
The Fungal, Nutritional, and Metabolomic Diagnostics of the Oil Palm Elaeis guineensis Affected by Bud Rot Disease in Esmeraldas, Ecuador

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Foliar Sample Collection
2.3. Soil Sample Collection
2.4. DRIS Analysis
2.5. Metagenomic Analysis
2.5.1. DNA Extraction
2.5.2. MiSeq Illumina Sequencing
2.6. Bioinformatics and Data Analysis
2.7. LC-MS
2.7.1. Extraction Process
2.7.2. LC-MS Analysis
3. Results
3.1. DRIS Analysis Based on the Soil and Foliar Determinations
3.2. Metagenomic Analysis
3.3. Liquid Chromatography-Mass Spectrometry (LC-MS) Analysis
4. Discussion
4.1. The Diagnosis and Recommendation Integrated System—DRIS
4.2. Metagenomic Analysis
4.3. LC-MS Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rozali, N.; Yarmo, M.; Abu Seman, I.; Kushairi, A.; Ramli, U. Metabolomics differentiation of oil palm (Elaeis guineensis Jacq.) spear leaf with contrasting susceptibility to Ganoderma boninense. Plant Omics J. 2017, 10, 45–52. [Google Scholar] [CrossRef]
- Naher, L.; Yusuf, U.K.; Ismail, A.; Tan, S.G.; Mondal, M.M.A. Ecological status of Ganoderma and basal stem rot disease of oil palms (Elaeis guineensis Jacq.). Aust. J. Crop Sci. 2013, 7, 1723–1727. [Google Scholar]
- Owoyele, B.V.; Owolabi, G.O. Traditional oil palm (Elaeis guineensis Jacq.) and its medicinal uses: A review. Cell Med. 2014, 4, e16. [Google Scholar] [CrossRef]
- Nair, P.K. Tree Crops. In Harvesting Cash from the World’s Important Cash Crops, 1st ed.; Springer Nature: Cham, Switzerland, 2021; pp. 249–285. [Google Scholar]
- Widiastuti, H.; Taniwiryono, D.; Hendarjanti, H.; Harjotedjo, S. Chemical and biological characteristics of selected wet soil in oil palm plantation attack with Ganoderma sp. IOP Conf. Ser. 2018, 183, 012019. [Google Scholar] [CrossRef]
- Chung, G.F. Effect of pests and diseases on oil palm yield. In Palm Oil; AOCS Press: Urbana, IL, USA, 2012; pp. 163–210. [Google Scholar]
- Aderungboye, F.O. Diseases of the oil palm. Int. J. Pest Manag. 1977, 23, 305–326. [Google Scholar] [CrossRef]
- Viera-Torres, M.; Sinde-González, I.; Gil-Docampo, M.; Bravo-Yandún, V.; Toulkeridis, T. Generating the Baseline in the Early Detection of Bud Rot and Red Ring Disease in Oil Palms by Geospatial Technologies. Remote Sens. 2020, 12, 3229. [Google Scholar] [CrossRef]
- Benítez, É.; García, C. The history of research on oil palm bud rot (Elaeis guineensis Jacq.) in Colombia. Agron. Colomb. 2014, 32, 390–398. [Google Scholar] [CrossRef]
- Sarria, G.A.; Torres, G.A.; Aya, H.A.; Ariza, J.G.; Rodríguez, J.; Vélez, D.C.; Varón, F.; Martínez, G. Phytophthora sp. es el responsable de las lesiones iniciales de la pudrición de cogollo (PC) de la palma de aceite en Colombia. Palmas 2008, 29, 31–41. [Google Scholar]
- Franqueville, H. Oil palm Bud Rot in latín America. Exp. Agric. 2003, 39, 225–240. [Google Scholar] [CrossRef]
- Available online: https://oec.world/en/profile/bilateral-product/palm-oil/reporter/ecu?redirect=true (accessed on 15 January 2023).
- Instituto Nacional de Estadística y Censos [INEC]. Manual del Encuestador y Supervisor “Encuesta de Superficie y Producción Agropecuaria Continua ESPAC” 2019; Instituto Nacional de Estadistica y Censos: Quito, Ecuador, 2019.
- Allen, W.; Paulus, A.; van Gundy, S.; Swinburne, T.R.; Ollagnier, M.; Renard, J.L. Report on the mission to Ecuador. In Document Cirad-Cp; Palmeras del Ecuador-Palmoriente: Montpellier, France, 1995. [Google Scholar]
- Gómez, P.; Ayala, L.; Munévar, F. Characteristics and management of bud rot, a disease of oil palm. In Plantation Tree Crops in the New Millennium: The Way Ahead; Pushparajah, E., Ed.; The Incorporated Society of Planters: Kuala Lumpur, Malaysia, 2000; Volume 1, pp. 545–553. [Google Scholar]
- Albertazzi, H.; Bulgarelli, J.; Chinchilla, C. Onset of spear root symptoms in oil palm and prior (and contemporary) events. ASD Oil Palm Pap. 2005, 21, e41. [Google Scholar]
- Gunasinghe, N.; Mingpei, P.Y.; Cawthray, G.R.; Barbetti, M.J. Cercosporin from Pseudocercosporella capsellae and its critical role in white leaf spot deveelopment. Plant Dis. 2016, 100, 1521–1531. [Google Scholar] [CrossRef] [PubMed]
- Avendaño, J.C.; Garzon, G.M. Experiencias del manejo de la pudrición del cogollo (PC) en Elaeis guineensis y el híbrido OxG en Astorga S.A. Palmas 2013, 34, 77–88. [Google Scholar]
- Aya, H.; Varón, F.; Martínez, G.; Cayón, G. Manejo químico de la Pudrición de cogollo en cuatro materiales de palma de aceite en Tumaco. Rev. Palmas-Colomb. 2011, 32, 45–52. [Google Scholar]
- Torres, G.A.; Sarria, G.A.; Martínez, G. Identificación Temprana y Manejo de la Pudrición del Cogollo de la Palma de Aceite, 1st ed.; Cenipalma-Fedepalma-Sena SAC: Bogotá, Colombia, 2010. [Google Scholar]
- Bennett, A.J.; Bending, G.D.; Chandler, D.; Hilton, S.; Mills, P. Meeting the demand for crop production: The challenge of yield decline in crops grown in short rotations. Biol. Rev. 2012, 87, 52–71. [Google Scholar] [CrossRef] [PubMed]
- Kirkman, E.R.; Hilton, S.; Sethuraman, G.; Elias, D.M.O.; Taylor, A.; Clarkson, J.; Soh, A.C.; Bass, D.; Ooi, G.T.; McNamara, N.P.; et al. Diversity and Ecological Guild Analysis of the Oil Palm Fungal Microbiome Across Root, Rhizosphere, and Soil Compartments. Front. Microbiol. 2022, 13, 792928. [Google Scholar] [CrossRef] [PubMed]
- Kazeeroni, E.A.; Al-Sadi, A.M. 454-pyrosequencing reveals variable fungal diversity across farming systems. Front. Plant Sci. 2016, 7, 314. [Google Scholar] [CrossRef] [PubMed]
- Pagare, S.; Manila, B.; Niraj, T.; Pagare, S.; Bansal, Y.K. Secondary Metabolites of Plants and their Role: Overview. Curr. Trends Biotechnol. Pharm. 2015, 9, 293–304. [Google Scholar]
- Ramírez-Orellana, A.; Ruiz-Palomo, D.; Rojo-Ramírez, A.; Burgos-Burgos, J.E. The Ecuadorian Banana Farms Managers’ Perceptions: Innovation as a Driver of Environmental Sustainability Practices. Agriculture 2021, 11, 213. [Google Scholar] [CrossRef]
- Ricaurte, D.V. Inventario y Georeferenciación de los Principales Riesgos en la Ciudad de Quinindé—Provincia de Esmeraldas; IAEN: Esmeraldas, Ecuador, 2011; p. 73. [Google Scholar]
- Direction de Planification GADPE Prefectura Esmeraldas—Plan de Desarrollo Y Ordenamiento Territorial 2015–2025. Available online: https://www.prefecturadeesmeraldas.gob.ec/web/assets/2017--pdot-gadpe-septiembre.pdf (accessed on 23 February 2023).
- Gobierno Autónomo Descentralizado de la Provincia de Esmeraldas [GADPE]. 2012. Available online: https://prefecturadeesmeraldas.gob.ec/ (accessed on 23 February 2023).
- Marrocos, P.C.L.; Loureiro, G.A.H.D.A.; De Araujo, Q.R.; Sodré, G.A.; Ahnert, D.; Baligar, V.C. Mineral nutrient ratios and cacao productivity. J. Plant Nutr. 2020, 43, 2368–2382. [Google Scholar] [CrossRef]
- Gieseking, J.E.; Snider, H.J.; Getz, C.A. Destruction of organic matter in plant material by the use of nitric and perchloric acids. Ind. Eng. Chem. Anal. 1935, 7, 185–186. [Google Scholar] [CrossRef]
- Gee, A.; Domingues, L.; Dietz, V. Constituents in sugar. Anal. Chem. 1954, 26, 1487–1492. [Google Scholar] [CrossRef]
- Matos, G.S.B.; Fernandes, A.R.; Wadt, P.G.S.; Pina, A.J.A.; Franzini, V.I.; Ramos, H.M.N. The use of DRIS for nutritional diagnosis in oil palm in the state of Pará. Rev. Bras. Cien. Solo 2017, 41, e0150466. [Google Scholar] [CrossRef]
- de la Torre, A.; Iglesias, I.; Carballo, M.; Ramírez, P.; Muñoz, M.J. An approach for mapping the vulnerability of European Union soils to antibiotic contamination. Sci. Total Environ. 2012, 414, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Beaufils, E.R. Diagnosis and recommendation integrated system (DRIS). Soil Sci. Bull. Univ. Natal. 1973, 1, 1–132. [Google Scholar]
- Badotti, F.; De Oliveira, F.S.; Garcia, C.F.; Vaz, A.B.M.; Fonseca, P.L.C.; Nahum, L.A.; Oliveira, G.; Góes-Neto, A. Effectiveness of ITS and sub-regions as DNA barcode markers for the identification of Basidiomycota (Fungi). BMC Microbiol. 2017, 17, 42. [Google Scholar] [CrossRef] [PubMed]
- Siddique, A.B.; Albrectsen, B.R.; Ilbi, H.; Siddique, A.B. Optimization of Protocol for Construction of Fungal ITS Amplicon Library for High-Throughput Illumina Sequencing to Study the Mycobiome of Aspen Leaves. Appl. Sci. 2022, 12, 1136. [Google Scholar] [CrossRef]
- Al-Bulushi, I.M.; Bani-Uraba, M.S.; Guizani, N.S.; Al-Khusaibi, M.K.; Al-Sadi, A.M. Illumina MiSeq sequencing analysis of fungal diversity in stored dates. BMC Microbiol. 2017, 17, 72. [Google Scholar] [CrossRef]
- Hoggard, M.; Vesty, A.; Wong, G.; Montgomery, J.M.; Fourie, C.; Douglas, R.G.; Biswas, K.; Taylor, M.W. Characterizing the Human Mycobiota: A Comparison of Small Subunit rRNA, ITS1, ITS2, and Large Subunit rRNA Genomic Targets. Front. Microbiol. 2018, 9, 2208. [Google Scholar] [CrossRef]
- Al-Sadi, A.M.; Kazerooni, E.A. Illumina-MiSeq analysis of fungi in acid lime roots reveals dominance of Fusarium and variation in fungal taxa. Sci. Rep. 2018, 8, 17388. [Google Scholar] [CrossRef]
- Bolger, A.; Lohse, M.; Usadel, B. Trimmomatic: A flexible read trimming tool for Illumina Sequence. Bioinformatics 2014, 30, 2114–2120. Available online: http://www.usadellab.org/cms/?page=trimmomatic (accessed on 23 February 2023). [CrossRef]
- Fan, S.; Miao, L.; Li, H.; Lin, A.; Song, F.; Zhang, P. Illumina-based analysis yields new insights into the diversity and composition of endophytic fungi in cultivated Huperzia serrata. PLoS ONE 2020, 15, e0242258. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://unite.ut.ee/ (accessed on 15 February 2023).
- Hashizume, H.; Taga, S.; Sakata, M.K.; Taha, M.H.M.; Siddig, E.E.; Minamoto, T.; Fahal, A.H.; Kaneko, S. Detection of multiple mycetoma pathogens using fungal metabarcoding analysis of soil DNA in an endemic area of Sudan. PLoS Negl. Trop. Dis. 2022, 16, e0010274. [Google Scholar] [CrossRef]
- Irakli, M.; Skendi, A.; Bouloumpasi, E.; Chatzopoulou, P.; Biliaderis, C.G. LC-MS Identification and Quantification of Phenolic Compounds in Solid Residues from the Essential Oil Industry. Antioxidants 2021, 10, 2016. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Price, W.E.; Ashton, J.; Tapsell, L.C.; Johnson, S. Identification and characterization of phenolic compounds in hydromethanolic extracts of. Food Chem. 2016, 211, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Bikoro Bi Athomo, A.; Engozogho Anris, S.P.; Safou Tchiama, R.; Eyma, F.; Arnaudguilhem, C.; Charrier, B. Identification of phenolic compounds from K. Ivorensis by selected chromatographic and spectrometric techniques. J. Renew. Mat. 2021, 9, 35–48. [Google Scholar] [CrossRef]
- OECD. Nutrient Balance (Indicator). 2023. Available online: https://doi.org/10.1787/82add6a9-en (accessed on 8 June 2023).
- Wadt, P.G.S.; Dias, J.R.M. Premises for DRIS application in forest species and palms. In Nutrition and Fertilization of Forest Species and Palms; Prado, R.M., Wadt, R.M.P.G.S., Eds.; FUNEP: Jaboticabal, Brazil, 2014; pp. 277–298. [Google Scholar]
- Thor, K. Calcium—Nutrient and Messenger. Front. Plant Sci. 2019, 10, 440. [Google Scholar] [CrossRef] [PubMed]
- Santos-Sánchez, N.F.; Salas-Coronado, R.; Hernández-Carlos, B.; Villanueva-Cañongo, C. Shikimic Acid Pathway in Biosynthesis of Phenolic Compounds. In Plant Physiological Aspects of Phenolic Compounds; IntechOpen: London, UK, 2019. [Google Scholar]
- Connolly, E.L.; Guerinot, M.L. Iron stress in plants. Genome Biol. 2002, 3, 1–4. [Google Scholar] [CrossRef]
- Viégas, I.J.M.; Furlan, J.J.; Frazão, D.A.C.; Batista, M.M.F. Influência do micronutriente ferro naocorrência do amarelecimento fatal do dendezeiro. Tech. Bull. 2000, 32–37. [Google Scholar]
- Abad, V. Análisis Etiológico de la Enfermedad Pudrición del Cogollo (PC) en Cultivo de Palma Aceitera (Elaeis guineensis Jacq.) en la Provincia de Esmeraldas, Ecuador; Universidad de Barcelona: Barcelona, Spain, 2022; Available online: https://www.tdx.cat/bitstream/handle/10803/675558/VMAV_TESIS.pdf?sequence=1&isAllowed=y (accessed on 13 April 2023).
- Orgiazzi, A.; Lumini, E.; Nilsson, R.H.; Girlanda, M.; Vizzini, A. Unravelling soil fungal communities from different Mediterranean land-use backgrounds. PLoS ONE 2012, 7, e34847. [Google Scholar] [CrossRef]
- Ramírez, M.; Benítez, E. Pudrición del Cogollo, la terrible enfermedad que ataca la palma de aceite. Crop Life Latinoamérica 2013. Available online: https://www.croplifela.org/es/plagas/listado-de-plagas/pudricion-del-cogollo (accessed on 13 April 2023).
- Sánchez-Potes, A. Enfermedades de la palma de aceite en América Latina. Rev. Palmas 1990, 11, 990. [Google Scholar]
- Terrero, P.; Peñaherrera, S.; Vera, D.; Navarrete, B.; Herrera, M. Prospección De Enfermedades Fúngicas En Plantaciones De Palma Aceitera En La Provincia De Los Ríos. Ecuad. Es Calid. 2017, 4, 21–25. [Google Scholar] [CrossRef]
- de Assis Costa, O.Y.; Tupinambá, D.D.; Bergmann, J.C.; Barreto, C.C.; Quirino, B.F. Correction: Fungal diversity in oil palm leaves showing symptoms of Fatal Yellowing disease. PLoS ONE 2021, 16, e0254042. [Google Scholar] [CrossRef] [PubMed]
- Castillo, S.R.M.; Miller, S.; Stewart, J. Colletotrichum spp. and other fungi associated with anthracnose on Coffea arabica L. in Mérida State, Venezuela. Summa Phytopathol. 2022, 48, 99–111. [Google Scholar] [CrossRef]
- Cao, X.; Xu, X.; Che, H.; West, J.S.; Luo, D. Eight Colletotrichum species, including a novel species, are associated with areca palm anthracnose in Hainan, China. Plant Dis. 2020, 104, 1369–1377. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.; Muharam, F.M.; Rashed, O.; Khairulmazmi, A. Soil fungal composition and diversity in oil palm plantation at Sungai ASAP, Sarawak, Malaysia. J. Oil Palm Res. 2021, 33, 215–226. [Google Scholar] [CrossRef]
- Moore-Landecker, E. Fundamentals of the Fungi; Prentice-Hall, Inc.: Englewood Cliffs, NJ, USA, 1972; pp. 80–100. [Google Scholar]
- Schmidt, O.; Bahmani, M.; Koch, G.; Potsch, T.; Brandt, K. Study of the fungal decay of oil palm wood using TEM and UV techniques. Int. Biodeterior. Biodegrad. 2016, 111, 37–44. [Google Scholar] [CrossRef]
- Rahman, R.; Ahmad Sofi, J.; Javeed, I.; Hussain Malik, T.; Nisar, S. Role of Micronutrients in Crop Production. Int. J. Curr. Microbiol. App Sci. 2020, 11, 2265–2287. [Google Scholar]
- Reglinski, T.; Walters, D. Induced Resistance for Plant Disease Control. In Disease Control in Crops: Biological and Environmentally-Friendly Approaches; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar] [CrossRef]
- Koziara, W.; Sulewska, H.; Panasiewicz, K. Effect of resistance stimulator application to some agricultural crops. J. Res. Appl. Agric. Eng. 2006, 51, 82–86. [Google Scholar]
- Jamiołkowska, A. Natural compounds as elicitors of plant resistance against diseases and new biocontrol strategies. Agronomy 2020, 10, 173. [Google Scholar] [CrossRef]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef]
- Nabil-Adam, A.; Elnosary, M.E.; Ashour, M.L.; El-Moneam, N.M.A.; Shreadah, M.A. Flavonoids Biosynthesis in Plants as a Defense Mechanism: Role and Function Concerning Pharmacodynamics and Pharmacokinetic Properties. In Flavonoid Metabolism—Recent Advances and Applications in Crop Breeding); Abbas, H.M.K., Ahmad, A., Eds.; IntechOpen: London, UK, 2023; p. 3. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, J.; Xia, R.; Li, D.; Wang, F. Antioxidant processes involving epicatechin decreased symptoms of pine wilt disease. Front. Plant Sci. 2022, 13, 1015970. Available online: https://www.frontiersin.org/articles/10.3389/fpls.2022.1015970 (accessed on 13 April 2023). [CrossRef] [PubMed]
- Chen, Z.; Liang, J.; Zhang, C.; Rodrigues, C.J. Epicatechin and catechin may prevent coffee berry disease by inhibition of appressorial melanization of Colletotrichum kahawae. Biotechnol. Lett. 2006, 28, 1637–1640. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Xu, X.; Li, Y.; Wang, Y.; Li, M.; Wang, Y.; Ding, X.; Chu, Z. Rutin-Mediated Priming of Plant Resistance to Three Bacterial Pathogens Initiating the Early SA Signal Pathway. PLoS ONE 2016, 11, e0146910. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Morishita, T.; Kim, S.-J.; Park, S.-U.; Woo, S.; Noda, T.; Takigawa, S. Physiological Roles of Rutin in the Buckwheat Plant. Jpn. Agric. Res. Q. 2015, 49, 37–43. [Google Scholar] [CrossRef]
- Chen, W.; Xiao, Z.; Wang, Y.; Wang, J.; Zhai, R.; Lin-Wang, K.; Espley, R.; Ma, F.; Li, P. Competition between anthocyanin and kaempferol glycosides biosynthesis affects pollen tube growth and seed set of Malus. Hort. Res. 2021, 8, 173. [Google Scholar] [CrossRef] [PubMed]
- Cid-Ortega, S.; Monroy-Rivera, J.A. Extraction of Kaempferol and Its Glycosides Using Supercritical Fluids from Plant Sources: A Review. Food Tech. Biotechnol. 2018, 56, 480–493. [Google Scholar] [CrossRef] [PubMed]
- De Melo, G.O.; Malvar, D.d.C.; Vanderlinde, F.A.; Rocha, F.F.; Pires, P.A.; Costa, E.A.; de Matos, L.G.; Kaiser, C.R.; Costa, S.S. Antinociceptive and anti-inflammatory kaempferol glycosides from Sedum dendroideum. J. Ethnopharmacol. 2009, 124, 228–232. [Google Scholar] [CrossRef]
- Yildiztugay, E.; Ozfidan-Konakci, C.; Kucukoduk, M.; Turkan, I. Flavonoid Naringenin Alleviates Short-Term Osmotic and Salinity Stresses Through Regulating Photosynthetic Machinery and Chloroplastic Antioxidant Metabolism in Phaseolus vulgaris. Front. Plant Sci. 2020, 11, 682. [Google Scholar] [CrossRef]
- An, J.; Kim, S.H.; Bahk, S.; Vuong, U.T.; Nguyen, N.T.; Do, H.L.; Kim, S.H.; Chung, W.S. Naringenin Induces Pathogen Resistance against Pseudomonas syringae through the Activation of NPR1 in Arabidopsis. Front. Plant Sci. 2021, 12, 672552. [Google Scholar] [CrossRef]
- Girish, T.K.; Kumar, K.A.; Prasada Rao, U.J.S. C-Glycosylated flavonoids from black gram husk: Protection against DNA and erythrocytes from oxidative damage and their cytotoxic effect on HeLa cells. Toxicol. Rep. 2016, 3, 652–663. [Google Scholar] [CrossRef]
- Dai, L.; Hu, Y.; Chen, C.-C.; Ma, L.; Guo, R.-T. Flavonoid C-Glycosyltransferases: Function, Evolutionary Relationship, Catalytic Mechanism and Protein Engineering. Chem. Bio Eng. Rev. 2021, 8, 15–26. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Potential Response to the Application of Nutrients (RPAN) | Inferior Limit (α) | Superior Limit |
|---|---|---|
| Cryodeficiency | −∞ | − |
| Prone to Deficiency | − | − |
| Nutritional Balance | − | |
| Prone to Excess or Toxicity | ||
| Excess or Toxicity | ∞ |
| Primer Sequence for ITS1 Region | |
|---|---|
| Forward | ITS_fwd_1 CTTGGTCATTTAGAGGAAGTAA |
| ITS_fwd_2 CTCGGTCATTTAGAGGAAGTAA | |
| ITS_fwd_3 CTTGGTCATTTAGAGGAACTAA | |
| ITS_fwd_4 CCCGGTCATTTAGAGGAAGTAA | |
| ITS_fwd_5 CTAGGCTATTTAGAGGAAGTAA | |
| Reverse | ITS_rev_1 GCTGCGTTCTTCATCGATGC |
| ITS_rev_2 GCTGCGTTCTTCATCGATGG | |
| ITS_rev_3 GCTACGTTCTTCATCGATGC | |
| ITS_rev_4 GCTGCGTTCTTCATCGATGT | |
| ITS_rev_5 ACTGTGTTCTTCATCGATGT | |
| Overhang Adapter Sequences | |
|---|---|
| Forward | TCGTCGGCAGCGTCAGATGTGTATAAGAGACAG-CTTGGTCATTTAGAGGAAGTAA |
| Reverse | GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAG-GCTGCGTTCTTCATCGATGC |
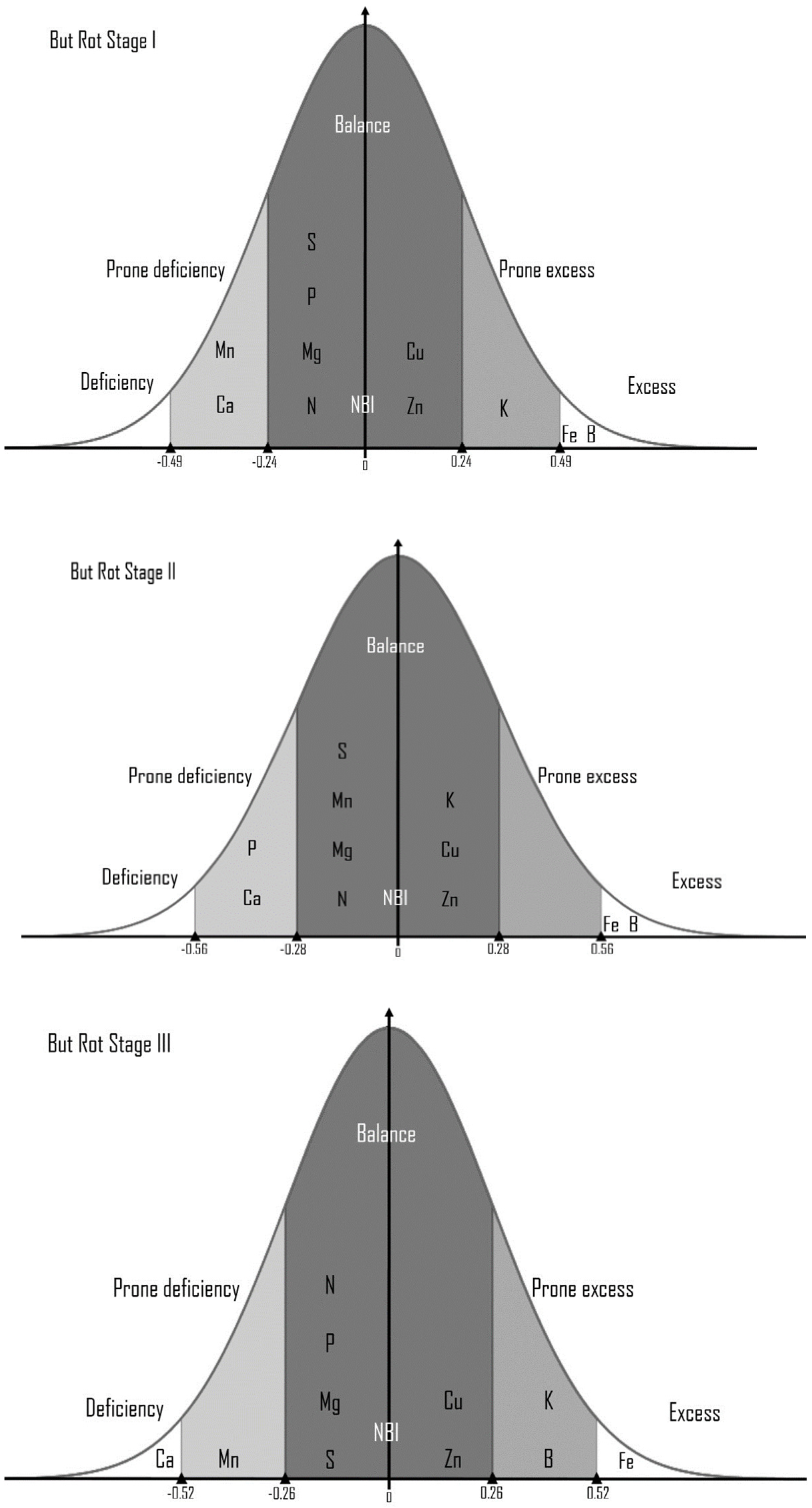
| DRIS Index/IBN | Bud Rot Disease Infection Stages in Oil Palm | |||||||
|---|---|---|---|---|---|---|---|---|
| Healthy Plants | Stage I | Stage II | Stage III | |||||
| Indices | RPANs | Indices | RPANs | Indices | RPANs | Indices | RPANs | |
| IN | −396.74 | −0.0 | −411.27 | −0.02 | −517.24 | −0.03 | −500.30 | −0.06 |
| IP | −622.36 | −0.44 | −468.81 | −0.15 | −513.36 | −0.02 | −567.44 | −0.18 |
| IK | 347.43 | 0.14 | 571.37 | 0.35 | 1012.20 | 0.70 | 774.99 | 0.50 |
| Ica | −266.58 | −0.41 | −274.21 | −0.39 | −360.95 | −0.33 | −246.98 | −0.65 |
| IMg | −343.01 | −0.15 | −327.64 | −0.21 | −345.97 | −0.37 | −411.53 | −0.14 |
| IS | −414.41 | −0.04 | −350.33 | −0.14 | −391.10 | −0.25 | −430.52 | −0.09 |
| IB | 175.26 | 0.82 | 204.91 | 0.68 | 271.70 | 0.61 | 295.41 | 0.47 |
| ICu | 451.92 | 0.12 | 367.41 | 0.09 | 349.42 | 0.36 | 432.07 | 0.09 |
| IZn | 513.27 | 0.25 | 442.28 | −0.26 | 496.37 | 0.01 | 380.87 | 0.22 |
| IMn | −365.38 | −0.09 | −524.50 | 0.65 | −872.40 | −0.56 | −622.09 | −0.28 |
| Ife | 865.11 | 0.77 | 773.28 | 921.22 | 0.61 | 897.74 | 0.64 | |
| IBN | 432.86 | 428.73 | 550.18 | 505.45 | ||||
| SD | 490.50 | 476.06 | 633.27 | 565.73 | ||||
| Category | Abundance % | |||
|---|---|---|---|---|
| A Infected Soil | B Healthy Soil | C Infected Plant | D Healthy Plant | |
| Unclassified at the species level | 25.8 | 30.89 | 31.03 | 19.78 |
| Ascochyta rabiei (A) | 20.79 | 9.3 | ||
| Talaromyces ruber (A) | 9.76 | 7.71 | ||
| Cryptococcus neoformans (B) | 6.58 | 5.5 | ||
| Antrodia sp. (B) | 3.43 | 3.25 | ||
| Saccharomyces sp. (A) | 3.37 | 7.9 | ||
| Candida sake (A) | 3.06 | |||
| Pyrenochaetopsis leptospora (A) | 1.98 | |||
| Wallemia sebi (B) | 2.37 | |||
| Acanthocorticium brueggemannii (B) | 1.99 | |||
| unidentified Hypocreales fam Incertae sedis sp. | 22.91 | 5.47 | ||
| Colletotrichum clidemiae (A) | 17.41 | |||
| Plectosphaerella cucumerina (A) | 12.93 | 2.33 | ||
| Acremonium stromaticum (A) | 5.49 | |||
| Fusarium solani (A) | 2.1 | 11.54 | ||
| Fusarium neocosmosporiellum (A) | 1.99 | 1043 | ||
| Plectosphaerella oratosquillae (A) | 0.8 | |||
| Cryptococcus nanyangensis (B) | 7.57 | |||
| Xenoacremonium recifei (A) | 4.21 | |||
| Fusarium ramigenum (A) | 3.88 | |||
| Wallemiales (B) | 0.74 | |||
| Myrmecridiales (A) | 0.4 | |||
| Magnaporthales (A) | 0.28 | |||
| Capnodiales (A) | 2.53 | |||
| Identified Compound | Molecular Formula | Ion Adduct | Molecular Weight (g/mol) | LC-MS | ||
|---|---|---|---|---|---|---|
| [M-H]− | rt | Fr | ||||
| Shikimic acid | C7H10O5 | M − H | 174.15 | 173.045 | 1.191 | |
| Epicatechin | C15H14O6 | M − H | 290.07904 | 289.072 | 1.609 | |
| (10E,15E)-9,12,13-trihydroxyoctadeca-10,15-dienoic acid | C18H32O5 | M − H | 328.4 | 327.218 | 21.467 | 3 |
| [(4E)-7-acetyloxy-6-hydroxy-2-methyl-10-oxo-2,3,6,7,8,9-hexahydrooxecin-3-yl] (E)-but-2-enoate | C16H22O7 | M + H | 326.34 | 325.129 | 31.257 | |
| [5-acetyloxy-3-(hydroxymethyl)-2-oxo-6-propan-2-ylcyclohex-3-en-1-yl] 3-methyl pentanoate | C18H28O6 | M + H | 340.4 | 339.181 | 32.56 | |
| 1-[2-methyl-6-[(2S,3R,4S,5S,6R)-3,4,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxyphenyl]ethanone | C14H18O7 | M + H | 298.29 | 311.114 | 32.046 | 21, 2, 12, 17, 5, 2 |
| 8-hydroxy-2,7,7,11,15-pentamethyl-5,12,16-trioxapentacyclo[9.8.0.0(2),.0,.0(1)(3),(1)]nonadec-13(18)-ene-3,17-dione | C21H28O6 | M − H | 376.4 | 377.102 | 1.14 | |
| alpha, alpha-Trehalose | C12H22O11 | M − H | 342.297 | 387.115 | 1.14 | 15 |
| Carnosine | C9H14N4O3 | M + H | 226.23 | 225.099 | 41.607 | 2, 3 |
| Cystine | C6H12N2O4S2 | M + H | 240.3 | 239.017 | 41.913 | 9, 1 |
| DOCOSANOL | C22H46O | M + H | 326.6 | 325.348 | 34.004 | |
| Ethylenediaminetetraacetic acid | C10H16N2O8 | M − H | 292.24 | 291.084 | 31.676 | 3 |
| IS_N-BENZOYL-D5-GLYCINE | C9H9NO3 | M − H | 184.2 | 183.082 | 1.191 | 4 |
| Canrenone | C22H28O3 | M − H | 340.2038 | 339.197 | 28.852 | 7, 3 |
| Isoorientin | C21H20O11 | M − H | 448.38 | 447.093 | 13.429 | 3, 3 |
| Massbank:IA000081 9-HODE | C18H32O3 | M − H1 | 296.235 | 295.228 | 32.232 | 7 |
| Massbank:IA000367 9-HOTrE | C18H30O3 | M − H1 | 294.219 | 293.212 | 30.656 | 3 |
| Citric acid | C6H8O7 | M + H | 192.12 | 191.02 | 1.191 | 2 |
| n-Capric acid | C10H20O2 | M − H | 172.146 | 171 | 1.123 | |
| Furosemide | C12H11ClN2O5S | M − H | 330.0077 | 329 | 30.21 | 2 |
| 2-Hydroxyhippuric acid|2-hydroxyhippurate | C9H9NO4 | M − H | 195.05316 | 194.1 | 28.272 | |
| Kaempferol-7-O-neohesperidoside | C27H30O15 | M − H | 594.15847 | 593.151 | 1.566 | 2 |
| Naringenin-7-O-glucoside | C21H22O10 | M − H | 434.397 | 433.114 | 32.002 | |
| Isovitexin | C21H20O10 | M − H | 432.381 | 431.098 | 1.744 | 1 |
| Caffeyl alcohol | C9H10O3 | M − H | 166.176 | 165.056 | 1.158 | |
| Massbank:PR309095 FA 18:2 + 2O | C18H32O4 | M − H | 312.45 | 311.222 | 29.94 | |
| Massbank:PR309165 MGMG 18:3 | C27H46O9 | M + HCOO | 514.656 | 559.312 | 31.229 | 3 |
| Massbank:PR309165 MGMG 18:3 | C27H46O9 | M + HCOO | 514.656 | 559.312 | 31.658 | 1 |
| Massbank:PR309171 DGMG 18:3 | C33H56O14 | M + HCOO | 676.8 | 721.363 | 29.236 | 3, 6 |
| Coumaroyl + C6H9O8 (isomer of 844, 845, 846) | C15H16O10 | M − H | 356.28 | 355.065 | 1.557 | 8 |
| Massbank:UT000256 9-HPODE | C18H32O4 | M − H | 312.23006 | 311.223 | 29.191 | 5 |
| Massbank:UT000264 9-HpOTrE | C18H30O4 | M − H | 310.21441 | 309.207 | 28.062 | 1 |
| Dodecylbenzenesulfonic acid | C18H30O3S | M-H | 326.19157 | 325.184 | 29.94 | 8 |
| Rutin | C27H30O16 | M − H | 610.15338 | 609.146 | 1.557 | |
| Sesamin | C20H18O6 | M + H | 354.4 | 353.103 | 33.906 | 14 |
| Sucrose | C12H22O11 | M − H | 342.3 | 341.109 | 1.14 | |
| Thymol-beta-D-glucoside | C16H24O6 | M + H | 312.36 | 311.15 | 34.672 | |
| Trihydroxy flavone-C-hexoside-C-pentoside | C27H30O15 | M − H | 594.5 | 563.141 | 1.557 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mihai, R.A.; Melo Heras, E.J.; Landazuri Abarca, P.A.; Catana, R.D. The Fungal, Nutritional, and Metabolomic Diagnostics of the Oil Palm Elaeis guineensis Affected by Bud Rot Disease in Esmeraldas, Ecuador. J. Fungi 2023, 9, 952. https://doi.org/10.3390/jof9090952
Mihai RA, Melo Heras EJ, Landazuri Abarca PA, Catana RD. The Fungal, Nutritional, and Metabolomic Diagnostics of the Oil Palm Elaeis guineensis Affected by Bud Rot Disease in Esmeraldas, Ecuador. Journal of Fungi. 2023; 9(9):952. https://doi.org/10.3390/jof9090952
Chicago/Turabian StyleMihai, Raluca A., Erly J. Melo Heras, Pablo A. Landazuri Abarca, and Rodica D. Catana. 2023. "The Fungal, Nutritional, and Metabolomic Diagnostics of the Oil Palm Elaeis guineensis Affected by Bud Rot Disease in Esmeraldas, Ecuador" Journal of Fungi 9, no. 9: 952. https://doi.org/10.3390/jof9090952