
Biodegradation of Aflatoxin B1 in Maize Grains and Suppression of Its Biosynthesis-Related Genes Using Endophytic Trichoderma harzianum AYM3



Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Maize Plants and Grains
2.2. Isolation and Phenotypic Identification of the Endophytic Fungi and the Ear Rot Pathogen and Extraction of AFB1
2.3. HPLC Conditions
2.4. Screening for the AFs-Degrading Capability of the Endophytic Fungi In Vitro
2.5. Molecular Identification and Phylogenetic Analysis
2.6. Assessment of the Antifungal Potential of T. harzianum AYM3 against A. flavus AYM2 In Vitro
2.7. Biodegradation of AFB1 by T. harzianum AYM3
2.8. Effect of Co-Culturing of T. harzianum AYM3 and A. flavus AYM2 on AFB1 Production
2.9. Expression Profiling of AF Biosynthesis-Related Genes
2.10. Detection of the Bioactive Anti-Aflatoxigenic Compounds from the Culture Filtrate of T. harzianum AYM3
2.11. Gas Chromatography/Mass Spectrometry (GC/MS) Analysis
2.12. Cytotoxicity of the Secondary Metabolites of T. harzianum AYM3
2.13. Statistical Analyses
3. Results
3.1. Isolation and Identification of the Endophytic Fungi and the Ear Rot Pathogen
3.2. Screening for the AFs-Degrading Capability of the Endophytic Fungi In Vitro
3.3. Molecular Identification and Phylogenetic Analysis
3.4. Antifungal Activity of T. harzianum AYM3 against A. flavus AYM2
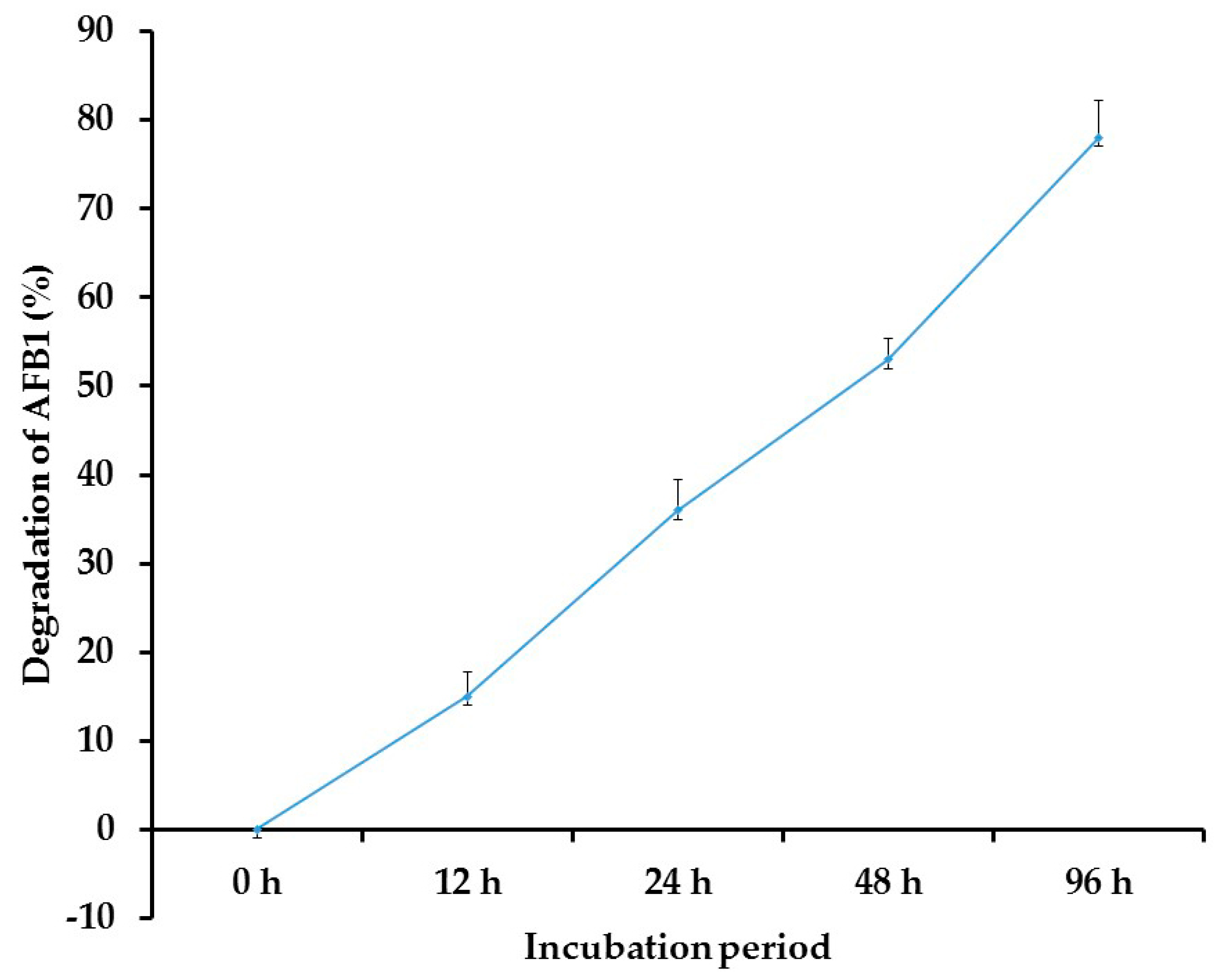
3.5. Biodegradation of AFB1 by T. harzianum AYM3
3.6. Effect of Co-Culturing of T. harzianum AYM3 and A. flavus AYM2 on AFB1 Production
3.7. Expression Profiling of AF Biosynthesis-Related Genes
3.8. TLC Analysis of T. harzianum AYM3 Metabolites
3.9. GC-MS Analysis
3.10. Cytotoxicity of Secondary Metabolites of T. harzianum AYM3
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT. Statistics Division; FAO: Rome, Italy, 2022; Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 12 May 2022).
- Wu, F.; Guclu, H. Global Maize Trade and Food Security: Implications from a Social Network Model. Risk Anal. 2013, 33, 2168–2178. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Y.; Du, Z.; Lin, D.; Huo, L.; Qin, L.; Wang, W.; Qiang, L.; Yao, Y.; An, Y. Screening, identification and antagonistic effect of antagonistic bacteria JTFM1001 against aflatoxin contamination in corn. Oil Crop Sci. 2021, 6, 1–7. [Google Scholar] [CrossRef]
- Ren, X.; Branà, M.T.; Haidukowski, M.; Gallo, A.; Zhang, Q.; Logrieco, A.F.; Li, P.; Zhao, S.; Altomare, C. Potential of Trichoderma spp. for Biocontrol of Aflatoxin-Producing Aspergillus flavus. Toxins 2022, 14, 86. [Google Scholar] [CrossRef]
- Dini, I.; Alborino, V.; Lanzuise, S.; Lombardi, N.; Marra, R.; Balestrieri, A.; Ritieni, A.; Woo, S.L.; Vinale, F. Trichoderma Enzymes for Degradation of Aflatoxin B1 and Ochratoxin A. Molecules 2022, 27, 3959. [Google Scholar] [CrossRef]
- Wu, F. Perspective: Time to face the fungal threat. Nature 2014, 516, S7. [Google Scholar] [CrossRef]
- Cleveland, T.E.; Yu, J.; Fedorova, N.; Bhatnagar, D.; Payne, G.A.; Nierman, W.C.; Bennett, J.W. Potential of Aspergillus flavus genomics for applications in biotechnology. Trends Biotechnol. 2009, 27, 151–157. [Google Scholar] [CrossRef]
- Amaike, S.; Keller, N.P. Aspergillus flavus. Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef]
- Xing, F.; Wang, L.; Liu, X.; Selvaraj, J.N.; Wang, Y.; Zhao, Y.; Liu, Y. Aflatoxin B1 inhibition in Aspergillus flavus by Aspergillus niger through downregulating expression of major biosynthetic genes and AFB1 degradation by atoxigenic A. flavus. Int. J. Food Microbiol. 2017, 256, 1–10. [Google Scholar] [CrossRef]
- Wang, L.; Huang, W.; Sha, Y.; Yin, H.; Liang, Y.; Wang, X.; Shen, Y.; Wu, X.; Wu, D.; Wang, J. Co-Cultivation of Two Bacillus Strains for Improved Cell Growth and Enzyme Production to Enhance the Degradation of Aflatoxin B1. Toxins 2021, 13, 435. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, F.; Zhou, X.; Liu, M.; Zang, H.; Liu, X.; Shan, A.; Feng, X. Alleviation of Oral Exposure to Aflatoxin B1-Induced Renal Dysfunction, Oxidative Stress, and Cell Apoptosis in Mice Kidney by Curcumin. Antioxidants 2022, 11, 1082. [Google Scholar] [CrossRef]
- Ostry, V.; Malir, F.; Toman, J.; Grosse, Y. Mycotoxins as human carcinogens-the IARC Monographs classification. Mycotoxin Res. 2017, 33, 65–73. [Google Scholar] [CrossRef]
- Shcherbakova, L.; Statsyuk, N.; Mikityuk, O.; Nazarova, T.; Dzhavakhiya, V. Aflatoxin B1 degradation by metabolites of Phoma glomerata PG41 isolated from natural substrate colonized by aflatoxigenic Aspergillus flavus. Jundishapur J. Microbiol. 2015, 8, e24324. [Google Scholar] [CrossRef] [PubMed]
- Kabak, B. The fate of mycotoxins during thermal food processing. J. Sci. Food Agric. 2009, 89, 549–554. [Google Scholar] [CrossRef]
- Vipin, A.V.; Rao, R.; Kurrey, N.K.; Appaiah, A.K.A.; Venkateswaran, G. Protective effects of phenolics rich extract of ginger against Aflatoxin B1-induced oxidative stress and hepatotoxicity. Biomed. Pharmacother. 2017, 91, 415–424. [Google Scholar] [CrossRef]
- Rao, K.R.; Vipina, A.V.; Venkateswarana, G. Mechanism of inhibition of aflatoxin synthesis by non-aflatoxigenic strains of Aspergillus flavus. Microb. Pathog. 2020, 147, 104280. [Google Scholar] [CrossRef]
- Ding, N.; Xing, F.; Liu, X.; Selvaraj, J.N.; Wang, L.; Zhao, Y.; Wang, Y.; Guo, W.; Dai, X.; Liu, Y. Variation in fungal microbiome (mycobiome) and aflatoxin in stored in-shell peanuts at four different areas of China. Front. Microbiol. 2015, 6, 1055. [Google Scholar] [CrossRef]
- Guo, B.; Chen, X.; Dang, P.; Scully, B.T.; Liang, X.; Holbrook, C.C.; Yu, J.; Culbreath, A.K. Peanut gene expression profiling in developing seeds at different reproduction stages during Aspergillus parasiticus infection. BMC Dev. Biol. 2008, 8, 12. [Google Scholar] [CrossRef]
- Soni, P.; Gangurde, S.S.; Ortega-Beltran, A.; Kumar, R.; Parmar, S.; Sudini, H.K.; Lei, Y.; Ni, X.; Huai, D.; Fountain, J.C.; et al. Functional biology and molecular mechanisms of host-pathogen interactions for aflatoxin contamination in groundnut (Arachis hypogaea L.) and maize (Zea mays L.). Front. Microbiol. 2020, 11, 227. [Google Scholar] [CrossRef]
- Agbetiameh, D.; Ortega-Beltran, A.; Awuah, R.T.; Atehnkeng, J.; Elzein, A.; Cotty, P.J.; Bandyopadhyay, R. Field efficacy of two atoxigenic biocontrol products for mitigation of aflatoxin contamination in maize and groundnut in Ghana. Biol. Control. 2020, 150, 104351. [Google Scholar] [CrossRef]
- Okwara, P.C.; Afolabi, I.S.; Ahuekwe, E.F. Application of laccase in aflatoxin B1 degradation: A review. IOP Conf. Ser. Mater. Sci. Eng. 2021, 1107, 012178. [Google Scholar] [CrossRef]
- Bilal, S.; Ali, L.; Khan, A.L.; Shahzad, R.; Asaf, S.; Imran, M.; Kang, S.-M.; Kim, S.-K.; Lee, I.-J. Endophytic fungus Paecilomyces formosus LHL10 produces sesterterpenoid YW3548 and cyclic peptide that inhibit urease and α-glucosidase enzyme activities. Arch. Microbiol. 2018, 200, 1493–1502. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.N.; Kumar, R.; Kumar, S.; Kumar, V.; Sugitha, T.; Singh, B.; Chauahan, V.S.; Dhaliwal, H.S.; Saxena, A.K. Beneficial microbiomes: Biodiversity and potential biotechnological applications for sustainable agriculture and human health. J. Appl. Biol. Biotechnol. 2017, 5, 1–13. [Google Scholar] [CrossRef]
- Guan, S.; Ji, C.; Zhou, T.; Li, J.; Ma, Q.; Niu, T. Aflatoxin B1 Degradation by Stenotrophomonas maltophilia and Other Microbes Selected Using Coumarin Medium. Int. J. Mol. Sci. 2008, 9, 1489–1503. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Xu, Y.; Yang, Q. Antifungal properties and AFB1 detoxification activity of a new strain of Lactobacillus plantarum. J. Hazard. Mater. 2021, 414, 125569. [Google Scholar] [CrossRef]
- Vinale, F.; Flematti, G.; Sivasithamparam, K.; Lorito, M.; Marra, R.; Skelton, B.W.; Ghisalberti, E.L. Harzianic Acid, an Antifungal and Plant Growth Promoting Metabolite from Trichoderma harzianum. J. Nat. Prod. 2009, 72, 2032–2035. [Google Scholar] [CrossRef]
- Yue, X.; Ren, X.; Fu, J.; Wei, N.; Altomare, C.; Haidukowski, M.; Logrieco, A.F.; Zhang, Q.; Li, P. Characterization and mechanism of aflatoxin degradation by a novel strain of Trichoderma reesei CGMCC3.5218. Front. Microbiol. 2022, 13, 1003039. [Google Scholar] [CrossRef]
- El-Sharkawy, H.H.; Rashad, Y.M.; Ibrahim, S.A. Biocontrol of stem rust disease of wheat using arbuscular mycorrhizal fungi and Trichoderma spp. Physiol. Mol. Plant Pathol. 2018, 103, 84–91. [Google Scholar] [CrossRef]
- Braun, H.; Woitsch, L.; Hetzer, B.; Geisen, R.; Zange, B.; Schmidt-Heydt, M. Trichoderma harzianum: Inhibition of mycotoxin producing fungi and toxin biosynthesis. Int. J. Food Microbiol. 2018, 280, 10–16. [Google Scholar] [CrossRef]
- Rashad, Y.M.; Moussa, T.A.A. Biocontrol Agents for Fungal Plant Diseases Management. In Cottage Industry of Biocontrol Agents and Their Applications; El-Wakeil, N., Saleh, M., Abu-hashim, M., Eds.; Springer: Cham, Switzerland, 2019; pp. 337–363. [Google Scholar] [CrossRef]
- Sivparsad, B.J.; Laing, M.D. Pre-harvest silk treatment with Trichoderma harzianum reduces aflatoxin contamination in sweetcorn. J. Plant Dis. Prot. 2016, 123, 285–293. [Google Scholar] [CrossRef]
- Caceres, I.; Khoury, A.A.I.; El Khoury, R.; Lorber, S.; Oswald, I.P.; El Khoury, A.; Atoui, A.; Puel, O.; Bailly, J.-D. Aflatoxin Biosynthesis and Genetic Regulation: A Review. Toxins 2020, 12, 150. [Google Scholar] [CrossRef]
- Chatterjee, S.; Ghosh, R.; Mandal, N.C. Inhibition of biofilm- and hyphal- development, two virulent features of Candida albicans by secondary metabolites of an endophytic fungus Alternaria tenuissima having broad spectrum antifungal potential. Microbiol. Res. 2020, 232, 126386. [Google Scholar] [CrossRef] [PubMed]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Ames, IA, USA, 2006. [Google Scholar] [CrossRef]
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.-B.; Hubka, V.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Susca, A.; Tanney, J.B.; et al. Phylogeny, identification and nomenclature of the genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef] [PubMed]
- Horn, B.; Singh, K.; Frisvad, J.C.; Thrane, U.; Mathur, S.B. An Illustrated Manual on Identification of Some Seed-Borne Aspergilli, Fusaria, and Penicillia and Their Mycotoxins. Mycologia 1993, 85, 139. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC, 15th ed.; Association of Official Analytical Chemists, Inc.: Rockville, MD, USA, 1990; pp. 1184–1213. [Google Scholar]
- Ali, S.; Hassan, M.; Essam, T.; Ibrahim, M.A.; Al-Amry, K. Biodegradation of aflatoxin by bacterial species isolated from poultry farms. Toxicon 2021, 195, 7–16. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) Software Version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Rahman, M.A.; Begum, M.F.; Alam, M.F. Screening of Trichoderma Isolates as a Biological Control Agent against Ceratocystis paradoxa Causing Pineapple Disease of Sugarcane. Mycobiology 2009, 37, 277–285. [Google Scholar] [CrossRef]
- Abdel-Shafi, S.; Shehata, S.; Shindia, A.; El-Meligy, K.; Khidr, A. Biodegradation of aflatoxins by bacteria. Egypt. J. Microbiol. 2018. [Google Scholar] [CrossRef]
- Wong, D.; Barbetti, M.; Sivasithamparam, K. Effects of soil temperature and moisture on the pathogenicity of fungi associated with root rot of subterranean clover. Aust. J. Agric. Res. 1984, 35, 675–684. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Suebrasri, T.; Somteds, A.; Harada, H.; Kanokmedhakul, S.; Jogloy, S.; Ekprasert, J.; Lumyong, S.; Boonlue, S. Novel endophytic fungi with fungicidal metabolites suppress sclerotium disease. Rhizosphere 2020, 16, 100250. [Google Scholar] [CrossRef]
- Roopan, S.M.; Kumar, S.H.S.; Madhumitha, G.; Suthindhiran, K. Biogenic-Production of SnO2 Nanoparticles and Its Cytotoxic Effect Against Hepatocellular carcinoma Cell Line (HepG2). Appl. Biochem. Biotechnol. 2015, 175, 1567–1575. [Google Scholar] [CrossRef] [PubMed]
- Qi-Zhang, L.; Zhu-Mei, H. Advances in research of the structural gene characteristics of the aflatoxin biosynthetic gene cluster. J. Plant Sci. Phytopathol. 2018, 2, 068–082. [Google Scholar] [CrossRef]
- Potshangbam, M.; Devi, S.I.; Sahoo, D.; Strobel, G.A. Functional Characterization of Endophytic Fungal Community Associated with Oryza sativa L. and Zea mays L. Front. Microbiol. 2017, 8, 325. [Google Scholar] [CrossRef] [PubMed]
- Gizachew, D.; Chang, C.-H.; Szonyi, B.; De La Torre, S.; Ting, W.-T.E. Aflatoxin B1 (AFB1) production by Aspergillus flavus and Aspergillus parasiticus on ground Nyjer seeds: The effect of water activity and temperature. Int. J. Food Microbiol. 2019, 296, 8–13. [Google Scholar] [CrossRef]
- Hackbart, H.C.S.; Machado, A.R.; Christ-Ribeiro, A.; Prietto, L.; Badiale-Furlong, E. Reduction of aflatoxins by Rhizopus oryzae and Trichoderma reesei. Mycotoxin Res. 2014, 30, 141–149. [Google Scholar] [CrossRef]
- Tomin, M.; Tomić, S. Oxidase or peptidase? A computational insight into a putative aflatoxin oxidase from Armillariella tabescens. Proteins 2019, 87, 390–400. [Google Scholar] [CrossRef]
- Xiong, D.; Wen, J.; Lu, G.; Li, T.; Long, M. Isolation, Purification, and Characterization of a Laccase-Degrading Aflatoxin B1 from Bacillus amyloliquefaciens B10. Toxins 2022, 14, 250. [Google Scholar] [CrossRef]
- Wang, X.; Qin, X.; Hao, Z.; Luo, H.; Yao, B.; Su, X. Degradation of Four Major Mycotoxins by Eight Manganese Peroxidases in Presence of a Dicarboxylic Acid. Toxins 2019, 11, 566. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Shu, X.; Wang, D.; Kan, W.; Su, P.; Hu, H.; Chen, X.; Wang, D.; Huang, S.; Wu, L. Non-enzymatic Transformation of Aflatoxin B1 by Pseudomonas geniculata m29. Front. Microbiol. 2021, 12, 724103. [Google Scholar] [CrossRef]
- Metz, N.; Hausladen, H. Trichoderma spp. As potential biological control agent against Alternaria solani in potato. Biol. Control. 2022, 166, 104820. [Google Scholar] [CrossRef]
- Monteiro, V.N.; do Nascimento Silva, R.; Steindorff, A.S.; Costa, F.T.; Noronha, E.F.; Ricart, C.A.O.; De Sousa, M.V.; Vainstein, M.H.; Ulhoa, C.J. New Insights in Trichoderma harzianum Antagonism of Fungal Plant Pathogens by Secreted Protein Analysis. Curr. Microbiol. 2010, 61, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Baiyee, B.; Pornsuriya, C.; Ito, S.-I.; Sunpapao, A. Trichoderma spirale T76-1 displays biocontrol activity against leaf spot on lettuce (Lactuca sativa L.) caused by Corynespora cassiicola or Curvularia aeria. Biol. Control. 2018, 129, 195–200. [Google Scholar] [CrossRef]
- Ruangwong, O.-U.; Wonglom, P.; Phoka, N.; Suwannarach, N.; Lumyong, S.; Ito, S.-I.; Sunpapao, A. Biological control activity of Trichoderma asperelloides PSU-P1 against gummy stem blight in muskmelon (Cucumis melo). Physiol. Mol. Plant Pathol. 2021, 115, 101663. [Google Scholar] [CrossRef]
- Adebo, O.A.; Njobeh, P.B.; Gbashi, S.; Nwinyi, O.C.; Mavumengwana, V. Review on microbial degradation of aflatoxins. Crit. Rev. Food Sci. Nutr. 2015, 57, 3208–3217. [Google Scholar] [CrossRef] [PubMed]
- El-Shiekh, H.H.; Mahdy, H.M.; El-Aaser, M.M. Bioremediation of aflatoxins by some reference fungal strains. Pol. J. Microbiol. 2007, 56, 215–223. [Google Scholar]
- Iannaccone, F.; Alborino, V.; Dini, I.; Balestrieri, A.; Marra, R.; Davino, R.; Di Francia, A.; Masucci, F.; Serrapica, F.; Vinale, F. In Vitro Application of Exogenous Fibrolytic Enzymes from Trichoderma Spp. to Improve Feed Utilization by Ruminants. Agriculture 2022, 12, 573. [Google Scholar] [CrossRef]
- de Oliveira, F.K.; Santos, L.O.; Buffon, J.G. Mechanism of action, sources, and application of peroxidases. Food Res. Int. 2021, 143, 110266. [Google Scholar] [CrossRef]
- Khan, R.; Ghazali, F.M.; Mahyudin, N.A.; Samsudin, N.I.P. Aflatoxin Biosynthesis, Genetic Regulation, Toxicity, and Control Strategies: A Review. J. Fungi 2021, 7, 606. [Google Scholar] [CrossRef]
- Yu, J.; Woloshuk, C.P.; Bhatnagar, D.; E Cleveland, T. Cloning and characterization of avfA and omtB genes involved in aflatoxin biosynthesis in three Aspergillus species. Gene 2000, 248, 157–167. [Google Scholar] [CrossRef]
- Yabe, K.; Ando, Y.; Hashimoto, J.; Hamasaki, T. Two distinct O-methyltransferases in aflatoxin biosynthesis. Appl. Environ. Microbiol. 1989, 55, 2172–2177. [Google Scholar] [CrossRef]
- Yu, J.-H.; Butchko, R.A.E.; Fernandes, M.; Keller, N.P.; Leonard, T.J.; Adams, T.H. Conservation of structure and function of the aflatoxin regulatory gene aflR from Aspergillus nidulans and A. flavus. Curr. Genet. 1996, 29, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Faustinelli, P.C.; Palencia, E.R.; Sobolev, V.S.; Horn, B.W.; Sheppard, H.T.; Lamb, M.C.; Wang, X.M.; Scheffler, B.E.; Castillo, J.M.; Arias, R.S. Study of the genetic diversity of the aflatoxin biosynthesis cluster in Aspergillus section Flavi using insertion/deletion markers in peanut seeds from Georgia, USA. Mycologia 2017, 109, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.; Chi, C.; Yu, J.; Shan, S.; Li, Q.; Li, Q.; Guan, B.; Nierman, W.C.; Bennett, J.W. The inhibitory effect of Bacillus megaterium on aflatoxin and cyclopiazonic acid biosynthetic pathway gene expression in Aspergillus flavus. Appl. Environ. Microbiol. 2014, 98, 5161–5172. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.-K. The Aspergillus parasiticus protein AFLJ interacts with the aflatoxin pathway-specific regulator AFLR. Mol. Genet. Genom. 2003, 268, 711–719. [Google Scholar] [CrossRef]
- Moon, Y.-S.; Kim, H.-M.; Chun, H.S.; Lee, S.-E. Organic acids suppress aflatoxin production via lowering expression of aflatoxin biosynthesis-related genes in Aspergillus flavus. Food Control. 2018, 88, 207–216. [Google Scholar] [CrossRef]
- Aiko, V.; Edamana, P.; Mehta, A. Decomposition and detoxification of aflatoxin B1 by lactic acid. J. Sci. Food Agric. 2015, 96, 1959–1966. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Code | Accession No. | Sequence (5′–3′) |
|---|---|---|
| aflD-F | AFLA_139390 | AGGCATCTGTGCTCGGATTG |
| aflD-R | TGCCCCGATGTAGTCTCCTTAGT | |
| aflO-F | AFLA_139220 | CCCCAAGAGTATACCTCGAGTGC |
| aflO-R | AAGGTCCCGAGATGTCGAATAGTT | |
| aflP-F | AFLA_139210 | TATTCTACATGACTATCCCGATGCTG |
| aflP-R | GCGCGACTTGCTTGGGT | |
| aflR-F | AFLA_139360 | GCGGCACAGCTTGTTCTGA |
| aflR-R | CCGGTATCCCTGCTGCATC | |
| aflS-F | AFLA_139340 | AACGGTCGTGCATGTGGG |
| aflS-R | CGGCCTTAGCTTCTGTCTGC | |
| β-tub-F | AFLA_068620 | AACGTCTACTTCAACGAGGCCA |
| β-tub-R | GTACCAGGCTCAAGATCAACGAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madbouly, A.K.; Rashad, Y.M.; Ibrahim, M.I.M.; Elazab, N.T. Biodegradation of Aflatoxin B1 in Maize Grains and Suppression of Its Biosynthesis-Related Genes Using Endophytic Trichoderma harzianum AYM3. J. Fungi 2023, 9, 209. https://doi.org/10.3390/jof9020209
Madbouly AK, Rashad YM, Ibrahim MIM, Elazab NT. Biodegradation of Aflatoxin B1 in Maize Grains and Suppression of Its Biosynthesis-Related Genes Using Endophytic Trichoderma harzianum AYM3. Journal of Fungi. 2023; 9(2):209. https://doi.org/10.3390/jof9020209
Chicago/Turabian StyleMadbouly, Adel K., Younes M. Rashad, Mohamed I. M. Ibrahim, and Nahla T. Elazab. 2023. "Biodegradation of Aflatoxin B1 in Maize Grains and Suppression of Its Biosynthesis-Related Genes Using Endophytic Trichoderma harzianum AYM3" Journal of Fungi 9, no. 2: 209. https://doi.org/10.3390/jof9020209