

Impact of Fusarium Infection on Potato Quality, Starch Digestibility, In Vitro Glycemic Response, and Resistant Starch Content

 , ,
, ,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Maintenance of Fungal Pure Culture and Potato Cultivars
2.2. Morphological and Molecular Identification of the Fungus
2.3. Tuber Inoculation and Estimation of Lesion Diameter, Lesion Depth, and Rot Volume
2.3.1. Inoculum Preparation
2.3.2. Tuber Inoculation
2.3.3. Periodic Disease Assessment
2.4. Tuber Processing for Biochemical Analysis
2.4.1. Determination of Total Starch Content
2.4.2. Determination of Amylose Content
2.5. Determination of Glycemic Index (GI), Glycemic Load (GL), and Resistant Starch (RS)
2.5.1. Estimation of In Vitro GI
2.5.2. Estimation of In Vitro GL
2.5.3. Determination of RS
2.6. Statistical Analysis
3. Results
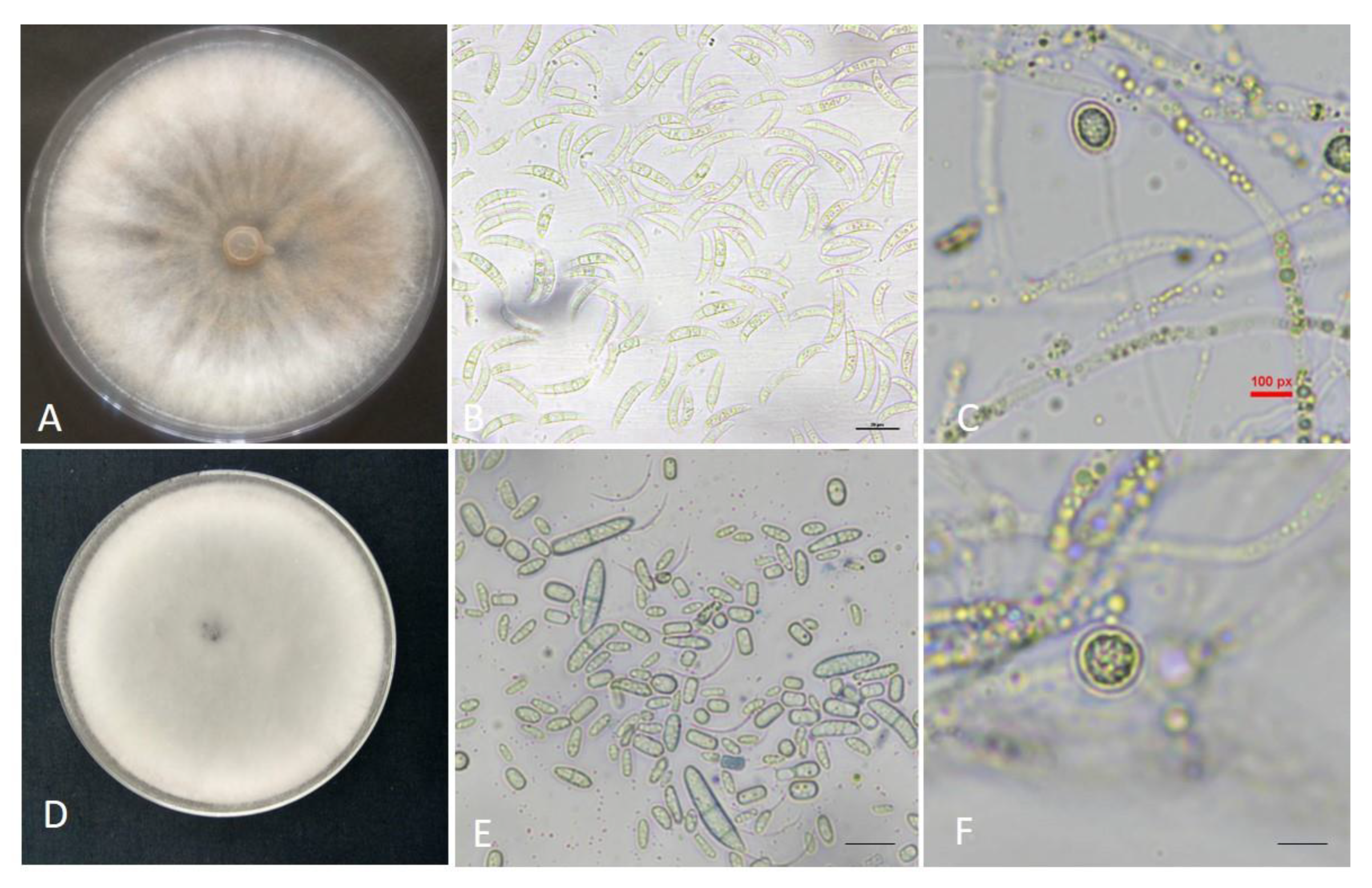
3.1. Morphological Identification of Fungal Isolates
3.2. Molecular Identification and Phylogenetic Analysis
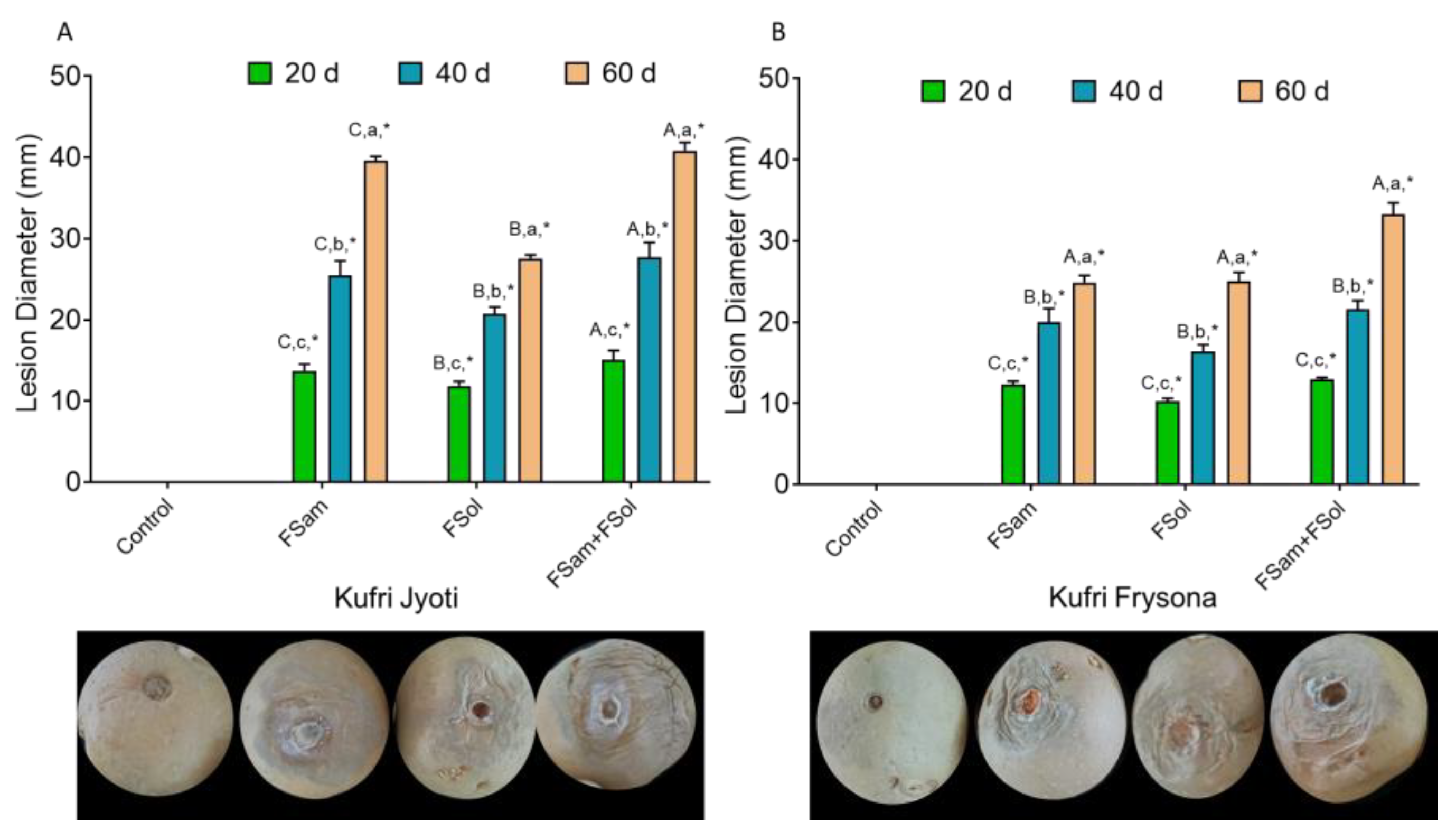
3.3. Effect of Dry Rot Infection on Lesion Diameter
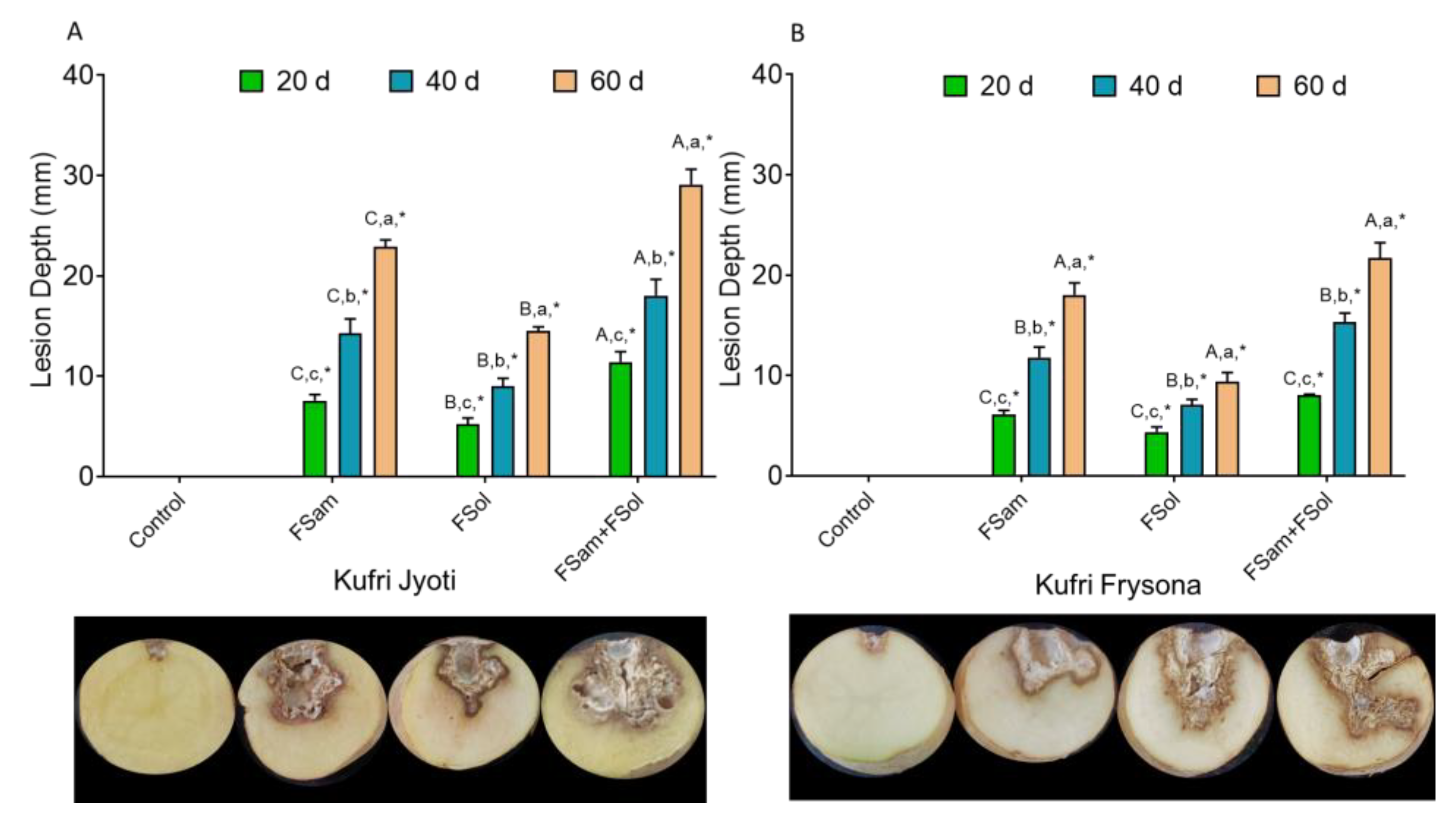
3.4. Estimation of Lesion Depth in Inoculated Tubers
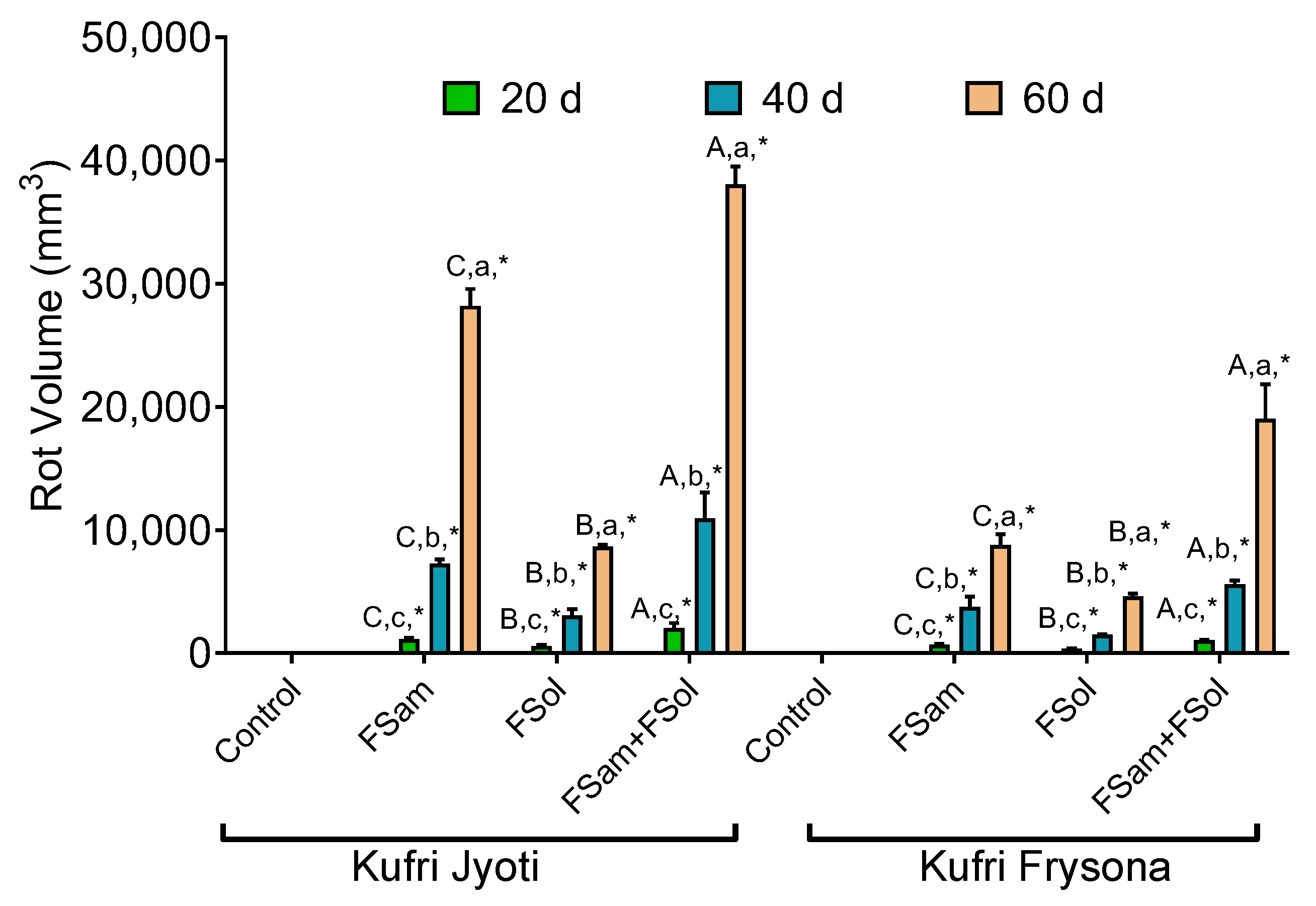
3.5. Association of Fusarium Infection and Rot Volume
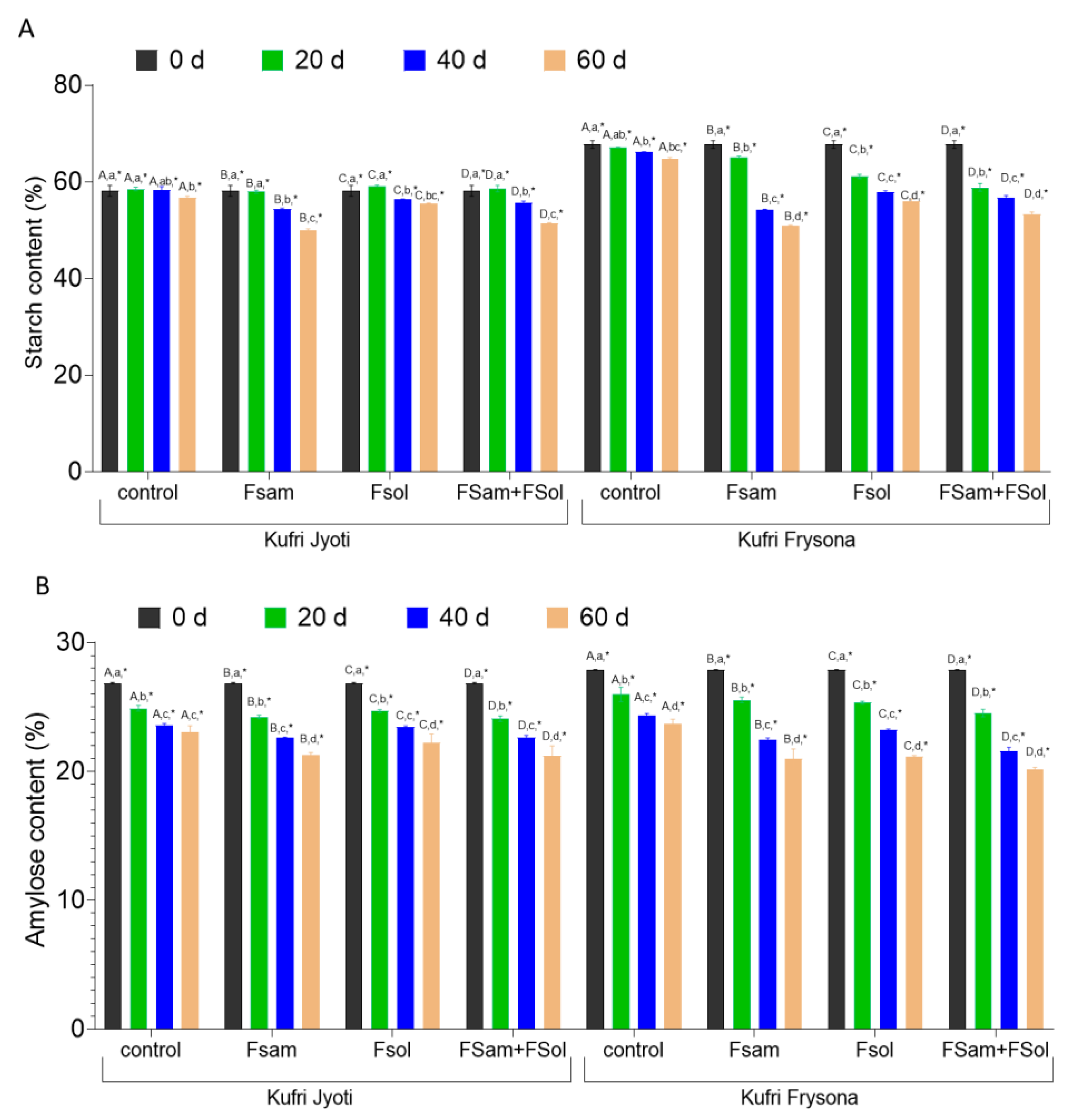
3.6. Starch and Amylose Content in Dry Rot Infected Tubers
3.7. Dry Rot Infection Affects Glycemic Index (GI), Resistant Starch (RS), and Glycemic Load (GL) of Potato Tubers
3.8. Pearson Correlation Analysis between the Parameters of Dry Rot Infection and Glycemic Response Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Devaux, A.; Goffart, J.P.; Petsakos, A.; Kromann, P.; Gatto, M.; Okello, J.; Suarez, V.; Hareau, G. Global Food Security, Contributions from Sustainable Potato Agri-Food Systems. In The Potato Crop: Its Agricultural, Nutritional and Social Contribution to Humankind; Springer International Publishing: Cham, Switzerland, 2019; pp. 3–35. ISBN 9783030286835. [Google Scholar]
- Muthoni, J.; Shimelis, H.; Crop, J.M.-A.J. Production and availability of good quality seed potatoes in the East African region: A review. Aust. J. Crop Sci. 2022, 16, 907–915. [Google Scholar] [CrossRef]
- Tiwari, R.K.; Kumar, R.; Sharma, S.; Naga, K.C.; Subhash, S.; Sagar, V. Continuous and Emerging Challenges of Silver Scurf Disease in Potato. Int. J. Pest Manag. 2020, 68, 89–101. [Google Scholar] [CrossRef]
- Larkin, R.P.; Lynch, R.P. Use and Effects of Different Brassica and Other Rotation Crops on Soilborne Diseases and Yield of Potato. Horticulturae 2018, 4, 37. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, R.K.; Kumar, R.; Sharma, S.; Sagar, V.; Aggarwal, R.; Naga, K.C.; Lal, M.K.; Chourasia, K.N.; Kumar, D.; Kumar, M. Potato Dry Rot Disease: Current Status, Pathogenomics and Management. 3 Biotech 2020, 10, 503. [Google Scholar] [CrossRef] [PubMed]
- Cullen, D.W.; Toth, I.K.; Pitkin, Y.; Boonham, N.; Walsh, K.; Barker, I.; Lees, A.K. Use of Quantitative Molecular Diagnostic Assays to Investigate Fusarium Dry Rot in Potato Stocks and Soil. Phytopathology 2005, 95, 1462–1471. [Google Scholar] [CrossRef] [Green Version]
- Azil, N.; Stefańczyk, E.; Sobkowiak, S.; Chihat, S.; Boureghda, H.; Śliwka, J. Identification and Pathogenicity of Fusarium Spp. Associated with Tuber Dry Rot and Wilt of Potato in Algeria. Eur. J. Plant Pathol. 2021, 159, 495–509. [Google Scholar] [CrossRef]
- Ammar, N.; Nefzi, A.; Jabnoun-Khiareddine, H.; Daami-Remadi, M. Control of Fusarium Dry Rot Incited by Fusarium oxysporum f. Sp. Tuberosi Using Sargassum vulgare Aqueous and Organic Extracts. J. Microb. Biochem. Technol. 2017, 9, 200–208. [Google Scholar] [CrossRef]
- Peters, J.C.; Lees, A.K.; Cullen, D.W.; Sullivan, L.; Stroud, G.P.; Cunnington, A.C. Characterization of Fusarium spp. Responsible for Causing Dry Rot of Potato in Great Britain. Plant Pathol. 2008, 57, 262–271. [Google Scholar] [CrossRef]
- Tiwari, R.K.; Bashyal, B.M.; Shanmugam, V.; Lal, M.K.; Kumar, R.; Sharma, S.; Naga, K.C.; Chourasia, K.N.; Aggarwal, R. First Report of Dry Rot of Potato Caused by Fusarium proliferatum in India. J. Plant Dis. Prot. 2022, 129, 173–179. [Google Scholar] [CrossRef]
- Masum, M.M.I.; Islam, S.M.M.; Islam, M.S.; Kabir, M.H. Estimation of Loss Due to Post Harvest Diseases of Potato in Markets of Different Districts in Bangladesh. Afr. J. Biotechnol. 2011, 10, 11892–11902. [Google Scholar] [CrossRef]
- Chen, D.; Nahar, K.; Bizimungu, B.; Soucy, S.; Peters, R.D.; De Koeyer, D.; Dickison, V. A Simple and Efficient Inoculation Method for Fusarium Dry Rot Evaluations in Potatoes. Am. J. Potato Res. 2020, 97, 265–271. [Google Scholar] [CrossRef]
- Du, M.; Ren, X.; Sun, Q.; Wang, Y.; Zhang, R. Characterization of Fusarium Spp. Causing Potato Dry Rot in China and Susceptibility Evaluation of Chinese Potato Germplasm to the Pathogen. Potato Res. 2012, 55, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Merlington, A.; Hanson, L.E.; Bayma, R.; Hildebrandt, K.; Steere, L.; Kirk, W.W. First Report of Fusarium proliferatum Causing Dry Rot in Michigan Commercial Potato (Solanum tuberosum) production. Plant Dis. 2014, 98, 843. [Google Scholar] [CrossRef]
- Sagar, V.; Sharma, S.; Jeevalatha, A.; Chakrabarti, S.K.; Singh, B.P. First Report of Fusarium Sambucinum Causing Dry Rot of Potato in India. New Dis. Rep. 2011, 24, 5. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, R.K.; Bashyal, B.M.; Shanmugam, V.; Lal, M.K.; Kumar, R.; Sharma, S.; Vinod; Gaikwad, K.; Singh, B.; Aggarwal, R. Impact of Fusarium Dry Rot on Physicochemical Attributes of Potato Tubers during Postharvest Storage. Postharvest Biol. Technol. 2021, 181, 111638. [Google Scholar] [CrossRef]
- Lastochkina, O.; Pusenkova, L.; Garshina, D.; Yuldashev, R.; Shpirnaya, I.; Kasnak, C.; Palamutoglu, R.; Mardanshin, I.; Garipova, S.; Sobhani, M.; et al. The Effect of Endophytic Bacteria Bacillus subtilis and Salicylic Acid on Some Resistance and Quality Traits of Stored Solanum tuberosum L. Tubers Infected with Fusarium Dry Rot. Plants 2020, 9, 738. [Google Scholar] [CrossRef]
- Sarian, F.D.; Van Der Kaaij, R.M.; Kralj, S.; Wijbenga, D.J.; Binnema, D.J.; Van Der Maarel, M.J.E.C.; Dijkhuizen, L. Enzymatic Degradation of Granular Potato Starch by Microbacterium aurum Strain B8.A. Appl. Microbiol. Biotechnol. 2012, 93, 645–654. [Google Scholar] [CrossRef] [Green Version]
- Torres, M.D.Á.; Parreño, W.C. Thermal Processing of Potatoes. In Advances in Potato Chemistry and Technology, 2nd ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 403–457. ISBN 9780128000021. [Google Scholar]
- Lal, M.K.; Singh, B.; Tiwari, R.K.; Kumar, S.; Gopalakrishnan, S.; Gaikwad, K.; Kumar, A.; Paul, V.; Singh, M.P. Interactive Effect of Retrogradation and Addition of Pulses, Cooking Oil on Predicted Glycemic Index and Resistant Starch of Potato. Starch-Starke 2022, 74, 2100221. [Google Scholar] [CrossRef]
- Bandana; Sharma, V.; Singh, B.; Raigond, P.; Kaushik, S.K. Role of invertase Activity in Processing Quality of potatoes: Effect of Storage Temperature and Duration. J. Environ. Biol. 2016, 37, 239–245. [Google Scholar]
- Singh, A.; Raigond, P.; Lal, M.K.; Singh, B.; Thakur, N.; Changan, S.S.; Kumar, D.; Dutt, S. Effect of Cooking Methods on Glycemic Index and in Vitro Bioaccessibility of Potato (Solanum tuberosum L.) Carbohydrates. LWT 2020, 127, 109363. [Google Scholar] [CrossRef]
- Raigond, P.; Parmar, V.; Thakur, A.; Lal, M.K.; Changan, S.S.; Kumar, D.; Dutt, S.; Singh, B. Composition of Different Carbohydrate Fractions in Potatoes: Effect of Cooking and Cooling. Starch-Stärke 2021, 73, 2100015. [Google Scholar] [CrossRef]
- Herman, D.J.; Knowles, L.O.; Knowles, N.R. Heat Stress Affects Carbohydrate Metabolism during Cold-Induced Sweetening of Potato (Solanum tuberosum L.). Planta 2017, 245, 563–582. [Google Scholar] [CrossRef]
- Murniece, I.; Karklina, D.; Galoburda, R.; Sabovics, M. Reducing Sugar Content and Colour Intensity of Fried Latvian Potato. LLU Raksti 2010, 24, 20–30. [Google Scholar]
- Novy, R.G.; Whitworth, J.L.; Stark, J.C.; Charlton, B.A.; Yilma, S.; Knowles, N.R.; Pavek, M.J.; Spear, R.R.; Brandt, T.L.; Olsen, N.; et al. Teton Russet: An Early-Maturing, Dual-Purpose Potato Cultivar Having Higher Protein and Vitamin C Content, Low Asparagine, and Resistances to Common Scab and Fusarium Dry Rot. Am. J. Potato Res. 2014, 91, 380–393. [Google Scholar] [CrossRef]
- Lal, M.K.; Singh, B.; Sharma, S.; Singh, M.P.; Kumar, A. Glycemic Index of Starchy Crops and Factors Affecting Its Digestibility: A Review. Trends Food Sci. Technol. 2021, 111, 741–755. [Google Scholar] [CrossRef]
- Lal, M.K.; Kumar, A.; Raigond, P.; Dutt, S.; Changan, S.S.; Chourasia, K.N.; Tiwari, R.K.; Kumar, D.; Sharma, S.; Chakrabarti, S.K.; et al. Impact of Starch Storage Condition on Glycemic Index and Resistant Starch of Cooked Potato (Solanum tuberosum) Tubers. Starch-Staerke 2021, 73, 1900281. [Google Scholar] [CrossRef]
- Kumar, A.; Dash, G.K.; Barik, M.; Panda, P.A.; Lal, M.K.; Baig, M.J.; Swain, P. Effect of Drought Stress on Resistant Starch Content and Glycemic Index of Rice (Oryza sativa L.). Starch-Staerke 2020, 72, 1900229. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Hoboken, NJ, USA, 2007; ISBN 0813819199. [Google Scholar]
- Heltoft, P.; Molteberg, E.L.; Nærstad, R.; Hermansen, A. Effect of Maturity Level and Potato Cultivar on Development of Fusarium Dry Rot in Norway. Potato Res. 2015, 58, 205–219. [Google Scholar] [CrossRef]
- McCready, R.M.; Guggolz, J.; Silviera, V.; Owens, H.S. Determination of Starch and Amylose in Vegetables. Anal. Chem. 1950, 22, 1156–1158. [Google Scholar] [CrossRef]
- Juliano, B.O.; Perez, C.M.; Blakeney, A.B.; Castillo, T.; Kongseree, N.; Laignelet, B.; Lapis, E.T.; Murty, V.V.S.; Paule, C.M.; Webb, B.D. International Cooperative Testing on the Amylose Content of Milled Rice. Starch-Stärke 1981, 33, 157–162. [Google Scholar] [CrossRef]
- Goñi, I.; Garcia-Alonso, A.; Saura-Calixto, F. A Starch Hydrolysis Procedure to Estimate Glycemic Index. Nutr. Res. 1997, 17, 427–437. [Google Scholar] [CrossRef]
- Goñi, I.; García-Diz, L.; Mañas, E.; Saura-Calixto, F. Analysis of Resistant Starch: A Method for Foods and Food Products. Food Chem. 1996, 56, 445–449. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Sahoo, U.; Baisakha, B.; Okpani, O.A.; Ngangkham, U.; Parameswaran, C.; Basak, N.; Kumar, G.; Sharma, S.G. Resistant Starch Could Be Decisive in Determining the Glycemic Index of Rice Cultivars. J. Cereal Sci. 2018, 79, 348–353. [Google Scholar] [CrossRef]
- Leslie, J.; Summerell, B. The Fusarium Laboratory Manual, 1st ed.; Blackwell Publishing Professional: New York, NY, USA, 2006; pp. 8–240. [Google Scholar]
- Mejdoub Trabelsi, B.; Khiareddine, H.J.; Remadi, M.D. Interactions between Four Fusarium Species in Potato Tubers and Consequences for Fungal Development and Susceptibility Assessment of Five Potato Cultivars under Different Storage Temperature. J. Plant Pathol. Microbiol. 2015, 6, 293. [Google Scholar] [CrossRef]
- Heltoft, P.; Brierley, J.L.; Lees, A.K.; Sullivan, L.; Lynott, J.; Hermansen, A. The relationship between soil inoculum and the development of Fusarium dry rot in potato cultivars Asterix and Saturna. Eur. J. Plant Pathol. 2016, 146, 711–714. [Google Scholar] [CrossRef]
- Fang, X.; Zhang, C.; Wang, Z.; Duan, T.; Yu, B.; Jia, X.; Pang, J.; Ma, L.; Wang, Y.; Nan, Z. Co-Infection by Soil-Borne Fungal Pathogens Alters Disease Responses among Diverse Alfalfa Varieties. Front. Microbiol. 2021, 12, 1829. [Google Scholar] [CrossRef] [PubMed]
- Machado, A.K.; Brown, N.A.; Urban, M.; Kanyuka, K.; Hammond-Kosack, K.E. RNAi as an Emerging Approach to Control Fusarium Head Blight Disease and Mycotoxin Contamination in Cereals. Pest Manag. Sci. 2018, 74, 790–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arias, M.M.D.; Leandro, L.F.; Munkvold, G.P. Aggressiveness of Fusarium Species and Impact of Root Infection on Growth and Yield of Soybeans. Ecol. Epidemiol. 2013, 103, 822–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzdraliński, A.; Szczerba, H.; Tofil, K.; Filipiak, A.; Garbarczyk, E.; Dziadko, P.; Muszyńska, M.; Solarska, E. Early PCR-Based Detection of Fusarium culmorum, F. Graminearum, F. Sporotrichioides and F. poae on Stem Bases of Winter Wheat throughout Poland. Eur. J. Plant Pathol. 2014, 140, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Lal, M.K.; Tiwari, R.K.; Kumar, R.; Naga, K.C.; Kumar, A.; Singh, B.; Raigond, P.; Dutt, S.; Chourasia, K.N.; Kumar, D.; et al. Effect of Potato Apical Leaf Curl Disease on Glycemic Index and Resistant Starch of Potato (Solanum tuberosum L.) Tubers. Food Chem. 2021, 359, 129939. [Google Scholar] [CrossRef]
- Kumar, A.; Sahoo, U.; Lal, M.K.; Tiwari, R.K.; Lenka, S.K.; Singh, N.R.; Gupta, O.P.; Sah, R.P.; Sharma, S. Biochemical Markers for Low Glycemic Index and Approaches to Alter Starch Digestibility in Rice. J. Cereal Sci. 2022, 106, 103501. [Google Scholar] [CrossRef]
- Lal, M.K.; Tiwari, R.K.; Jaiswal, A.; Luthra, S.K.; Singh, B.; Kumar, S.; Gopalakrishnan, S.; Gaikwad, K.; Kumar, A.; Paul, V.; et al. Combinatorial Interactive Effect of Vegetable and Condiments with Potato on Starch Digestibility and Estimated in Vitro Glycemic Response. J. Food Meas. Charact. 2022, 16, 2446–2458. [Google Scholar] [CrossRef]
- Lal, M.K.; Sharma, N.; Adavi, S.B.; Sharma, E.; Altaf, M.A.; Tiwari, R.K.; Kumar, R.; Kumar, A.; Dey, A.; Paul, V.; et al. From Source to Sink: Mechanistic Insight of Photoassimilates Synthesis and Partitioning under High Temperature and Elevated [CO2]. Plant Mol. Biol. 2022, 1, 305–324. [Google Scholar] [CrossRef]
- Kapinga, R.; Ndunguru, J.; Mulokozi, G.; Tumwegamire, S. Impact of Common Sweetpotato Viruses on Total Carotenoids and Root Yields of an Orange-Fleshed Sweetpotato in Tanzania. Sci. Hortic. 2009, 122, 1–5. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Lesion Diameter | Lesion Depth | Rot Volume | Starch | Amylose | Glycemic Index | Glycemic Load | Resistant Starch |
|---|---|---|---|---|---|---|---|---|
| Lesion Diameter | 1.00 | |||||||
| Lesion Depth | 0.96 | 1.00 | ||||||
| Rot Volume | 0.82 | 0.87 | 1.00 | |||||
| Starch | −0.79 | −0.78 | −0.64 | 1.00 | ||||
| Amylose | −0.92 | −0.88 | −0.64 | 0.82 | 1.00 | |||
| Glycemic Index | 0.30 | 0.31 | 0.15 | −0.03 | −0.41 | 1.00 | ||
| Glycemic Load | 0.56 | 0.59 | 0.40 | −0.30 | −0.63 | 0.92 | 1.00 | |
| Resistant Starch | −0.60 | −0.59 | −0.41 | 0.29 | 0.65 | −0.91 | −0.96 | 1.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiwari, R.K.; Lal, M.K.; Kumar, R.; Sharma, S.; Sagar, V.; Kumar, A.; Singh, B.; Aggarwal, R. Impact of Fusarium Infection on Potato Quality, Starch Digestibility, In Vitro Glycemic Response, and Resistant Starch Content. J. Fungi 2023, 9, 466. https://doi.org/10.3390/jof9040466
Tiwari RK, Lal MK, Kumar R, Sharma S, Sagar V, Kumar A, Singh B, Aggarwal R. Impact of Fusarium Infection on Potato Quality, Starch Digestibility, In Vitro Glycemic Response, and Resistant Starch Content. Journal of Fungi. 2023; 9(4):466. https://doi.org/10.3390/jof9040466
Chicago/Turabian StyleTiwari, Rahul Kumar, Milan Kumar Lal, Ravinder Kumar, Sanjeev Sharma, Vinay Sagar, Awadhesh Kumar, Brajesh Singh, and Rashmi Aggarwal. 2023. "Impact of Fusarium Infection on Potato Quality, Starch Digestibility, In Vitro Glycemic Response, and Resistant Starch Content" Journal of Fungi 9, no. 4: 466. https://doi.org/10.3390/jof9040466