
Endophytic Fungal Community of Stellera chamaejasme L. and Its Possible Role in Improving Host Plants’ Ecological Flexibility in Degraded Grasslands

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Culturable Endophytic Fungal Community Analysis
2.3. Culture-Independent Endophytic Fungal Community Analysis
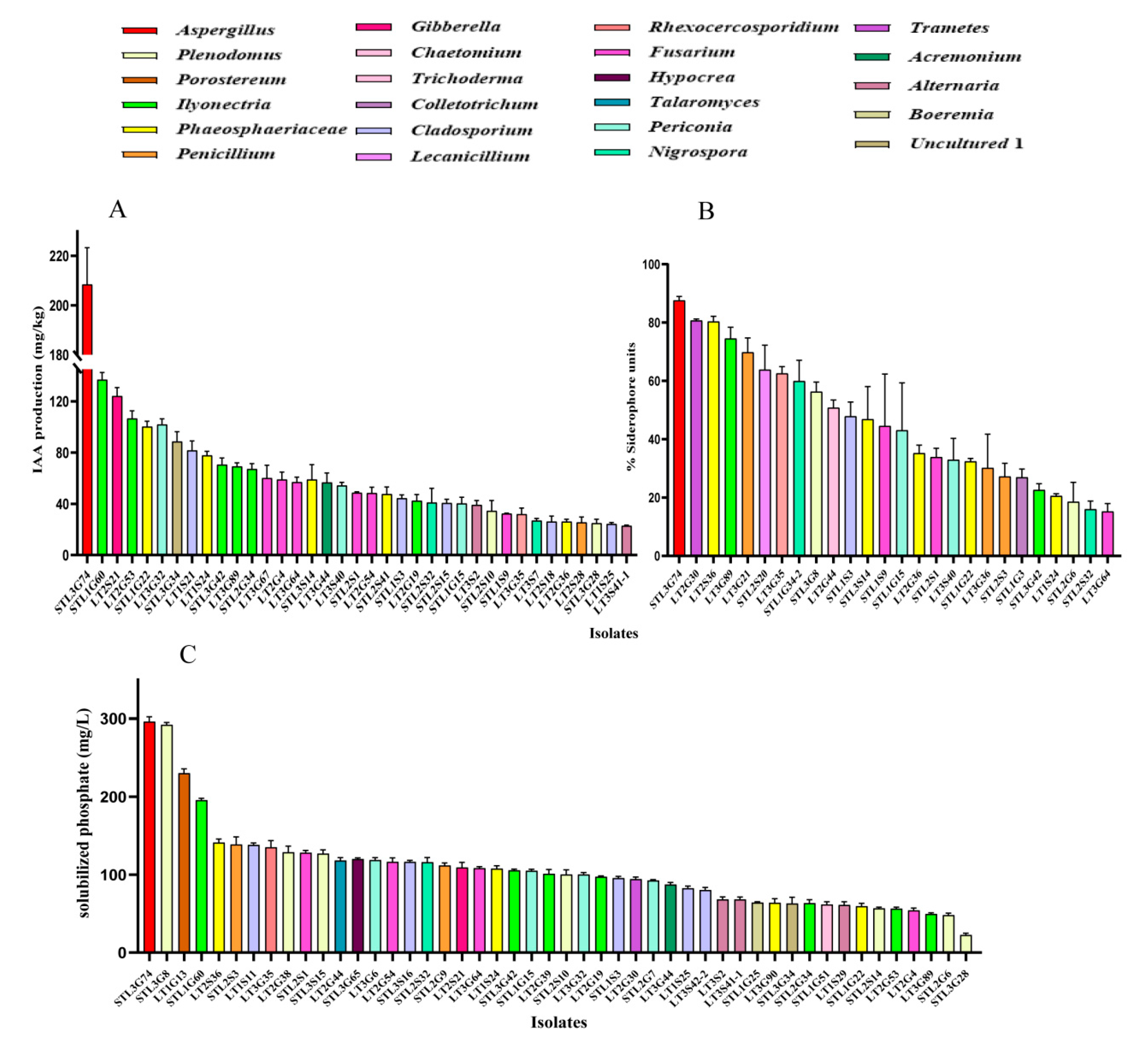
2.4. Growth-Promoting Traits Test
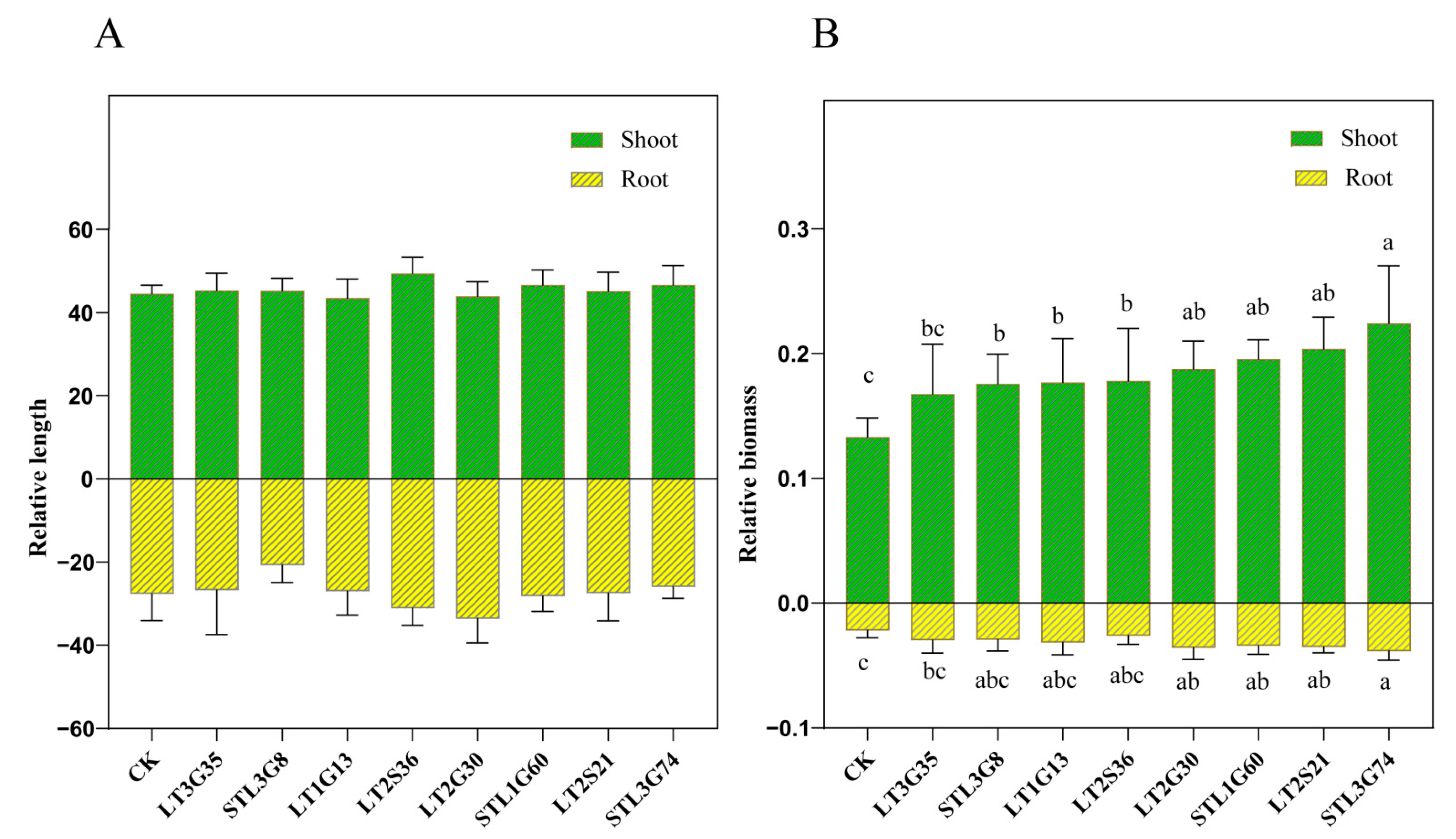
2.5. Pot Experiments
2.6. Statistical Analysis
3. Results
3.1. Culturable Endophytic Fungal Community
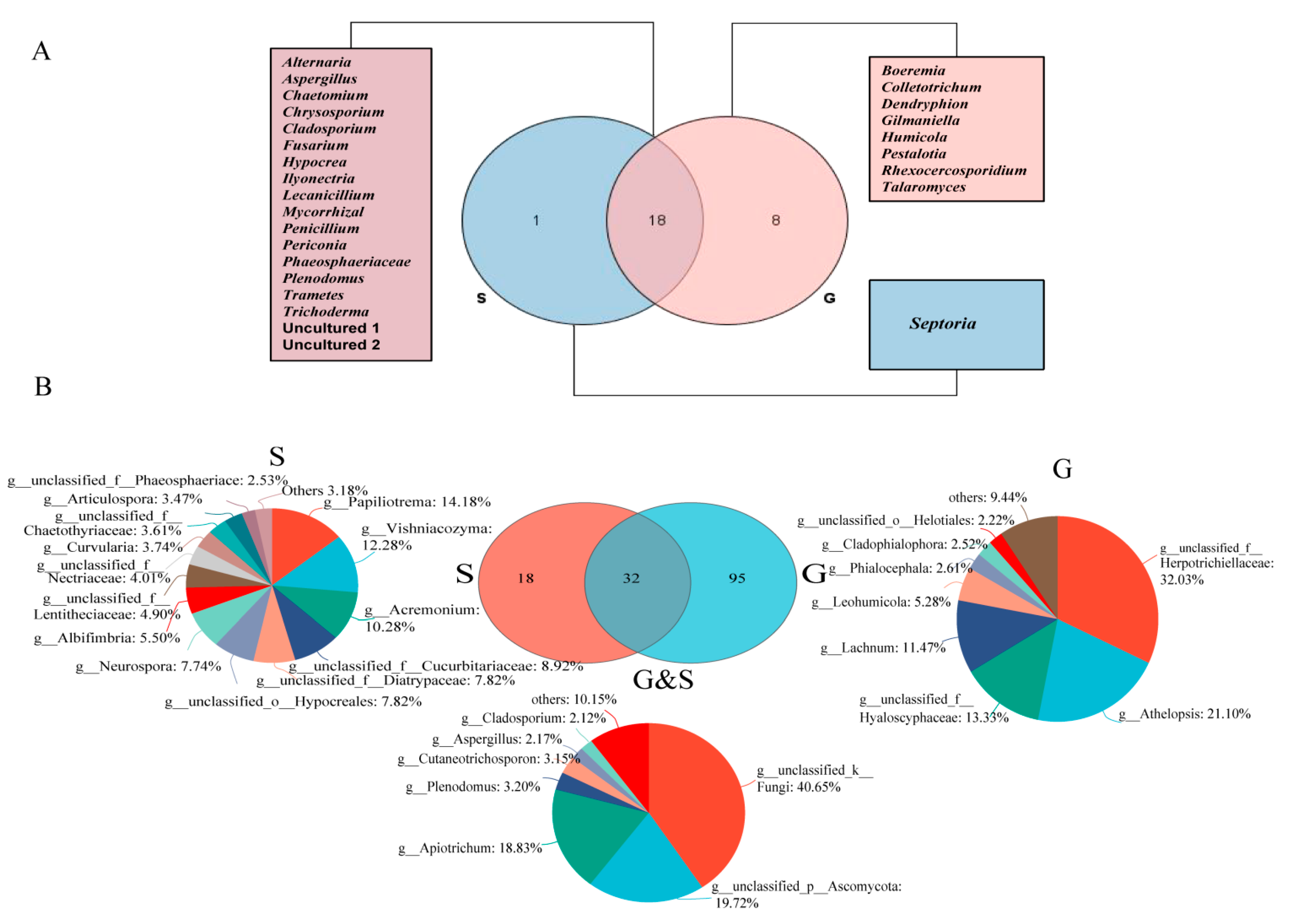
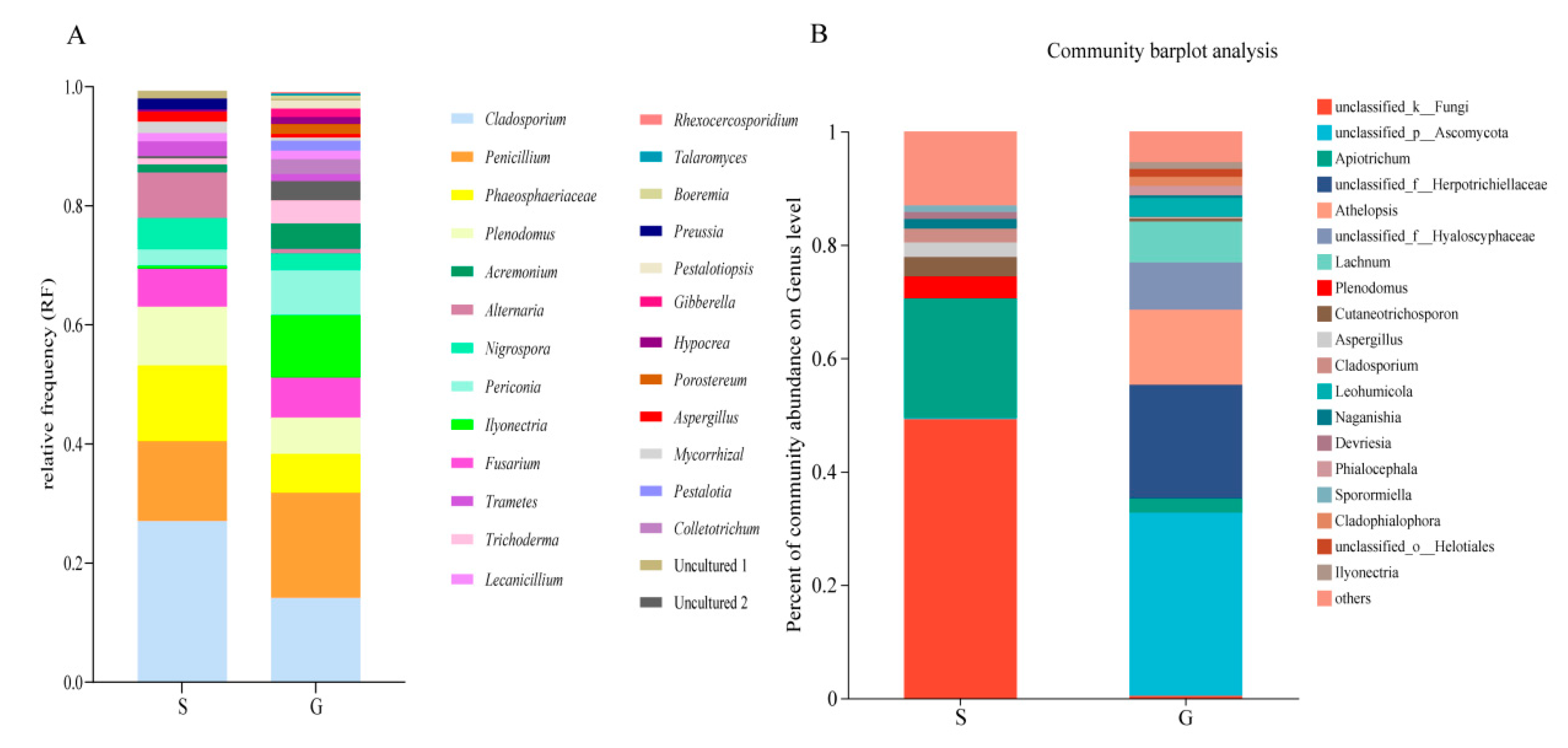
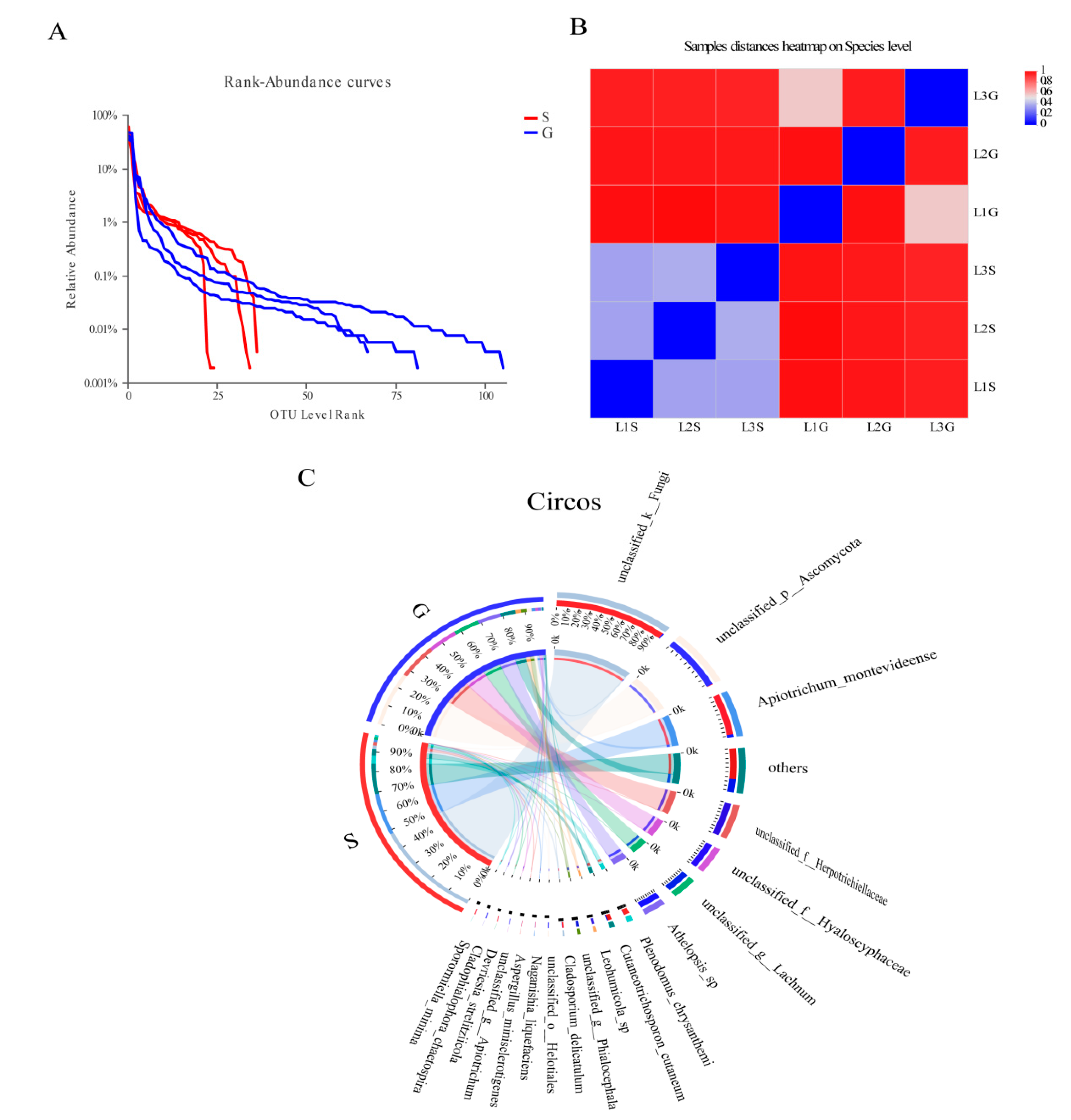
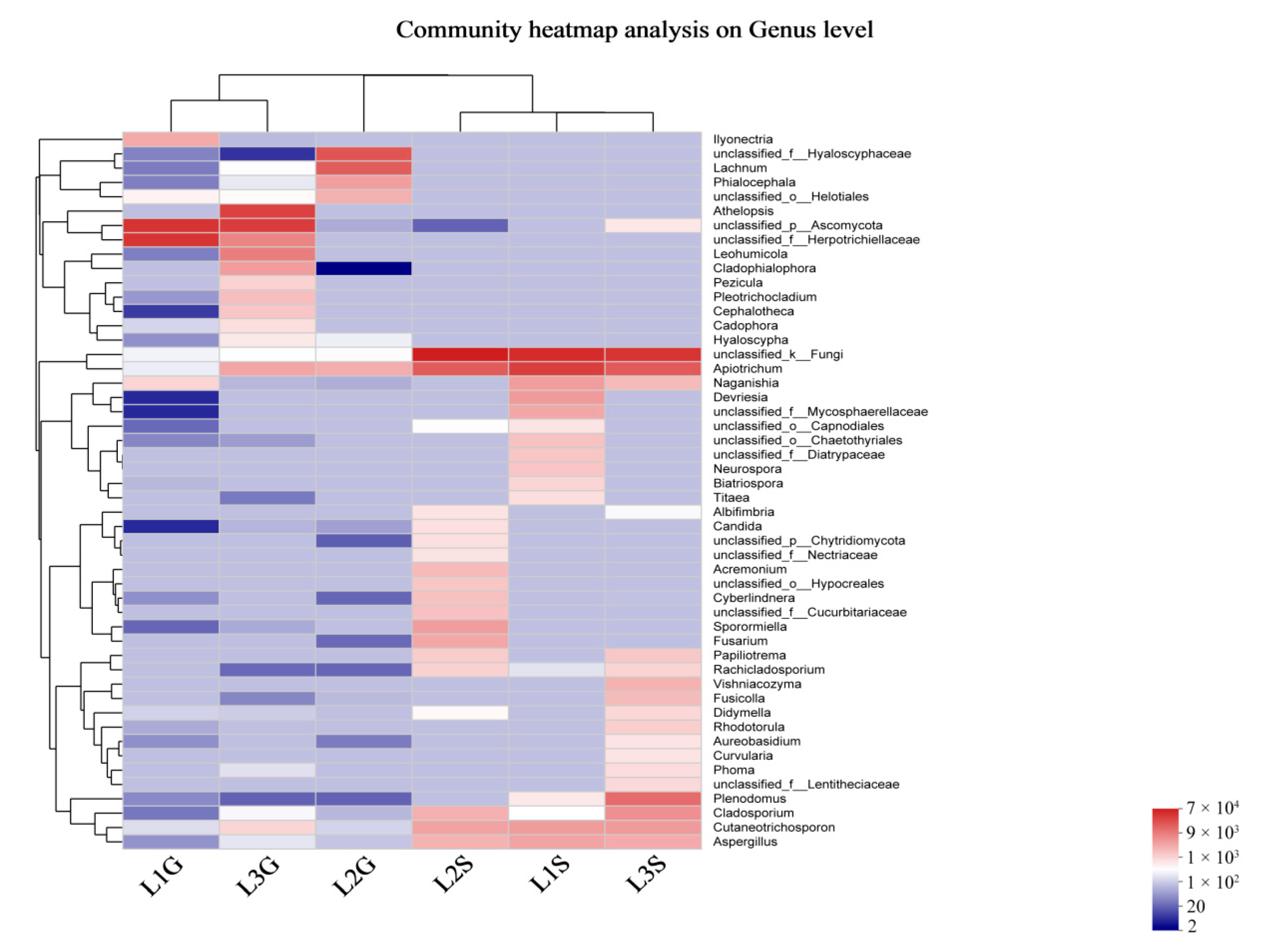
3.2. Culture-Independent Endophytic Fungal Community
3.3. PGP Traits of Fungal Endophytes
3.4. Pot Experiments
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Y.; Wesche, K. Vegetation and Soil Responses to Livestock Grazing in Central Asian Grasslands: A Review of Chinese Literature. Biodivers. Conserv. 2016, 25, 2401–2420. [Google Scholar] [CrossRef]
- Meng, B.; Ge, J.; Liang, T.; Yang, S.; Gao, J.; Feng, Q.; Cui, X.; Huang, X.; Xie, H. Evaluation of Remote Sensing Inversion Error for the Above-Ground Biomass of Alpine Meadow Grassland Based on Multi-Source Satellite Data. Remote Sens. 2017, 9, 372. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Yang, X.C.; Tao, W.G.; Miao, J.M.; Yang, Z.; Liu, H.Q.; Jin, Y.X.; Zhu, X.H.; Qin, Z.H.; Lv, H.Y.; et al. MODIS-Based Remote-Sensing Monitoring of the Spatiotemporal Patterns of China’s Grassland Vegetation Growth. Int. J. Remote Sens. 2013, 34, 3867–3878. [Google Scholar] [CrossRef]
- Zhou, W.; Yang, H.; Huang, L.; Chen, C.; Lin, X.; Hu, Z.; Li, J. Grassland Degradation Remote Sensing Monitoring and Driving Factors Quantitative Assessment in China from 1982 to 2010. Ecol. Indic. 2017, 83, 303–313. [Google Scholar] [CrossRef]
- Zhang, Z.; Sun, J.; Liu, M.; Xu, M.; Wang, Y.; Wu, G.; Zhou, H.; Ye, C.; Tsechoe, D.; Wei, T. Don’t Judge Toxic Weeds on Whether They Are Native but on Their Ecological Effects. Ecol. Evol. 2020, 10, 9014–9025. [Google Scholar] [CrossRef]
- Pan, L.; Li, X.; Yan, Z.; Guo, H.; Qin, B. Phytotoxicity of Umbelliferone and Its Analogs: Structure–Activity Relationships and Action Mechanisms. Plant Physiol. Biochem. 2015, 97, 272–277. [Google Scholar] [CrossRef]
- Guo, L.; Li, J.; He, W.; Liu, L.; Huang, D.; Wang, K. High Nutrient Uptake Efficiency and High Water Use Efficiency Facilitate the Spread of Stellera Chamaejasme L. in Degraded Grasslands. BMC Ecol. 2019, 19, 50. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.-H.; Volis, S.; Sun, H. Chloroplast Phylogeny and Phylogeography of Stellera Chamaejasme on the Qinghai-Tibet Plateau and in Adjacent Regions. Mol. Phylogenet. Evol. 2010, 57, 1162–1172. [Google Scholar] [CrossRef]
- Liang, C.; Xie, J.; Yan, J. The Complete Chloroplast Genome Sequence of Stellera chamaejasme f . chrysantha (Thymelaeaceae). Mitochondrial DNA Part B 2020, 5, 3251–3252. [Google Scholar] [CrossRef]
- Sun, G.; Luo, P.; Wu, N.; Qiu, P.F.; Gao, Y.H.; Chen, H.; Shi, F.S. Stellera chamaejasme L. Increases Soil N Availability, Turnover Rates and Microbial Biomass in an Alpine Meadow Ecosystem on the Eastern Tibetan Plateau of China. Soil Biol. Biochem. 2009, 41, 86–91. [Google Scholar] [CrossRef]
- Song, M.; Wang, Y.; Bao, G.; Wang, H.; Yin, Y.; Li, X.; Zhang, C. Effects of Stellera chamaejasme Removal on the Nutrient Stoichiometry of S. chamaejasme-Dominated Grasslands in the Qinghai–Tibetan Plateau. PeerJ 2020, 8, e9239. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Zhao, H.; Zhai, X.; Wang, K.; Liu, L.; Wang, K.; Huang, D. Study on Life Histroy Traits of Stellera Chamaejasme Provide Insights into Its Control on Degraded Typical Steppe. J. Environ. Manag. 2021, 291, 112716. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Detheridge, A.; Liu, Y.; Wang, L.; Wei, H.; Griffith, G.W.; Scullion, J.; Wei, Y. Variation in Soil Fungal Composition Associated with the Invasion of Stellera chamaejasme L. in Qinghai–Tibet Plateau Grassland. Microorganisms 2019, 7, 587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.-Y.; Wei, D.-Q.; Shen, M.; Zhou, Z.-P. Endophytes and Their Role in Phytoremediation. Fungal Divers. 2012, 54, 11–18. [Google Scholar] [CrossRef]
- Chitnis, V.R.; Suryanarayanan, T.S.; Nataraja, K.N.; Prasad, S.R.; Oelmüller, R.; Shaanker, R.U. Fungal Endophyte-Mediated Crop Improvement: The Way Ahead. Front. Plant Sci. 2020, 11, 561007. [Google Scholar] [CrossRef]
- Chen, Z.; Jin, Y.; Yao, X.; Chen, T.; Wei, X.; Li, C.; White, J.F.; Nan, Z. Fungal Endophyte Improves Survival of Lolium Perenne in Low Fertility Soils by Increasing Root Growth, Metabolic Activity and Absorption of Nutrients. Plant Soil 2020, 452, 185–206. [Google Scholar] [CrossRef]
- Buckley, H.; Young, C.A.; Charlton, N.D.; Hendricks, W.Q.; Haley, B.; Nagabhyru, P.; Rudgers, J.A. Leaf Endophytes Mediate Fertilizer Effects on Plant Yield and Traits in Northern Oat Grass (Trisetum spicatum). Plant Soil 2019, 434, 425–440. [Google Scholar] [CrossRef]
- Baron, N.C.; Costa, N.T.A.; Mochi, D.A.; Rigobelo, E.C. First Report of Aspergillus Sydowii and Aspergillus Brasiliensis as Phosphorus Solubilizers in Maize. Ann. Microbiol. 2018, 68, 863–870. [Google Scholar] [CrossRef]
- Jin, H.; Yan, Z.; Liu, Q.; Yang, X.; Chen, J.; Qin, B. Diversity and Dynamics of Fungal Endophytes in Leaves, Stems and Roots of Stellera Chamaejasme L. in Northwestern China. Antonie Van Leeuwenhoek 2013, 104, 949–963. [Google Scholar] [CrossRef]
- Jin, H.; Yang, X.; Lu, D.; Li, C.; Yan, Z.; Li, X.; Zeng, L.; Qin, B. Phylogenic Diversity and Tissue Specificity of Fungal Endophytes Associated with the Pharmaceutical Plant, Stellera chamaejasme L. Revealed by a Cultivation-Independent Approach. Antonie Van Leeuwenhoek 2015, 108, 835–850. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, J.; Chen, Y.; Cheng, F.; Liu, G.; He, Z. Remote-Sensing Monitoring of Grassland Degradation Based on the GDI in Shangri-La, China. Remote Sens. 2019, 11, 3030. [Google Scholar] [CrossRef] [Green Version]
- Vannier, N.; Bittebiere, A.-K.; Mony, C.; Vandenkoornhuyse, P. Root Endophytic Fungi Impact Host Plant Biomass and Respond to Plant Composition at Varying Spatio-Temporal Scales. Fungal Ecol. 2020, 44, 100907. [Google Scholar] [CrossRef]
- Li, H.-Y.; Shen, M.; Zhou, Z.-P.; Li, T.; Wei, Y.; Lin, L. Diversity and Cold Adaptation of Endophytic Fungi from Five Dominant Plant Species Collected from the Baima Snow Mountain, Southwest China. Fungal Divers. 2012, 54, 79–86. [Google Scholar] [CrossRef]
- Li, X.; Li, W.; Chu, L.; White, J.F.; Xiong, Z.; Li, H. Diversity and Heavy Metal Tolerance of Endophytic Fungi from Dysphania Ambrosioides, a Hyperaccumulator from Pb–Zn Contaminated Soils. J. Plant Interact. 2016, 11, 186–192. [Google Scholar] [CrossRef] [Green Version]
- Pan, H.; Yu, J.; Wu, Y.; Zhang, T.; Wang, H. Diversity Analysis of Soil Dematiaceous Hyphomycetes from the Yellow River Source Area: I. J. Zhejiang Univ. Sci. B 2008, 9, 829–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter-Cevera, J.C.; Sotos, L. Screening for a “New” Enzyme in Nature: Haloperoxidase Production by Death Valley Dematiaceous Hyphomycetes. Microb. Ecol. 1986, 12, 121–127. [Google Scholar] [CrossRef]
- Jain, R.; Bhardwaj, P.; Pandey, S.S.; Kumar, S. Arnebia Euchroma, a Plant Species of Cold Desert in the Himalayas, Harbors Beneficial Cultivable Endophytes in Roots and Leaves. Front. Microbiol. 2021, 12, 696667. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Zhang, C.; Lin, F.; Kubicek, C.P. Identity, Diversity, and Molecular Phylogeny of the Endophytic Mycobiota in the Roots of Rare Wild Rice (Oryza granulate) from a Nature Reserve in Yunnan, China. Appl. Environ. Microbiol. 2010, 76, 1642–1652. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Ding, Q.; Hyde, K.D.; Guo, L.D. Community Structure and Preference of Endophytic Fungi of Three Woody Plants in a Mixed Forest. Fungal Ecol. 2012, 5, 624–632. [Google Scholar] [CrossRef]
- Higgins, K.L.; Coley, P.D.; Kursar, T.A.; Arnold, A.E. Culturing and Direct PCR Suggest Prevalent Host Generalism among Diverse Fungal Endophytes of Tropical Forest Grasses. Mycologia 2011, 103, 247–260. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.; Sun, X.; He, C.; Maitra, P.; Li, X.-C.; Guo, L.-D. Phyllosphere Epiphytic and Endophytic Fungal Community and Network Structures Differ in a Tropical Mangrove Ecosystem. Microbiome 2019, 7, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huhe; Chen, X.; Hou, F.; Wu, Y.; Cheng, Y. Bacterial and Fungal Community Structures in Loess Plateau Grasslands with Different Grazing Intensities. Front. Microbiol. 2017, 8, 606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Xiong, Z.; Wu, G.; Bai, W.; Zhu, Z.; Gao, Y.; Parmar, S.; Sharma, V.K.; Li, H. Fungal Endophytic Communities of Two Wild Rosa Varieties with Different Powdery Mildew Susceptibilities. Front. Microbiol. 2018, 9, 2462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Xie, H.; Cao, L.; Zhang, R.; Xu, Z.; Wang, Z.; Deng, Z. Effects of Cd- and Pb-Resistant Endophytic Fungi on Growth and Phytoextraction of Brassica Napus in Metal-Contaminated Soils. Environ. Sci. Pollut. Res. 2017, 24, 417–426. [Google Scholar] [CrossRef]
- Pandya, M.; Rajput, M.; Rajkumar, S. Exploring Plant Growth Promoting Potential of Non Rhizobial Root Nodules Endophytes of Vigna Radiata. Microbiology 2015, 84, 80–89. [Google Scholar] [CrossRef]
- Matzrafi, M.; Preston, C.; Brunharo, C.A. Review: Evolutionary Drivers of Agricultural Adaptation in Lolium Spp. Pest Manag. Sci. 2021, 77, 2209–2218. [Google Scholar] [CrossRef]
- Parmar, S.; Sharma, V.K.; Li, T.; Tang, W.; Li, H. Fungal Seed Endophyte FZT214 Improves Dysphania Ambrosioides Cd Tolerance Throughout Different Developmental Stages. Front. Microbiol. 2022, 12, 783475. [Google Scholar] [CrossRef]
- Guevara-Araya, M.J.; Vilo, C.; Urzúa, A.; González-Teuber, M. Differences in Community Composition of Endophytic Fungi between Above- and below-Ground Tissues of Aristolochia Chilensis in an Arid Ecosystem. Rev. Chil. Hist. Nat. 2020, 93, 3. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, P.; Yang, F.; Li, M.; Yan, H.; Li, N.; Zhang, X.; Wang, W. Composition and Diversity of Endophytic Bacterial Community in Seeds of Super Hybrid Rice ‘Shenliangyou 5814’ (Oryza Sativa L.) and Its Parental Lines. Plant Growth Regul. 2019, 87, 257–266. [Google Scholar] [CrossRef]
- Ramula, S.; Mathew, S.A.; Kalske, A.; Nissinen, R.; Saikkonen, K.; Helander, M. Glyphosate Residues Alter the Microbiota of a Perennial Weed with a Minimal Indirect Impact on Plant Performance. Plant Soil 2022, 472, 161–174. [Google Scholar] [CrossRef]
- González-Teuber, M.; Vilo, C.; Bascuñán-Godoy, L. Molecular Characterization of Endophytic Fungi Associated with the Roots of Chenopodium Quinoa Inhabiting the Atacama Desert, Chile. Genom. Data 2017, 11, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Ghimire, S.R.; Charlton, N.D.; Bell, J.D.; Krishnamurthy, Y.L.; Craven, K.D. Biodiversity of Fungal Endophyte Communities Inhabiting Switchgrass (Panicum virgatum L.) Growing in the Native Tallgrass Prairie of Northern Oklahoma. Fungal Divers. 2011, 47, 19–27. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, Q.; Ju, M.; Yan, S.; Zhang, Q.; Gu, P. The Endophytic Fungi Diversity, Community Structure, and Ecological Function Prediction of Sophora Alopecuroides in Ningxia, China. Microorganisms 2022, 10, 2099. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.-Y.; Guo, L.-D.; Hyde, K.D. Response of Endophytic Fungi of Stipa Grandis to Experimental Plant Function Group Removal in Inner Mongolia Steppe, China. Fungal Divers. 2010, 43, 93–101. [Google Scholar] [CrossRef]
- Pang, B.; Yin, D.; Zhai, Y.; He, A.; Qiu, L.; Liu, Q.; Ma, N.; Shen, H.; Jia, Q.; Liang, Z.; et al. Diversity of Endophytic Fungal Community in Huperzia Serrata from Different Ecological Areas and Their Correlation with Hup A Content. BMC Microbiol. 2022, 22, 191. [Google Scholar] [CrossRef] [PubMed]
- Alfiky, A.; Weisskopf, L. Deciphering Trichoderma–Plant–Pathogen Interactions for Better Development of Biocontrol Applications. J. Fungi 2021, 7, 61. [Google Scholar] [CrossRef]
- Terhonen, E.; Blumenstein, K.; Kovalchuk, A.; Asiegbu, F.O. Forest Tree Microbiomes and Associated Fungal Endophytes: Functional Roles and Impact on Forest Health. Forests 2019, 10, 42. [Google Scholar] [CrossRef] [Green Version]
- Ye, D.; Li, T.; Yi, Y.; Zhang, X.; Zou, L. Characteristics of Endophytic Fungi from Polygonum Hydropiper Suggest Potential Application for P-Phytoextraction. Fungal Ecol. 2019, 41, 126–136. [Google Scholar] [CrossRef]
- Baron, N.C.; Rigobelo, E.C. Endophytic Fungi: A Tool for Plant Growth Promotion and Sustainable Agriculture. Mycology 2022, 13, 39–55. [Google Scholar] [CrossRef]
- Argumedo-Delira, R.; Gómez-Martínez, M.J.; Mora-Delgado, J. Plant Growth Promoting Filamentous Fungi and Their Application in the Fertilization of Pastures for Animal Consumption. Agronomy 2022, 12, 3033. [Google Scholar] [CrossRef]
- Mundim, G.d.S.M.; Maciel, G.M.; Mendes, G.d.O. Aspergillus Niger as a Biological Input for Improving Vegetable Seedling Production. Microorganisms 2022, 10, 674. [Google Scholar] [CrossRef] [PubMed]
- Sugiharto, A. Response of Growth of Garlic Towards Aspergillus Niger and Fusarium Sp. Inoculant. IOP Conf. Ser. Earth Environ. Sci. 2019, 308, 012058. [Google Scholar] [CrossRef]
- Araújo, V.C.; Rossati, K.F.; Xavier, L.V.; Oliveira, V.A.d.; Carmo, G.J.d.S.; Assis, G.A.d.; Mendes, G.d.O. Enhanced Growth in Nursery of Coffee Seedlings Inoculated with the Rhizosphere Fungus Aspergillus Niger for Field Transplantation. Rhizosphere 2020, 15, 100236. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, Q.; Mao, W.; Tang, W.; White, J.F.; Li, H. Endophytic Bacterial Community of Stellera Chamaejasme L. and Its Role in Improving Host Plants’ Competitiveness in Grasslands. Environ. Microbiol. 2022, 24, 3322–3333. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Morphotype | Solubilized Phosphate (mg/L) | IAA (mg/L) | SU% |
|---|---|---|---|---|
| STL3G74 | Aspergillus niger | 296.33 | 208.42 | 87.64 |
| LT2S21 | Gibberella sp. | 109.30 | 124.28 | - |
| STL1G60 | Ilyonectria robusta | 195.67 | 136.96 | - |
| LT2G30 | Trametes sp. | 94.27 | - | 80.74 |
| LT2S36 | Phaeosphaeriaceae sp. | 141.1 | - | 80.36 |
| LT1G13 | Porostereum spadiceum | 230.18 | - | - |
| STL3G8 | Plenodomus tracheiphilus | 292.78 | - | 56.31 |
| LT3G35 | Rhexocercosporidium sp. | 135.03 | 32.05 | 62.56 |
| Sample | No. of Segments Plated (No. of Segments Colonized by EF) | No. of EF Isolated | CR (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| G | S | Total | G | S | Total | G | S | Total | |
| Ⅰ | 188 (70) | 195 (42) | 383 (112) | 125 | 67 | 192 | 37.23 | 21.54 | 29.24 |
| Ⅱ | 180 (53) | 195 (50) | 375 (103) | 110 | 81 | 191 | 29.44 | 25.64 | 27.47 |
| Ⅲ | 181 (60) | 175 (28) | 373 (80) | 113 | 50 | 163 | 33.15 | 16 | 21.45 |
| Average | - | 33.27 ± 3.9 a | 22.39 ± 2.92 b | 30.16 ± 3.25 | |||||
| Total | 549 (183) | 565 (120) | 1114 (303) | 348 | 198 | 546 | - | ||
| Tissue | Sample ID | Diversity Indices of EF | ||||
|---|---|---|---|---|---|---|
| Taxa (S) | H′ | Evenness | Chao1 | D | ||
| Shoot | S1 | 14 | 2.29 | 0.87 | 15.13 | 0.87 |
| S2 | 15 | 2.28 | 0.84 | 21.25 | 0.87 | |
| S3 | 10 | 1.81 | 0.78 | 10.25 | 0.78 | |
| Average | 13 ± 2.65 a | 2.13 ± 0.28 a | 0.83 ± 0.04 a | 15.54 ± 5.51 a | 0.84 ± 0.06 a | |
| Root | G1 | 21 | 2.78 | 0.91 | 25.00 | 0.93 |
| G2 | 17 | 2.37 | 0.84 | 19.67 | 0.88 | |
| G3 | 21 | 2.50 | 0.82 | 30.00 | 0.89 | |
| Average | 19.67 ± 2.31 b | 2.55 ± 0.21 a | 0.86 ± 0.05 a | 24.89 ± 5.17 a | 0.90 ± 0.03 a | |
| Tissue | Sample ID | Number of Sequences | OTU | α-Diversity | |||
|---|---|---|---|---|---|---|---|
| Simpson | Shannon | ace | Chao | ||||
| Shoot | S1 | 119,019 | 82 | 0.30 | 1.733 | 25.75 | 82 |
| S2 | 124,023 | 0.38 | 1.715 | 35 | 68 | ||
| S3 | 109,998 | 0.21 | 2.207 | 37 | 108.45 | ||
| Average | 117,680 | 0.30 | 1.885 | 32.59 | 86.15 | ||
| Root | G1 | 98,814 | 201 | 0.43 | 1.143 | 82 | 82 |
| G2 | 53,288 | 0.30 | 1.663 | 68 | 68 | ||
| G3 | 109,136 | 0.24 | 2.036 | 108.45 | 108.5 | ||
| Average | 87,079 | 0.32 | 1.61 | 86.15 | 86.17 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, W.; Gong, W.; Xiao, R.; Mao, W.; Zhao, L.; Song, J.; Awais, M.; Ji, X.; Li, H. Endophytic Fungal Community of Stellera chamaejasme L. and Its Possible Role in Improving Host Plants’ Ecological Flexibility in Degraded Grasslands. J. Fungi 2023, 9, 465. https://doi.org/10.3390/jof9040465
Tang W, Gong W, Xiao R, Mao W, Zhao L, Song J, Awais M, Ji X, Li H. Endophytic Fungal Community of Stellera chamaejasme L. and Its Possible Role in Improving Host Plants’ Ecological Flexibility in Degraded Grasslands. Journal of Fungi. 2023; 9(4):465. https://doi.org/10.3390/jof9040465
Chicago/Turabian StyleTang, Wenting, Weijun Gong, Ruitong Xiao, Wenqin Mao, Liangzhou Zhao, Jinzhao Song, Muhammad Awais, Xiuling Ji, and Haiyan Li. 2023. "Endophytic Fungal Community of Stellera chamaejasme L. and Its Possible Role in Improving Host Plants’ Ecological Flexibility in Degraded Grasslands" Journal of Fungi 9, no. 4: 465. https://doi.org/10.3390/jof9040465