
Overexpression of the Capebp2 Gene Encoding the PEBP-like Protein Promotes the Cap Redifferentiation in Cyclocybe aegerita

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Capebp2 Cloning and Sequence Analysis
2.3. Vector Construction and Transformation
2.4. Transformant Verification and Expression Analysis
2.5. Data Analysis
3. Results
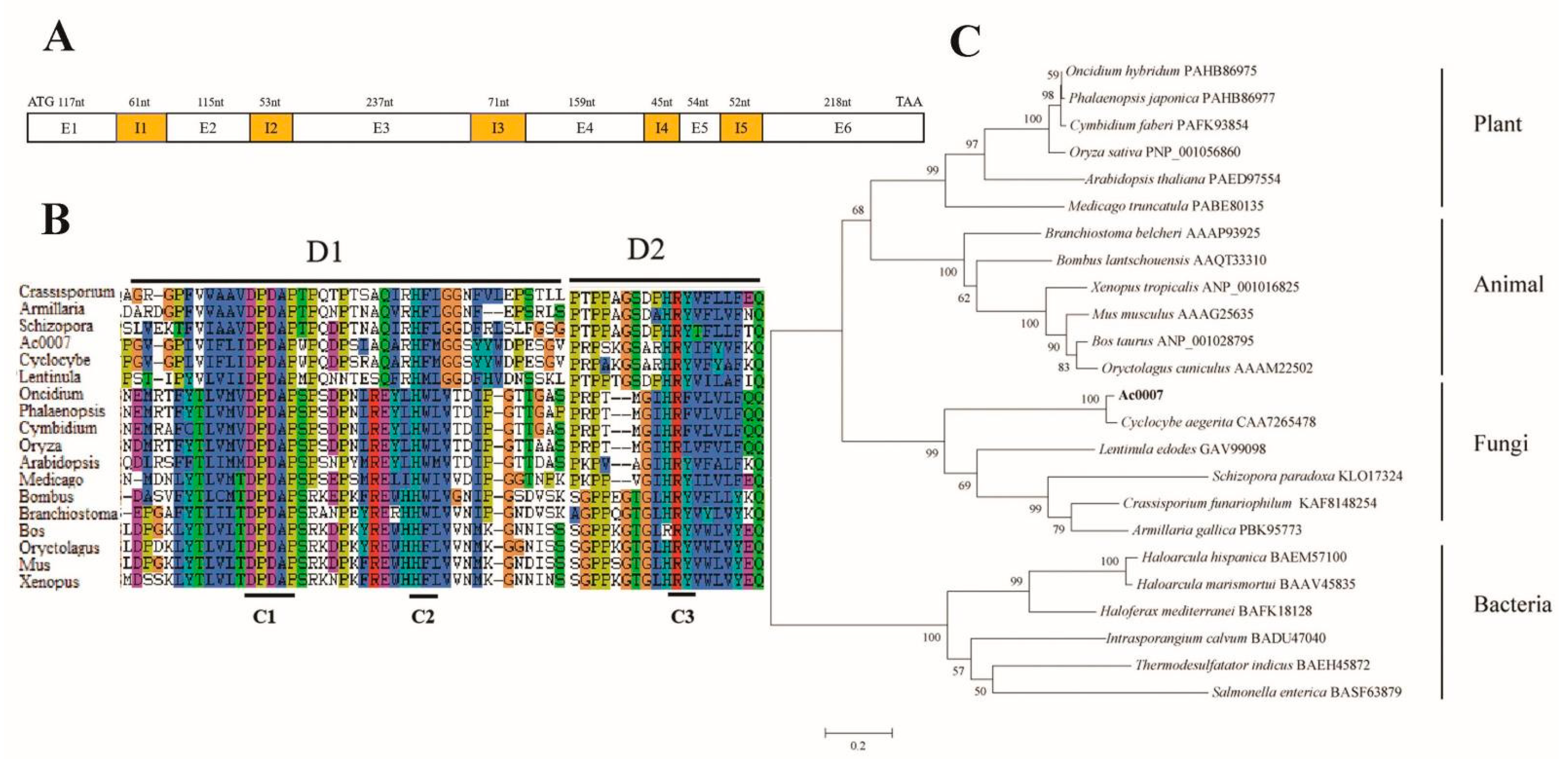
3.1. Sequence Analysis of Capebp2
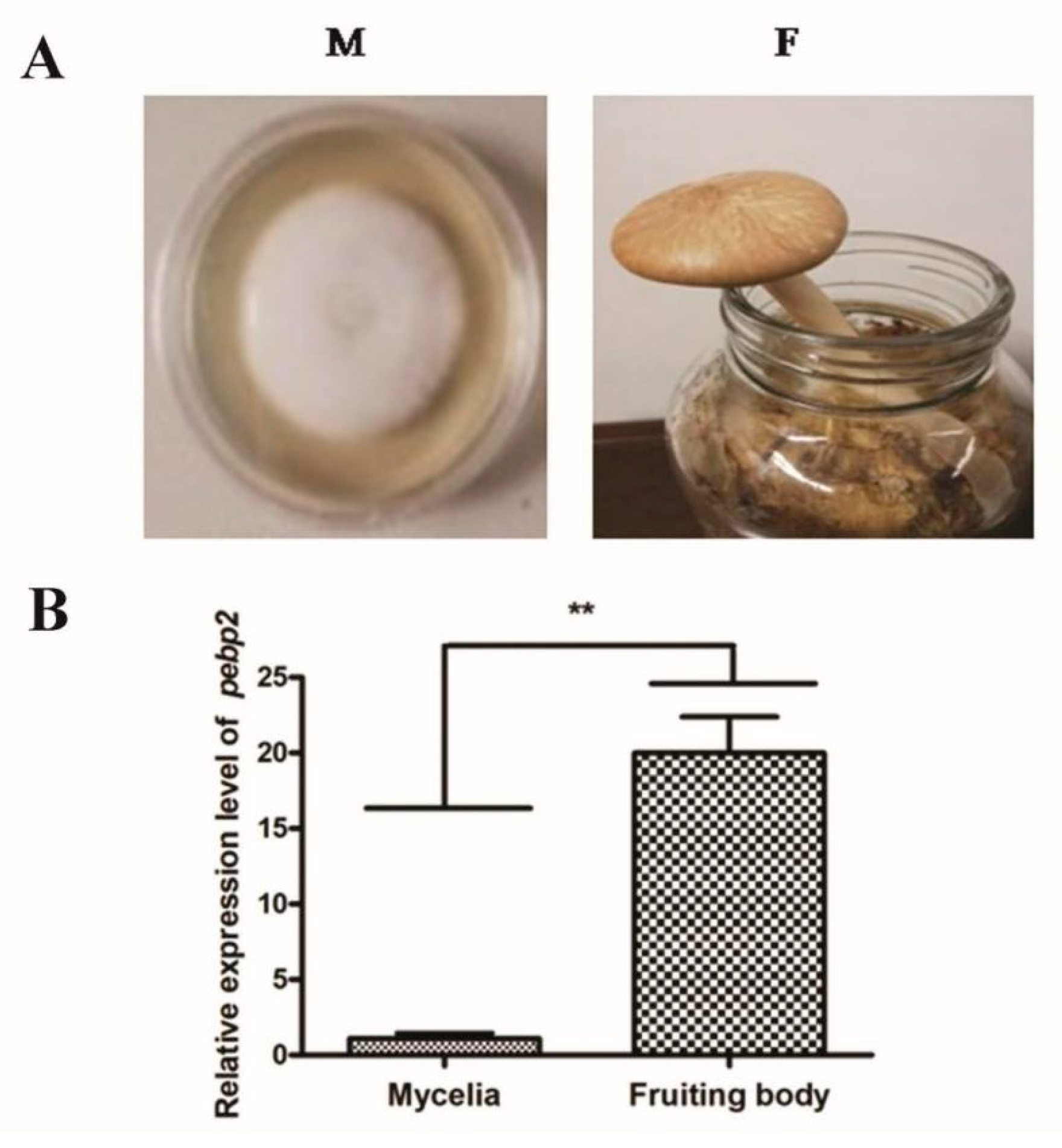
3.2. Characterization of the Expression of Capebp2 during the Development of C. aegerita
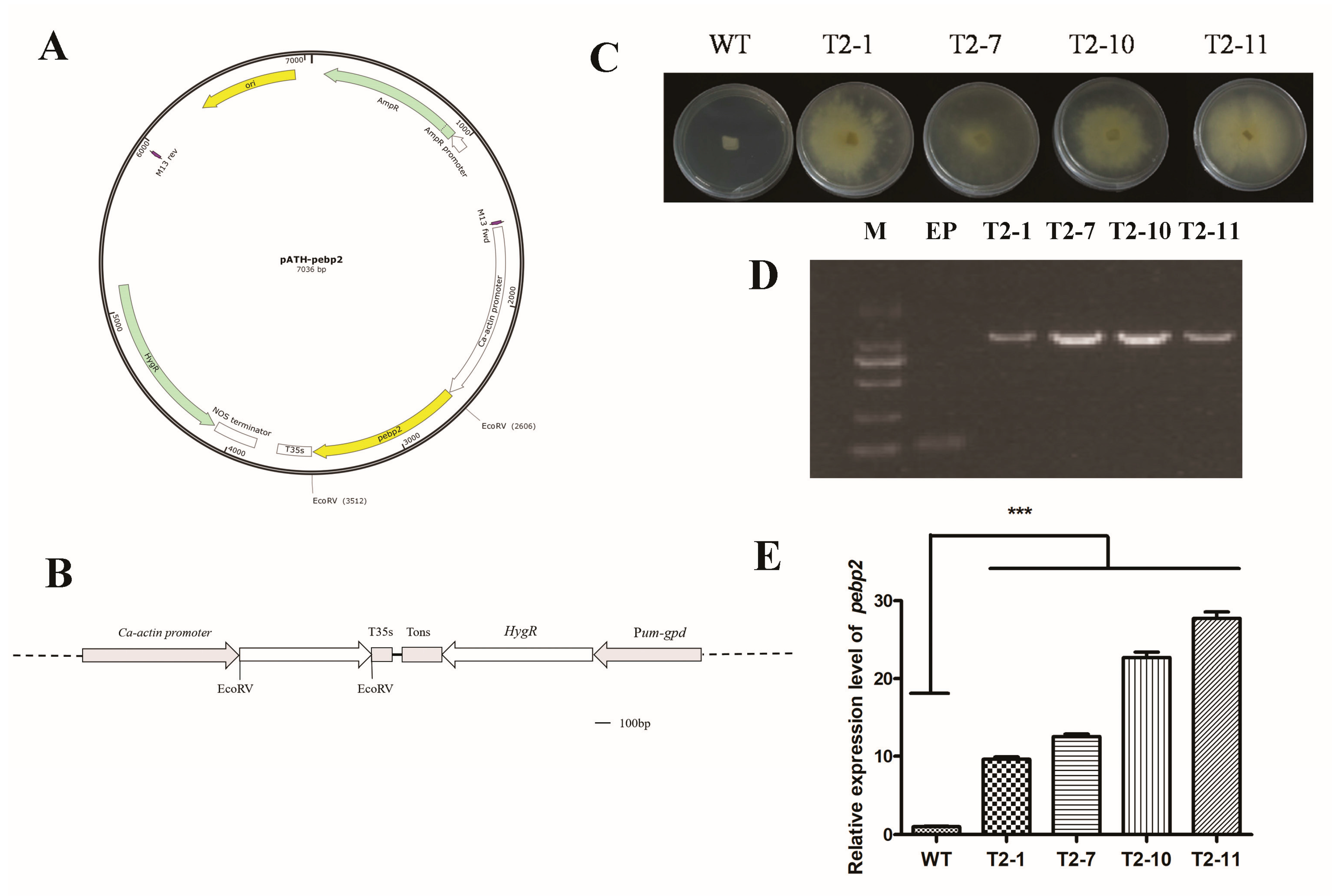
3.3. Validation of Transformants and Expression of Capebp2
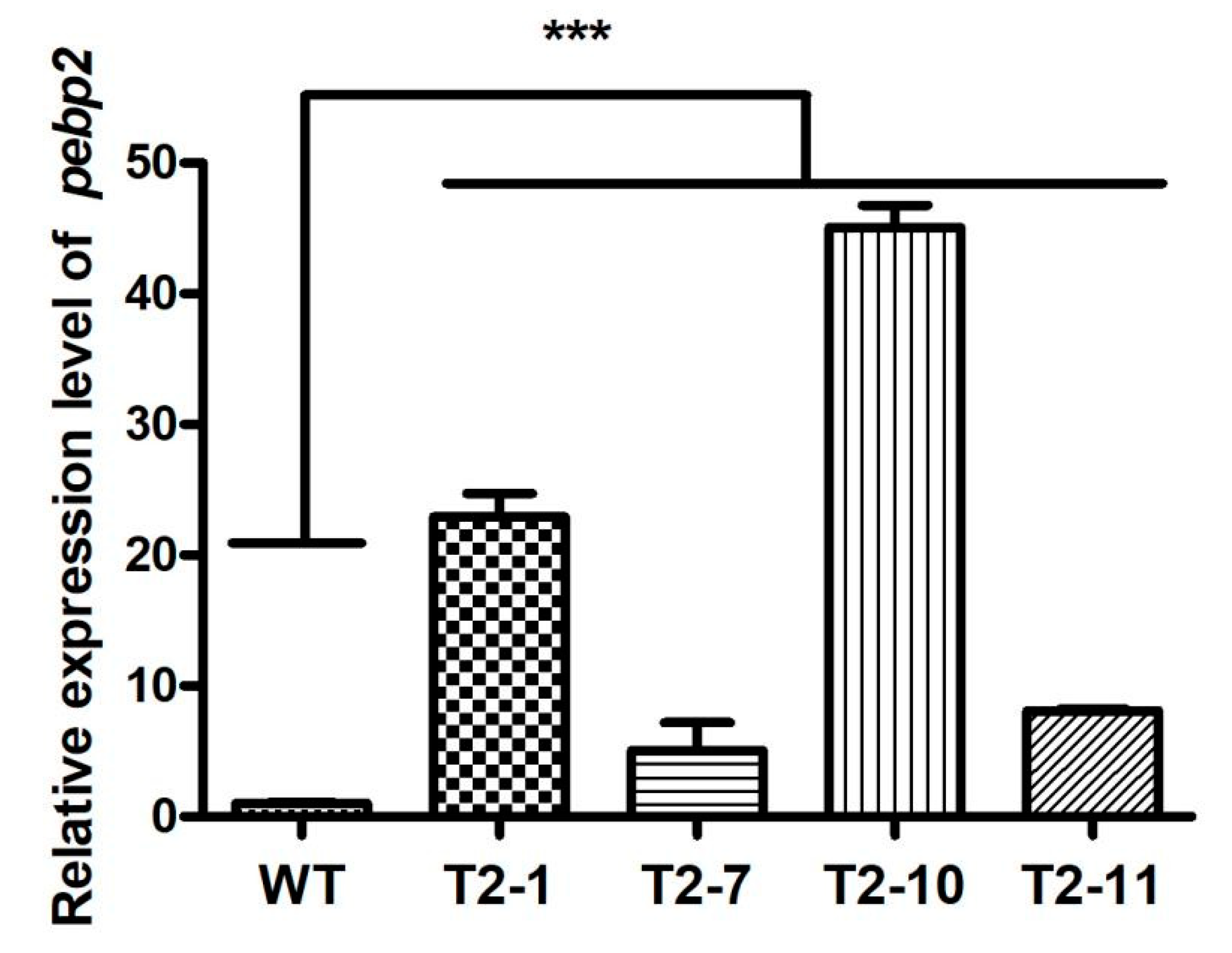
3.4. The Effect of Overexpression of Capebp2 on the Fruiting Bodies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bernier, I.D.A.; Jollés, P. Purification and characterization of a basic 23 kDa cytosolic protein from bovine brain. Biochim. Biophys. Acta (BBA)-Protein Struct. Mol. Enzymol. 1984, 790, 174–181. [Google Scholar] [CrossRef]
- Bernier, I.; Tresca, J.P.; Jollès, P. Ligand-binding studies with a 23 kDa protein purified from bovine brain cytosol. Biochim. Biophys. Acta (BBA)-Protein Struct. Mol. Enzymol. 1986, 871, 19–23. [Google Scholar] [CrossRef]
- Hickox, D.M.; Gibbs, G.; Morrison, J.R.; Sebire, K.; Edgar, K.; Keah, H.H.; Alter, K.; Loveland, K.L.; Hearn, M.T.W.; de Kretser, D.M. Identification of a novel testis-specific member of the phosphatidylethanolamine binding protein family, pebp-2. Biol. Reprod. 2002, 67, 917–927. [Google Scholar] [CrossRef] [Green Version]
- Toyoda, T.; Matsukawa, N.; Sagisaka, T.; Uematsu, N.; Kanamori, T.; Kato, D.; Mizuno, M.; Wake, H.; Hida, H.; Borlongan, C.V.; et al. Suppression of astrocyte lineage in adult hippocampal progenitor cells expressing hippocampal cholinergic neurostimulating peptide precursor in an in vivo ischemic model. Cell Transplant. 2012, 21, 2159–2169. [Google Scholar] [CrossRef] [Green Version]
- Al-Mulla, F.; Bitar, M.S.; Al-Maghrebi, M.; Behbehani, A.I.; Al-Ali, W.; Rath, O.; Doyle, B.; Tan, K.Y.; Pitt, A.; Kolch, W. Raf kinase inhibitor protein RKIP enhances signaling by glycogen synthase kinase-3β. Cancer Res. 2011, 71, 1334–1343. [Google Scholar] [CrossRef] [Green Version]
- Eves, E.M.; Rosner, M.R. MAP kinase regulation of the mitotic spindle checkpoint. Methods Mol. Biol. 2010, 661, 497–505. [Google Scholar]
- Chailakhyan, M.K. New facts in support of the hormonal theory of plant development. Dokl. Akad. Nauk 1936, 13, 79–83. [Google Scholar]
- Putterill, J.; Varkonyi-Gasic, E. FT and florigen long-distance flowering control in plants. Curr. Opin. Plant Biol. 2016, 33, 77–82. [Google Scholar] [CrossRef]
- Danilevskaya, O.N.; Meng, X.; Hou, Z.L.; Ananiev, E.V.; Simmons, C.R. A genomic and expression compendium of the expanded PEBP gene family from maize. Plant Physiol. 2008, 146, 250–264. [Google Scholar] [CrossRef] [Green Version]
- Karlgren, A.; Gyllenstrand, N.; Källman, T.; Sundström, J.F.; Moore, D.; Lascoux, M.; Lagercrantz, U. Evolution of the PEBP gene family in plants: Functional diversification in seed plant evolution. Plant Physiol. 2011, 156, 1967–1977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lifschitz, E.; Eviatar, T.; Rozman, A.; Shalit, A.; Goldshmidt, A.; Amsellem, Z.; Alvarez, J.P.; Eshed, Y. The tomato FT ortholog triggers systemic signals that regulate growth and flowering and substitute for diverse environmental stimuli. Proc. Natl. Acad. Sci. USA 2006, 103, 6398–6403. [Google Scholar] [CrossRef] [Green Version]
- Corbesier, L.; Vincent, C.; Jang, S.; Fornara, F.; Fan, Q.Z.; Searle, I.; Giakountis, A.; Farrona, S.; Gissot, L.; Turnbull, C.; et al. FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis. Science 2007, 316, 1030–1033. [Google Scholar] [CrossRef] [Green Version]
- Tamaki, S.; Matsuo, S.; Wong, H.L.; Yokoi, S.; Shimamoto, K. Hd3a protein is a mobile flowering signal in rice. Science 2007, 316, 1033–1036. [Google Scholar] [CrossRef]
- Bradley, D.; Ratcliffe, O.; Vincent, C.; Carpenter, R.; Coen, E. Inflorescence commitment and architecture in Arabidopsis. Science 1997, 275, 80–83. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.B.; Luo, L.; He, L.L.; Ni, J.; Xu, Z. A promoter analysis of MOTHER of FT and TFL11 (JcMFT1), a seed-preferential gene from the biofuel plant Jatropha curcas. J. Plant Res. 2014, 127, 513–524. [Google Scholar] [CrossRef]
- Li, Q.; Fan, C.M.; Zhang, X.M.; Wang, X.; Wu, F.Q.; Hu, R.B.; Fu, Y.F. Identification of a soybean MOTHER of FT and TFL1 homolog involved in regulation of seed germination. PLoS ONE 2014, 9, e99642. [Google Scholar] [CrossRef] [PubMed]
- Kardailsky, I.; Shukla, V.K.; Ahn, J.H.; Dagenais, N.; Christensen, S.K.; Nguyen, J.T.; Chory, J.; Harrison, M.J.; Weigel, D. Activation tagging of the floral inducer FT. Science 1999, 286, 1962–1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, A.; Kobayashi, Y.; Goto, K.; Abe, M.; Araki, T. TWIN SISTER of FT (TSF) acts as a floral pathway integrator redundantly with FT. Plant Cell Physiol. 2005, 46, 1175–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, S.K.; Chung, K.S.; Kim, J.; Lee, J.H.; Hong, S.M.; Yoo, S.J.; Yoo, S.Y.; Lee, J.S.; Ahn, J.H. Constans activates suppressor of overexpression of constans 1 through Flowering Locus T to promote flowering in Arabidopsis. Plant Physiol. 2005, 139, 770–778. [Google Scholar] [CrossRef] [Green Version]
- Xi, W.; Liu, C.; Hou, X.; Yu, H. MOTHER of FT and TFL1 regulates seed germination through a negative feedback loop modulating ABA signaling in Arabidopsis. Plant Cell 2010, 22, 1733–1748. [Google Scholar] [CrossRef] [Green Version]
- Hecht, V.; Laurie, R.E.; Vander Schoor, J.K.; Ridge, S.; Knowles, C.L.; Liew, L.C.; Sussmilch, F.C.; Murfet, I.C.; Macknight, R.C.; Weller, J.L. The pea GIGAS gene is a FLOWERING LOCUS T homolog necessary for graft-transmissible specification of flowering but not for responsiveness to photoperiod. Plant Cell 2011, 23, 147–161. [Google Scholar] [CrossRef] [Green Version]
- Faure, S.; Higgins, J.; Turner, A.; Laurie, D.A. The FLOWERING LOCUS T-like gene family in barley (Hordeum vulgare). Genetics 2007, 176, 599–609. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, R.; Wang, C.T.; Ma, C.; Shevchenko, O.; Dye, S.J.; Puzey, J.R.; Etherington, E.; Sheng, X.Y.; Meilan, R.; Strauss, S.H.; et al. Populus CEN/TFL1 regulates first onset of flowering, axillary meristem identity and dormancy release in Populus. Plant J. 2010, 62, 674–688. [Google Scholar] [CrossRef] [PubMed]
- Molinero-Rosales, N.; Latorre, A.; Jamilena, M.; Lozano, R. SINGLE FLOWER TRUSS regulates the transition and maintenance of flowering in tomato. Planta 2004, 218, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Pin, P.A.; Benlloch, R.; Bonnet, D.; Wremerth-Weich, E.; Kraft, T.; Gielen, J.J.L.; Nilsson, O. An antagonistic pair of FT homologs mediates the control of flowering time in sugar beet. Science 2010, 330, 1397–1400. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Liu, B.; Xia, Z.; Bo, M.K. Two coordinately regulated homologs of FLOWERING LOCUS T are involved in the control of photoperiodic flowering in soybean. Plant Physiol. 2010, 154, 1220–1231. [Google Scholar] [CrossRef] [Green Version]
- Navarro, C.; Abelenda, J.A.; Cruz-Oró, E.; Cuéllar, C.A.; Tamaki, S.; Silva, J.; Shimamoto, K.; Prat, S. Control of flowering and storage organ formation in potato by FLOWERING LOCUS T. Nature 2011, 478, 119–122. [Google Scholar] [CrossRef]
- Collani, S.; Neumann, M.; Yant, L.; Schimid, M. FT modulates genomewide DNA-binding of the bZIP transcription factor FD. Plant Physiol. 2019, 180, 367–380. [Google Scholar] [CrossRef] [Green Version]
- Goretti, D.; Silvestre, M.; Collani, S.; Langenecker, T.; Mendez, C.; Madueno, F.; Schmid, M. TERMINAL FLOWER1 functions as a mobile transcriptional cofactor in the shoot apical meristem. Plant Physiol. 2020, 182, 2081–2095. [Google Scholar] [CrossRef] [Green Version]
- Romera-Branchat, M.; Severing, E.; Pocard, C.; Ohr, H.; Vincent, C.; Née, G.; Martinez-Gallegos, R.; Jang, S.; Andrés, F.; Madrigal, P.; et al. Functional divergence of the Arabidopsis florigen-interacting bZIP transcription factors FD and FDP. Cell Rep. 2020, 31, 107717. [Google Scholar] [CrossRef]
- Zhu, Y.; Klasfeld, S.; Jeong, C.W.; Jin, R.; Goto, K.; Yamaguchi, N.; Wagner, D. TERMINAL FLOWER 1–FD complex target genes and competition with FLOWERING LOCUS T. Nat. Commun. 2020, 11, 5118. [Google Scholar] [CrossRef] [PubMed]
- Pnueli, L.; Carmel-Goren, L.; Hareven, D.; Gutfinger, T.; Alvarez, J.; Ganal, M.; Zamir, D.; Lifschitz, E. The SELF-PRUNING gene of tomato regulates vegetative to reproductive switching of sympodial meristems and is the ortholog of CEN and TFL1. Development 1998, 125, 1979–1989. [Google Scholar] [CrossRef]
- Silva, W.B.; Vicente, M.H.; Robledo, J.M.; Reartes, D.S.; Ferrari, R.C.; Bianchetti, R.; Araújo, W.L.; Freschi, L.; Peres, L.E.P.; Zsögön, A. SELF-PRUNING acts synergistically with DIAGEOTROPICA to guide auxin responses and proper growth form. Plant Physiol. 2018, 176, 2904–2916. [Google Scholar] [CrossRef] [Green Version]
- Cerise, M.; Giaume, F.; Galli, M.; Khahani, B.; Lucas, J.; Podico, F.; Tavakol, E.; Parcy, F.; Gallavotti, A.; Brambilla, V.; et al. OsFD4 promotes the rice floral transition via florigen activation complex formation in the shoot apical meristem. New Phytol. 2021, 229, 429–443. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Thames, S.; Best, N.B.; Jiang, H.; Huang, P.; Dilkes, B.P.; Eveland, A.L. Brassinosteroids modulate meristem fate and differentiation of unique infforescence morphology in Setaria viridis. Plant Cell 2018, 30, 48–66. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Gao, Y.; Liu, Y.; Zhang, X.; Gu, X.X.; Ma, D.B.; Zhao, Z.W.; Yuan, Z.J.; Xue, H.W.; Liu, H.T. BES1-regulated BEE1 controls photoperiodic flowering downstream of blue light signaling pathway in Arabidopsis. New Phytol. 2019, 223, 1407–1419. [Google Scholar] [CrossRef]
- Riboni, M.; Robustelli, T.A.; Galbiati, M.; Tonelli, C.; Conti, L. ABA dependent control of GIGANTEA signalling enables drought escape via up-regulation of FLOWERING LOCUS T in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 6309–6322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, G.X.; Yang, W.J.; Zhang, Q.L.; Huang, J.L.; Yang, Y.P.; Hu, X.Y. ABI5- BINDING PROTEIN2 coordinates CONSTANS to delay flowering by recruiting the transcriptional corepressor TPR2. Plant Physiol. 2019, 179, 477–490. [Google Scholar] [CrossRef] [Green Version]
- Vaistij, F.E.; Gan, Y.; Penffeld, S.; Gilday, A.D.; Dave, A.; He, Z.; Josse, E.M.; Choi, G.; Halliday, K.J.; Graham, I.A. Differential control of seed primary dormancy in Arabidopsis ecotypes by the transcription factor SPATULA. Proc. Natl. Acad. Sci. USA 2013, 110, 10866–10871. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol Biol Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Tao, N.; Cui, X.H.; Chai, H.M.; Ma, Y.H.; Zhao, Y.C.; Chen, W.M. Establishment of genetic transformation system of Agrocybe aegerita using PEG-mediated method. Mycosystema 2020, 39, 1100–1108. [Google Scholar]
- Maurya, J.P.; Miskolczi, P.C.; Mishra, S.; Singh, R.K.; Bhalerao, R.P. A genetic framework for regulation and seasonal adaptation of shoot architecture in hybrid aspen. Proc. Natl. Acad. Sci. USA 2020, 117, 11523–11530. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.K.; Xu, X.P.; Chen, X.H.; Chen, Y.; Zhang, Z.H.; Xu, H.X.; Lin, Y.L.; Lai, Z.X. Seed-Specific Gene MOTHER of FT and TFL1 (MFT) Involved in Embryogenesis, Hormones and Stress Responses in Dimocarpus longan Lour. Int. J. Mol. Sci. 2018, 19, 2403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Périlleux, C.; Bouché, F.; Randoux, M.; Orman-Ligeza, B. Turning meristems into fortresses. Trends Plant Sci. 2019, 24, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Nasim, Z.; Susila, H.; Ji, H.A. Evolution and functional diversification of FLOWERING LOCUS T/TERMINAL FLOWER 1 family genes in plants. Semin. Cell Dev. Biol. 2021, 109, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Liu, C.; Hou, X.L.; Xi, W.Y.; Shen, L.S.; Tao, Z.; Wang, Y.; Yu, H. FTIP1 is an essential regulator required for florigen transport. PLoS Biol. 2012, 10, e1001313. [Google Scholar] [CrossRef] [Green Version]
- Jackson, S.D.; Hong, Y. Systemic movement of FT mRNA and a possible role in floral induction. Front. Plant Sci. 2012, 3, 127. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Zhu, Y.; Shen, L.S.; Yu, H. Emerging insights into florigen transport. Curr. Opin. Plant Biol. 2013, 16, 607–613. [Google Scholar] [CrossRef]
- Madelin, M.F. The influence of light and temperature on fruiting of Coprinus lagopus Fr. in pure culture. Ann. Bot. 1956, 20, 467–480. [Google Scholar] [CrossRef]
- Manachère, G. Conditions essential for controlled fruiting of macromycetes—A review. Trans. Brit. Mycol. Soc. 1980, 75, 255–270. [Google Scholar] [CrossRef]
- Kües, U.; Liu, Y. Fruiting body production in basidiomycetes. Appl. Microbiol. Biotechnol. 2000, 54, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Eastwood, D.C.; Herman, B.; Noble, R.; DobrovinPennington, A.; Sreenivasaprasad, S.; Burton, K.S. Environmental regulation of reproductive phase change in Agaricus bisporus by 1-octen-3-ol, temperature and CO2. Fungal Genet. Biol. 2013, 55, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Perkins, J.H. Morphogenesis in Schizophyllum commune. I. Effects of white light. Plant Physiol. 1969, 44, 1706–1711. [Google Scholar] [CrossRef] [Green Version]
- Raudaskoski, M.; Yli-Mattila, T. Capacity for photoinduced fruiting in the dikaryon of Schizophyllum commune. Trans. Br. Mycol. Soc. 1985, 85, 145–151. [Google Scholar] [CrossRef]
- Kües, U. Life history and developmental processes in the basidiomycete Coprinus cinereus. Microbiol. Mol. Biol. Rev. 2000, 64, 316–353. [Google Scholar] [CrossRef] [Green Version]
- Lu, B.C. The control of meiosis progression in the fungus Coprinus cinereus by light/dark cycles. Fungal Genet. Biol. 2000, 31, 33–41. [Google Scholar] [CrossRef]
- Blumenstein, A.; Vienken, K.; Tasler, R.; Purschwitz, J.; Veith, D.; Frankenberg-Dinkel, N.; Fischer, R. The Aspergillus nidulans phytochrome FphA represses sexual development in red light. Curr. Biol. 2005, 15, 1833–1838. [Google Scholar] [CrossRef]
- Purschwitz, J.; Müller, S.; Kastner, C.; Schöser, M.; Haas, H.; Espeso, E.A.; Atoui, A.; Calvo, A.M.; Fischer, R. Functional and physical interaction of blue- and red-light sensors in Aspergillus nidulans. Curr. Biol. 2008, 18, 255–259. [Google Scholar] [CrossRef] [Green Version]
- Muraguchi, H.; Fujita, T.; Kishibe, Y.; Konno, K.; Ueda, N.; Nakahori, K.; Yanagi, S.O.; Kamada, T. The exp1 gene essential for pileus expansion and autolysis of the inky cap mushroom Coprinopsis cinerea (Coprinus cinereus) encodes an HMG protein. Fungal Genet. Biol. 2008, 45, 890–896. [Google Scholar] [CrossRef] [PubMed]
- Teo, C.J.; Takahashi, K.; Shimizu, K.; Shimamoto, K.; Taoka, K.I. Potato tuber induction is regulated by interactions between components of a tuberigen complex. Plant Cell Physiol. 2016, 58, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Kaneko-Suzuki, M.; Kurihara-Ishikawa, R.; Okushita-Terakawa, C.; Nagano-Fujiwara, M.; Ohki, I.; Tsuji, H.; Shimamoto, K.; Taoka, K.I. TFL1-like proteins in rice antagonize rice FT-like protein in inflorescence development by competition for complex formation with 14-3-3 and FD. Plant Cell Physiol. 2018, 59, 458–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Campbell, R.; Ducreux, L.J.M.; Morris, J.; Hedley, P.E.; Mellado-Ortega, E.; Roberts, A.G.; Stephens, J.; Bryan, G.J.; Torrance, L.; et al. TERMINAL FLOWER-1 /CENTRORADIALIS inhibits tuberisation via protein interaction with the tuberigen activation complex. Plant J. 2020, 103, 2263–2278. [Google Scholar] [CrossRef] [PubMed]
- Fornara, F.; de Montaigu, A.; Coupland, G. SnapShot: Control of flowering in Arabidopsis. Cell 2010, 141, 550. [Google Scholar] [CrossRef] [Green Version]
- Wagner, D. Key developmental transitions during flower morphogenesis and their regulation. Curr. Opin. Genet. Dev. 2017, 45, 44–50. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′ to 3′) | Description |
|---|---|---|
| capebpF | TAACGAATAATAGCCGATATCATGCGTTCTCTCATCCTCTTCATC | Amplification for the whole length of Capebp2 |
| capebpR | CCGGTCGGCATCTACGATATCTTAGTCCCCATACCTGAACTTAAAC | |
| 19ha3 | TCACCGTAACGAATAATAGCC | Verification of transformants |
| 19ha4 | CCCTTATCTGGGAACTACTCAC | |
| Gpd qF | AGGCTGTCGGCAAGGTTATC | Detection for gpd expression level |
| Gpd qR | TGCGGTGTGACCAATGAAG | |
| Pebp qF | CAAGCGAAGAAGGCACTAATC | Detection for Capebp2 expression level |
| Pebp qR | TAAACGAAGGACTCAGGCG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, B.; Tao, N.; Ma, Y.; Chai, H.; Liu, P.; Chen, W.; Zhao, Y. Overexpression of the Capebp2 Gene Encoding the PEBP-like Protein Promotes the Cap Redifferentiation in Cyclocybe aegerita. J. Fungi 2023, 9, 657. https://doi.org/10.3390/jof9060657
Cheng B, Tao N, Ma Y, Chai H, Liu P, Chen W, Zhao Y. Overexpression of the Capebp2 Gene Encoding the PEBP-like Protein Promotes the Cap Redifferentiation in Cyclocybe aegerita. Journal of Fungi. 2023; 9(6):657. https://doi.org/10.3390/jof9060657
Chicago/Turabian StyleCheng, Bopu, Nan Tao, Yuanhao Ma, Hongmei Chai, Ping Liu, Weimin Chen, and Yongchang Zhao. 2023. "Overexpression of the Capebp2 Gene Encoding the PEBP-like Protein Promotes the Cap Redifferentiation in Cyclocybe aegerita" Journal of Fungi 9, no. 6: 657. https://doi.org/10.3390/jof9060657