.; Li, W.-J. Tissue stiffness dictates development, homeostasis, and disease progression. Organogenesis 2015, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Guo, S.S.; Fässler, R. Integrin-mediated mechanotransduction. J. Cell Biol. 2016, 215, 445–456. [Google Scholar] [CrossRef]
- Momany, F.A.; Sessa, D.J.; Lawton, J.W.; Selling, G.W.; Hamaker, S.A.H. Structural characterization of alpha-zein. J. Agric. Food Chem. 2006, 54, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Miao, Y.; Wang, Y.; Zhang, L.-M.; Lin, X. Electrospun Zein/Gelatin Scaffold-Enhanced Cell Attachment and Growth of Human Periodontal Ligament Stem Cells. Materials 2017, 10, 1168. [Google Scholar] [CrossRef]
- Yao, C.; Li, X.; Song, T. Electrospinning and crosslinking of zein nanofiber mats. Appl. Polym. Sci. 2007, 103, 380–385. [Google Scholar] [CrossRef]
- Khalesi, H.; Lu, W.; Nishinari, K.; Fang, Y. Fundamentals of composites containing fibrous materials and hydrogels: A review on design and development for food applications. Food Chem. 2021, 364, 130329. [Google Scholar] [CrossRef] [PubMed]
- Mao, W.; Lee, S.; Kim, S.-R.; Kim, K.-N.; Yoo, H.S. Electrospun nanohybrid hydrogels for enhanced differentiation of myoblasts. J. Ind. Eng. Chem. 2019, 80, 838–845. [Google Scholar] [CrossRef]
- Fernandez, P.; Bausch, A.R. The compaction of gels by cells: A case of collective mechanical activity. Integr. Biol. 2009, 1, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Lueckgen, A.; Garske, D.S.; Ellinghaus, A.; Mooney, D.J.; Duda, G.N.; Cipitria, A. Enzymatically-degradable alginate hydrogels promote cell spreading and in vivo tissue infiltration. Biomaterials 2019, 217, 217. [Google Scholar] [CrossRef] [PubMed]
- Ostrovidov, S.; Hosseini, V.; Ahadian, S.; Fujie, T.; Parthiban, S.P.; Ramalingam, M.; Bae, H.; Hirokazu, K.; Khademhosseini, A. Skeletal Muscle Tissue Engineering: Methods to Form Skeletal Myotubes and Their Applications. Tissue Eng. Part B Rev. 2014, 20, 403–436. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Su, J.; Brennan, C.S.; Van der Meeren, P.; Zhang, N.; Tong, Y.; Wang, P. Recent developments of electrospun zein nanofibres: Strategies, fabrication and therapeutic applications. Mater. Today Adv. 2022, 16, 100307. [Google Scholar] [CrossRef]
- Kalluri, L.; Satpathy, M.; Duan, Y. Effect of Electrospinning Parameters on the Fiber Diameter and Morphology of PLGA Nanofibers. Dent. Oral Biol. Craniofacial Res. 2021, 4. [Google Scholar] [CrossRef]
- Li, Y.; Li, J.; Xia, Q.; Zhang, B.; Wang, Q.; Huang, Q. Understanding the dissolution of α-zein in aqueous ethanol and acetic acid solutions. J. Phys. Chem. B 2012, 116, 12057–12064. [Google Scholar] [CrossRef]
- Selling, G.W.; Biswas, A.; Patel, A.; Walls, D.J.; Dunlap, C.; Wei, Y. Impact of Solvent on Electrospinning of Zein and Analysis of Resulting Fibers. Macromol. Chem. Phys. 2007, 208, 1002–1010. [Google Scholar] [CrossRef]
- Haider, A.; Haider, S.; Kang, I.K. A comprehensive review summarizing the effect of electrospinning parameters and potential applications of nanofibers in biomedical and biotechnology. Arab. J. Chem. 2018, 11, 1165–1188. [Google Scholar] [CrossRef]
- Meng, L.; Shao, C.; Yang, J. Ionically Cross-Linked Silk Microfibers/Alginate Tough Composite Hydrogels with Hierarchical Structures. ACS Sustain. Chem. Eng. 2018, 6, 16788–16796. [Google Scholar] [CrossRef]
- Jana, S.; Leung, M.; Chang, J.; Zhang, M. Effect of nano- and micro-scale topological features on alignment of muscle cells and commitment of myogenic differentiation. Biofabrication 2014, 6, 035012. [Google Scholar] [CrossRef] [PubMed]
- Yeo, M.; Kim, G. Nano/microscale topographically designed alginate/PCL scaffolds for inducing myoblast alignment and myogenic differentiation. Carbohydr. Polym. 2019, 223, 115041. [Google Scholar] [CrossRef] [PubMed]
- Formigli, L.; Meacci, E.; Sassoli, C.; Squecco, R.; Nosi, D.; Chellini, F. Cytoskeleton/stretch-activated ion channel interaction regulates myogenic differentiation of skeletal myoblasts. J. Cell. Physiol. 2007, 211, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Messmer, T.; Dohmen, R.G.; Schaeken, L.; Melzener, L.; Hueber, R.; Godec, M. Single-cell analysis of bovine muscle-derived cell types for cultured meat production. Front. Nutr. 2023, 10, 1212196. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | 20 wt% | 25 wt% | 28 wt% | 30 wt% | 35 wt% |
| Zein concentration [mg∙ml−1] | 200 | 250 | 280 | 300 | 350 |
| Viscosity [mPa.s] | 81 | 208 | 359 | 544 | 1708 |
| Conductivity [µS cm−1] | 281.2 | 258.2 | 240.0 | 226.8 | 195.6 |
| Taylor cone formation | yes | yes | yes | yes | no |
| Morphology | bead formation | mostly fibers w/some bead formation | homogeneous fibers | heterogeneous melted fibers | heterogeneous thick fibers |
| Fiber diameter [µm] | 0.130 | 0.365 | 0.664 | 0.980 | 1.815 |
| Fiber size distribution [µm] | ±0.029 | ±0.136 | ±0.202 | ±0.639 | ±0.889 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melzener, L.; Spaans, S.; Hauck, N.; Pötgens, A.J.G.; Flack, J.E.; Post, M.J.; Doğan, A. Short-Stranded Zein Fibers for Muscle Tissue Engineering in Alginate-Based Composite Hydrogels. Gels 2023, 9, 914. https://doi.org/10.3390/gels9110914
Melzener L, Spaans S, Hauck N, Pötgens AJG, Flack JE, Post MJ, Doğan A. Short-Stranded Zein Fibers for Muscle Tissue Engineering in Alginate-Based Composite Hydrogels. Gels. 2023; 9(11):914. https://doi.org/10.3390/gels9110914
Chicago/Turabian StyleMelzener, Lea, Sergio Spaans, Nicolas Hauck, André J. G. Pötgens, Joshua E. Flack, Mark J. Post, and Arın Doğan. 2023. "Short-Stranded Zein Fibers for Muscle Tissue Engineering in Alginate-Based Composite Hydrogels" Gels 9, no. 11: 914. https://doi.org/10.3390/gels9110914