
α-Pheromone Precursor Protein Foc4-PP1 Is Essential for the Full Virulence of Fusarium oxysporum f. sp. cubense Tropical Race 4


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Fungal Strains
2.2. CDS Amplification and Sequence Analysis
2.3. Generation of Foc4-PP1 Deletion Mutants
2.4. Pathogenicity Assays
2.5. Growth, Stress Sensitivity, and Cellophane Membrane Assays
2.6. Subcellular Localization and Cell Death Suppression Assay
3. Results
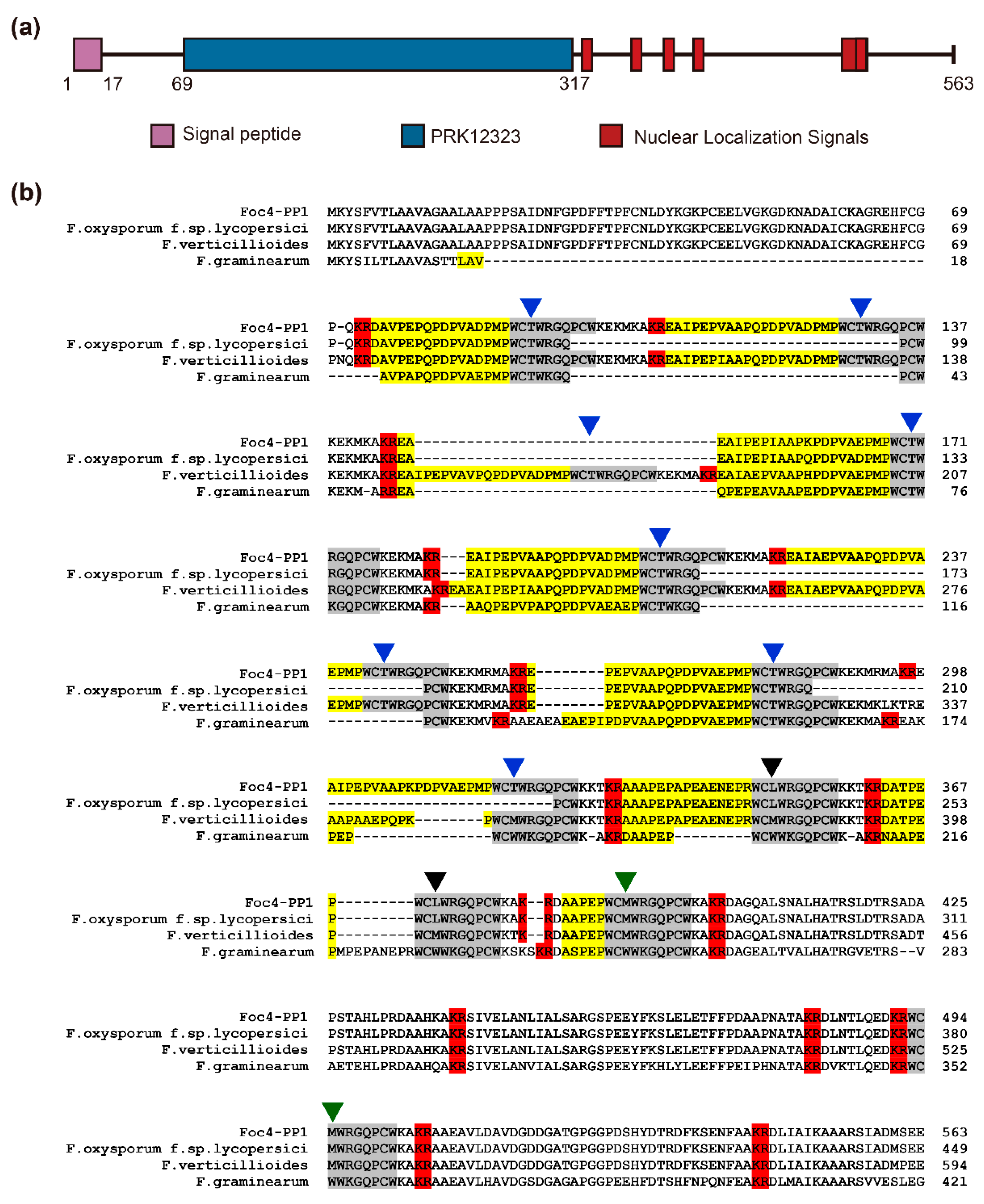
3.1. Sequence Analysis of the Foc4 PP1 Gene and Protein
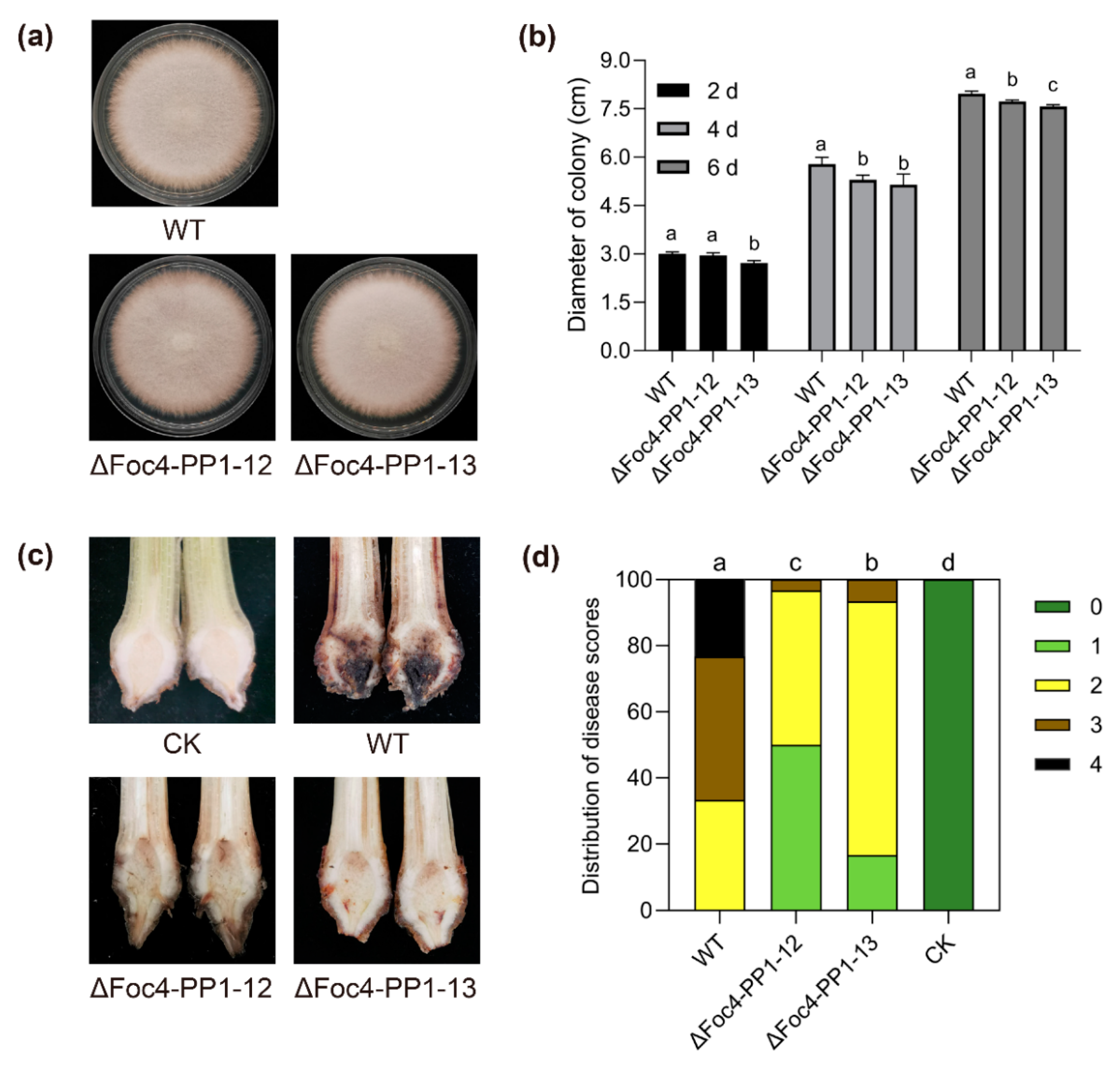
3.2. Deletion of Foc4-PP1 Compromises Fungal Growth and the Virulence of Foc TR4 14013 in Cavendish Banana
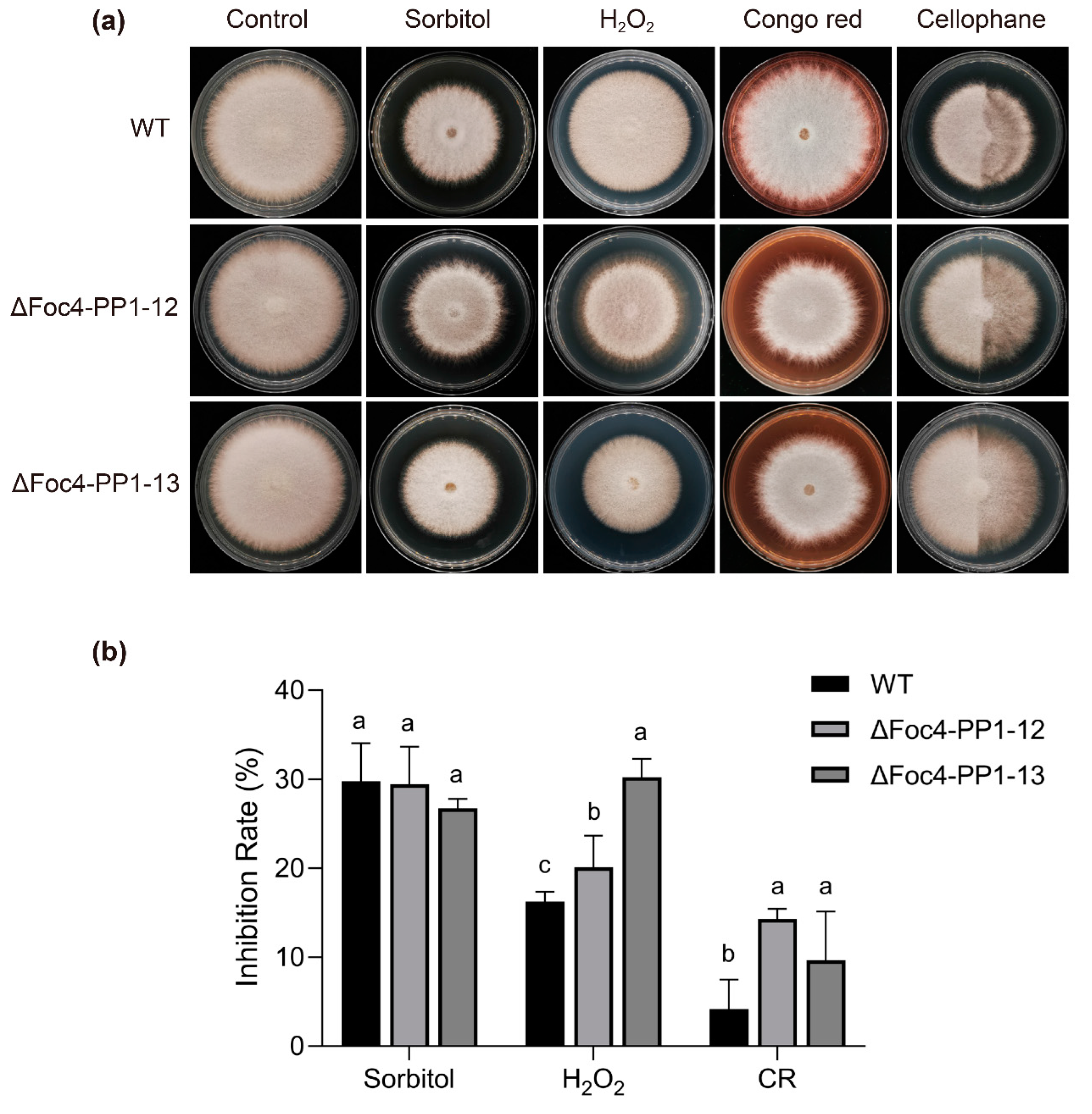
3.3. Sensitivity of the Foc4-PP1 Deletion Mutants to Abiotic Stresses
3.4. Foc4-PP1-Mediated Suppression of BAX-Induced Cell Death in N. benthamiana
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ploetz, R.C. Management of Fusarium wilt of banana: A review with special reference to tropical race 4. Crop Prot. 2015, 73, 7–15. [Google Scholar] [CrossRef]
- Ploetz, R.C.; Correll, J.C. Vegetative compatibility among races of Fusarium oxysporum f. sp. cubense. Plant Dis. 1988, 72, 325–328. [Google Scholar] [CrossRef]
- Ploetz, R.; Pegg, K. Fungal diseases of root, corm and pseudostem. In Handbook of Diseases of Banana, Abacá and Enset; CAB international: Wallingford, UK, 2000; pp. 143–159. [Google Scholar]
- Waite, B.H. Wilt of Heliconia spp. caused by Fusarium oxysporum f. sp. cubense race 3. Trop. Agric. 1963, 40, 299–305. [Google Scholar]
- Hwang, S.-C.; Ko, W.-H. Cavendish Banana Cultivars Resistant to Fusarium Wilt Acquired through Somaclonal Variation in Taiwan. Plant Dis. 2004, 88, 580–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuguang, W.; Qiyu, X.; Guihua, W.; He, Z.; Xuehua, L.; Jianbo, S.; Xin, Z. Differential gene expression in banana roots in response to Fusarium wilt. Can. J. Plant Pathol. 2017, 39, 163–175. [Google Scholar]
- Buddenhagen, I. Understanding strain diversity in Fusarium oxysporum f. sp. cubense and history of introduction of ‘tropical race 4′ to better manage banana production. In III International Symposium on Banana: ISHS-ProMusa Symposium on Recent Advances in Banana Crop Protection for Sustainable 828; ISHS: Leuven, Belgium, 2009; pp. 193–204. [Google Scholar]
- Acuña, R.; Rouard, M.; Leiva, A.M.; Marques, C.; Olortegui, J.A.; Ureta, C.; Cabrera-Pintado, R.M.; Rojas, J.C.; Lopez-Alvarez, D.; Cenci, A.; et al. First Report of Fusarium oxysporum f. sp. cubense Tropical Race 4 Causing Fusarium Wilt in Cavendish Bananas in Peru. Plant Dis. 2022, 106, 2268. [Google Scholar] [CrossRef]
- Ordonez, N.; Seidl, M.F.; Waalwijk, C.; Drenth, A.; Kilian, A.; Thomma, B.P.H.J.; Ploetz, R.C.; Kema, G.H.J. Worse Comes to Worst: Bananas and Panama Disease—When Plant and Pathogen Clones Meet. PLoS Pathog. 2015, 11, e1005197. [Google Scholar] [CrossRef] [Green Version]
- Dong, Z.; Luo, M.; Wang, Z. An Exo-Polygalacturonase Pgc4 Regulates Aerial Hyphal Growth and Virulence in Fusarium oxysporum f. sp. cubense race 4. Int. J. Mol. Sci. 2020, 21, 5886. [Google Scholar] [CrossRef]
- Ding, Z.; Yang, L.; Wang, G.; Guo, L.; Liu, L.; Wang, J.; Huang, J. Fusaric acid is a virulence factor of Fusarium oxysporum f. sp. cubense on banana plantlets. Trop. Plant Pathol. 2018, 43, 297–305. [Google Scholar] [CrossRef]
- Guo, L.; Wang, J.; Liang, C.; Yang, L.; Zhou, Y.; Liu, L.; Huang, J. Fosp9, a Novel Secreted Protein, Is Essential for the Full Virulence of Fusarium oxysporum f. sp. cubense on Banana (Musa spp.). Appl. Environ. Microbiol. 2022, 88, e00604-21. [Google Scholar] [CrossRef] [PubMed]
- Widinugraheni, S.; Niño-Sánchez, J.; Does, H.C.v.d.; Dam, P.v.; García-Bastidas, F.A.; Subandiyah, S.; Meijer, H.J.G.; Kistler, H.C.; Kema, G.H.J.; Rep, M. A SIX1 homolog in Fusarium oxysporum f.sp. cubense tropical race 4 contributes to virulence towards Cavendish banana. PLoS ONE 2018, 13, e0205896. [Google Scholar]
- An, B.; Hou, X.; Guo, Y.; Zhao, S.; Luo, H.; He, C.; Wang, Q. The effector SIX8 is required for virulence of Fusarium oxysporum f.sp. cubense tropical race 4 to Cavendish banana. Fungal Biol. 2019, 123, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wu, B.; Yang, J.; Bi, F.; Dong, T.; Yang, Q.; Hu, C.; Xiang, D.; Chen, H.; Huang, H.; et al. A Cerato-Platanin Family Protein FocCP1 Is Essential for the Penetration and Virulence of Fusarium oxysporum f. sp. cubense Tropical Race 4. Int. J. Mol. Sci. 2019, 20, 3785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Huang, H.; Wu, B.; Xie, J.; Viljoen, A.; Wang, W.; Mostert, D.; Xie, Y.; Fu, G.; Xiang, D.; et al. The M35 Metalloprotease Effector FocM35_1 Is Required for Full Virulence of Fusarium oxysporum f. sp. cubense Tropical Race 4. Pathogens 2021, 10, 670. [Google Scholar] [CrossRef]
- Caldwell, G.A.; Naider, F.; Becker, J.M. Fungal lipopeptide mating pheromones: A model system for the study of protein prenylation. Microbiol. Rev. 1995, 59, 406–422. [Google Scholar] [CrossRef]
- Casselton, L.A. Mate recognition in fungi. Heredity 2002, 88, 142–147. [Google Scholar] [CrossRef]
- Jenness, D.D.; Burkholder, A.C.; Hartwell, L.H. Binding of α-factor pheromone to yeast a cells: Chemical and genetic evidence for an α-factor receptor. Cell 1983, 35, 521–529. [Google Scholar] [CrossRef]
- Hagen, D.C.; McCaffrey, G.; Sprague, G.F. Evidence the yeast STE3 gene encodes a receptor for the peptide pheromone a factor: Gene sequence and implications for the structure of the presumed receptor. Proc. Natl. Acad. Sci. USA 1986, 83, 1418–1422. [Google Scholar] [CrossRef] [Green Version]
- Bardwell, L. A walk-through of the yeast mating pheromone response pathway. Peptides 2005, 26, 339–350. [Google Scholar] [CrossRef]
- Hoi, J.; Dumas, B. Ste12 and Ste12-Like Proteins, Fungal Transcription Factors Regulating Development and Pathogenicity. Eukaryot. Cell 2010, 9, 480–485. [Google Scholar]
- Singh, A.; Chen, E.Y.; Lugovoy, J.M.; Chang, C.N.; Hitzeman, R.A.; Seeburg, P.H. Saccharomyces cerevisiae contains two discrete genes coding for the α-factor pheromone. Nucleic Acids Res. 1983, 11, 4049–4063. [Google Scholar] [CrossRef]
- Julius, D.; Blair, L.; Brake, A.; Sprague, G.; Thorner, J. Yeast α factor is processed from a larger precursor polypeptide: The essential role of a membrane-bound dipeptidyl aminopeptidase. Cell 1983, 32, 839–852. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.H.; Wingfield, B.D.; Wingfield, M.J.; Steenkamp, E.T. Causes and Consequences of Variability in Peptide Mating Pheromones of Ascomycete Fungi. Mol. Biol. Evol. 2011, 28, 1987–2003. [Google Scholar] [CrossRef] [Green Version]
- Caplan, S.; Kurjan, J. Role of α-Factor and the Mfα1 α-Factor Precursor in Mating in Yeast. Genetics 1991, 127, 299–307. [Google Scholar] [CrossRef]
- Guo, L.; Yang, Y.; Yang, L.; Wang, F.; Wang, G.; Huang, J. Functional analysis of the G-protein α subunits FGA1 and FGA3 in the banana pathogen Fusarium oxysporum f. sp. cubense. Physiol. Mol. Plant Pathol. 2016, 94, 75–82. [Google Scholar] [CrossRef]
- Liu, J.; An, B.; Luo, H.; He, C.; Wang, Q. The histone acetyltransferase FocGCN5 regulates growth, conidiation, and pathogenicity of the banana wilt disease causal agent Fusarium oxysporum f.sp. cubense tropical race 4. Res. Microbiol. 2022, 173, 103902. [Google Scholar] [CrossRef]
- Liu, S.; Li, J.; Zhang, Y.; Liu, N.; Viljoen, A.; Mostert, D.; Zuo, C.; Hu, C.; Bi, F.; Gao, H.; et al. Fusaric acid instigates the invasion of banana by Fusarium oxysporum f. sp. cubense TR4. New Phytol. 2020, 225, 913–929. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Lin, B.; Huang, Q.; Sun, T.; Wang, W.; Liao, J.; Zhuo, K. The Meloidogyne javanica effector Mj2G02 interferes with jasmonic acid signalling to suppress cell death and promote parasitism in Arabidopsis. Mol. Plant Pathol. 2021, 22, 1288–1301. [Google Scholar] [CrossRef]
- Turrà, D.; El Ghalid, M.; Rossi, F.; Di Pietro, A. Fungal pathogen uses sex pheromone receptor for chemotropic sensing of host plant signals. Nature 2015, 527, 521–524. [Google Scholar] [CrossRef] [PubMed]
- Molina, L.; Kahmann, R. An Ustilago maydis Gene Involved in H2O2 Detoxification Is Required for Virulence. Plant Cell 2007, 19, 2293–2309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, X.; Li, X.; Guo, H.; Guo, N.; Cheng, H. VdPLP, A Patatin-Like Phospholipase in Verticillium dahliae, Is Involved in Cell Wall Integrity and Required for Pathogenicity. Genes 2018, 9, 162. [Google Scholar] [CrossRef] [Green Version]
- Sarmiento-Villamil, J.L.; Prieto, P.; Klosterman, S.J.; García-Pedrajas, M.D. Characterization of two homeodomain transcription factors with critical but distinct roles in virulence in the vascular pathogen Verticillium dahliae. Mol. Plant Pathol. 2018, 19, 986–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edlich, F. BCL-2 proteins and apoptosis: Recent insights and unknowns. Biochem. Biophys. Res. Commun. 2018, 500, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Lacomme, C.; Santa Cruz, S. Bax-induced cell death in tobacco is similar to the hypersensitive response. Proc. Natl. Acad. Sci. USA 1999, 96, 7956–7961. [Google Scholar] [CrossRef] [Green Version]
- Bender, A.; Sprague, G.F., Jr. Pheromones and pheromone receptors are the primary determinants of mating specificity in the yeast Saccharomyces cerevisiae. Genetics 1989, 121, 463–476. [Google Scholar] [CrossRef]
- Bobrowicz, P.; Pawlak, R.; Correa, A.; Bell-Pedersen, D.; Ebbole, D.J. The Neurospora crassa pheromone precursor genes are regulated by the mating type locus and the circadian clock. Mol. Microbiol. 2002, 45, 795–804. [Google Scholar] [CrossRef]
- Daniels, K.J.; Srikantha, T.; Lockhart, S.R.; Pujol, C.; Soll, D.R. Opaque cells signal white cells to form biofilms in Candida albicans. EMBO J. 2006, 25, 2240–2252. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-K.; Lee, T.; Yun, S.-H. A putative pheromone signaling pathway is dispensable for self-fertility in the homothallic ascomycete Gibberella zeae. Fungal Genet. Biol. 2008, 45, 1188–1196. [Google Scholar] [CrossRef]
- Sridhar, P.S.; Trofimova, D.; Subramaniam, R.; González-Peña Fundora, D.; Foroud, N.A.; Allingham, J.S.; Loewen, M.C. Ste2 receptor-mediated chemotropism of Fusarium graminearum contributes to its pathogenicity against wheat. Sci. Rep. 2020, 10, 10770. [Google Scholar] [CrossRef] [PubMed]
- Vitale, S.; Partida-Hanon, A.; Serrano, S.; Martínez-del-Pozo, Á.; Di Pietro, A.; Turrà, D.; Bruix, M. Structure-Activity Relationship of α Mating Pheromone from the Fungal Pathogen Fusarium oxysporum. J. Biol. Chem. 2017, 292, 3591–3602. [Google Scholar] [CrossRef] [Green Version]
- Vitale, S.; Di Pietro, A.; Turrà, D. Autocrine pheromone signalling regulates community behaviour in the fungal pathogen Fusarium oxysporum. Nat. Microbiol. 2019, 4, 1443–1449. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.E.; Rose, M.D. Cell fusion in yeast is negatively regulated by components of the cell wall integrity pathway. Mol. Biol. Cell 2019, 30, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Dudin, O.; Merlini, L.; Martin, S.G. Spatial focalization of pheromone/MAPK signaling triggers commitment to cell-cell fusion. Genes Dev. 2016, 30, 2226–2239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, S.G. Molecular mechanisms of chemotropism and cell fusion in unicellular fungi. J. Cell Sci. 2019, 132, jcs230706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sieber, B.; Coronas-Serna, J.M.; Martin, S.G. A focus on yeast mating: From pheromone signaling to cell-cell fusion. Semin. Cell Dev. Biol. 2023, 133, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Frawley, D.; Bayram, Ö. The pheromone response module, a mitogen-activated protein kinase pathway implicated in the regulation of fungal development, secondary metabolism and pathogenicity. Fungal Genet. Biol. 2020, 144, 103469. [Google Scholar] [CrossRef]
- Partida-Hanon, A.; Maestro-López, M.; Vitale, S.; Turrà, D.; Di Pietro, A.; Martínez-del-Pozo, Á.; Bruix, M. Structure of Fungal α Mating Pheromone in Membrane Mimetics Suggests a Possible Role for Regulation at the Water-Membrane Interface. Front. Microbiol. 2020, 11, 1090. [Google Scholar] [CrossRef]
- Nordzieke, D.E.; Fernandes, T.R.; El Ghalid, M.; Turrà, D.; Di Pietro, A. NADPH oxidase regulates chemotropic growth of the fungal pathogen Fusarium oxysporum towards the host plant. New Phytol. 2019, 224, 1600–1612. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Huang, Y.; Song, H.; Luo, M.; Dong, Z. α-Pheromone Precursor Protein Foc4-PP1 Is Essential for the Full Virulence of Fusarium oxysporum f. sp. cubense Tropical Race 4. J. Fungi 2023, 9, 365. https://doi.org/10.3390/jof9030365
Liu L, Huang Y, Song H, Luo M, Dong Z. α-Pheromone Precursor Protein Foc4-PP1 Is Essential for the Full Virulence of Fusarium oxysporum f. sp. cubense Tropical Race 4. Journal of Fungi. 2023; 9(3):365. https://doi.org/10.3390/jof9030365
Chicago/Turabian StyleLiu, Lu, Yinghua Huang, Handa Song, Mei Luo, and Zhangyong Dong. 2023. "α-Pheromone Precursor Protein Foc4-PP1 Is Essential for the Full Virulence of Fusarium oxysporum f. sp. cubense Tropical Race 4" Journal of Fungi 9, no. 3: 365. https://doi.org/10.3390/jof9030365