
Entomopathogenic Fungi-Mediated Solubilization and Induction of Fe Related Genes in Melon and Cucumber Plants
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolates and Inoculum Preparation
2.2. In Vitro Study of Fe Biodisponibility by Production of Siderophores
2.3. In Planta and Soil Studies of Fe Biodisponibility
2.4. Ferric Reductase Activity and Fe Acquisition Gene Expression
2.4.1. Growth Conditions and Vegetal Material
2.4.2. Inoculum Preparation and Roots Priming
2.4.3. Measure of Ferric Reductase Activity (FRA)
2.4.4. RNA Isolation, cDNA Synthesis and qRT-PCR Analysis
2.5. Data Analysis
3. Results
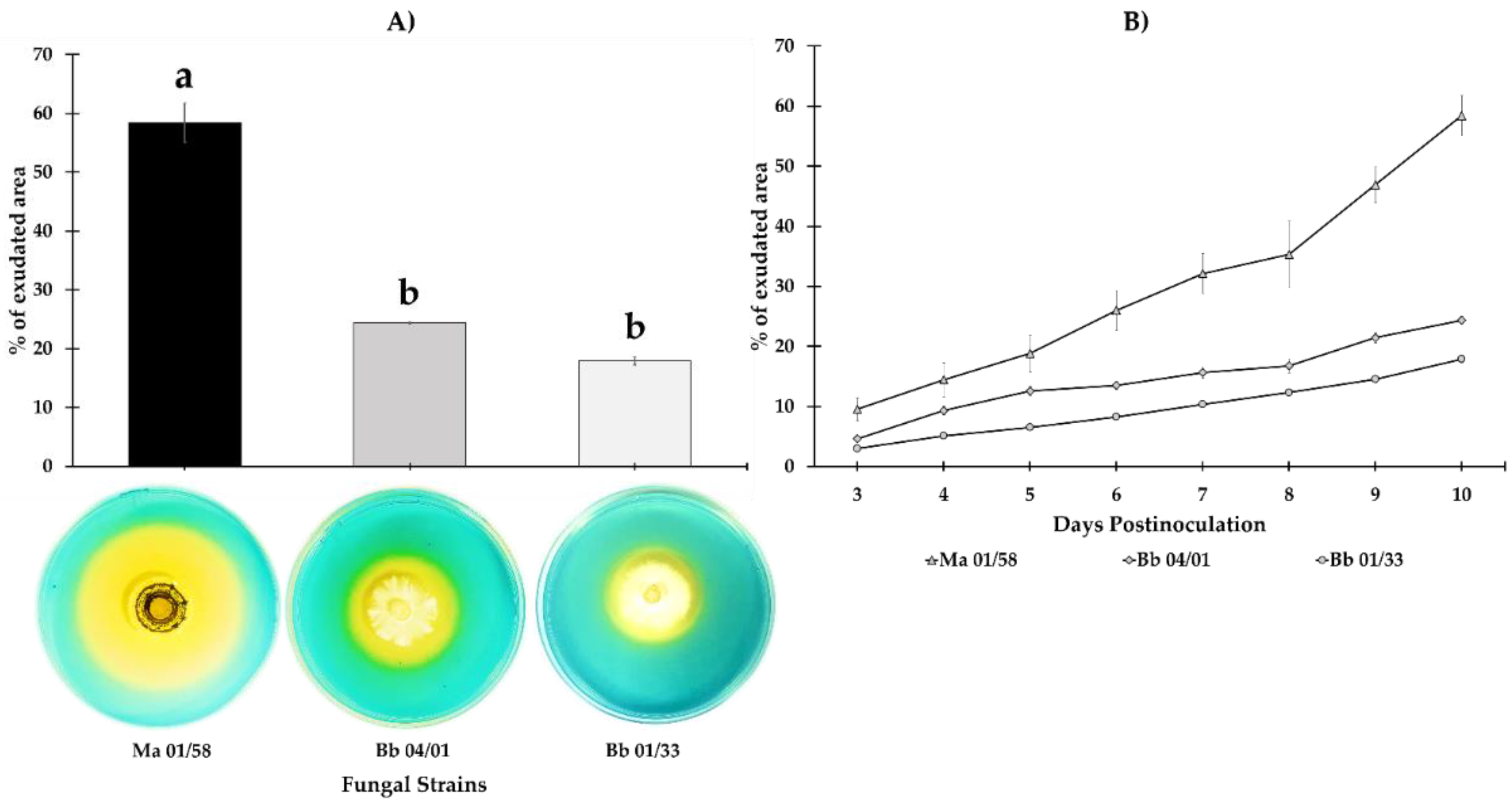
3.1. Iron Siderophores Exudation
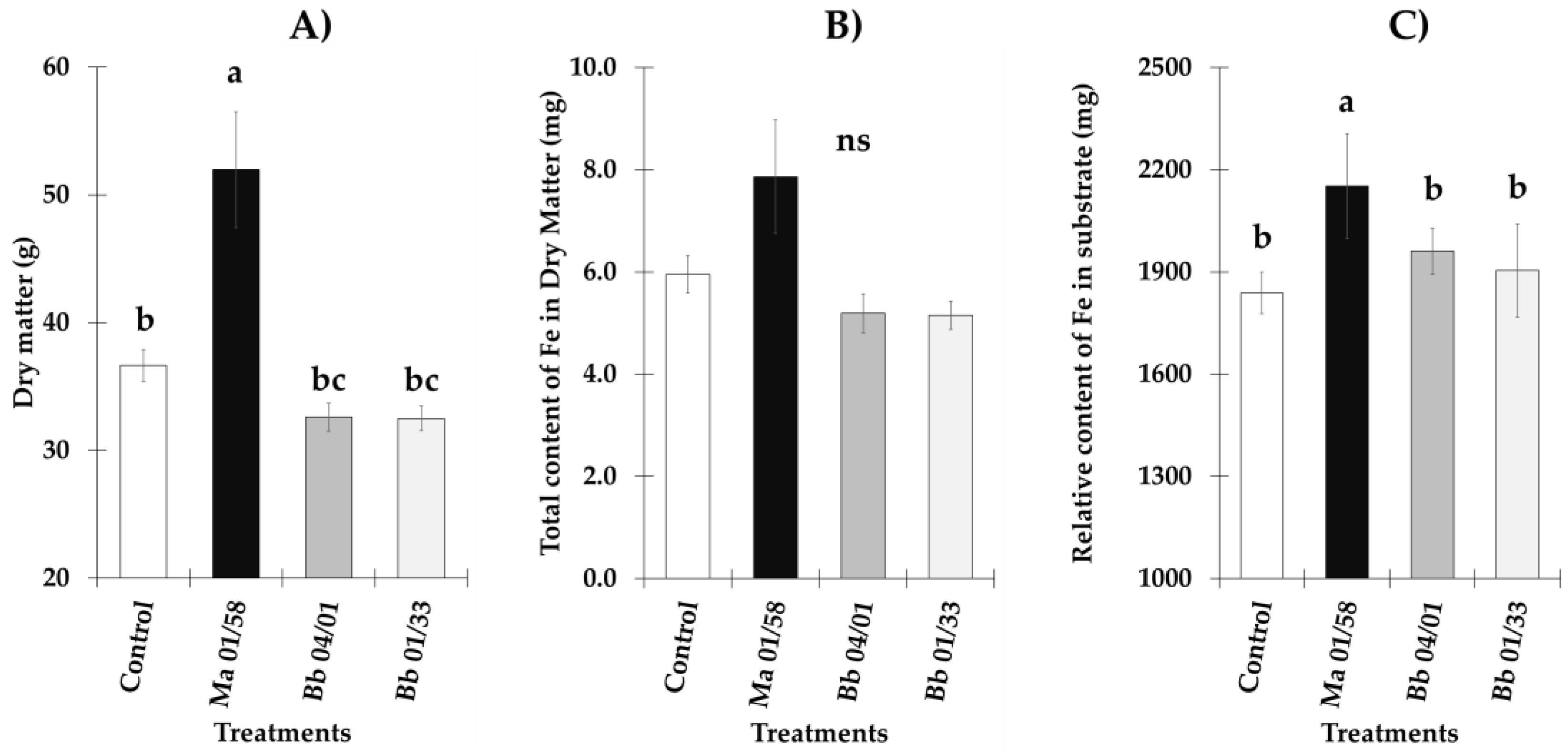
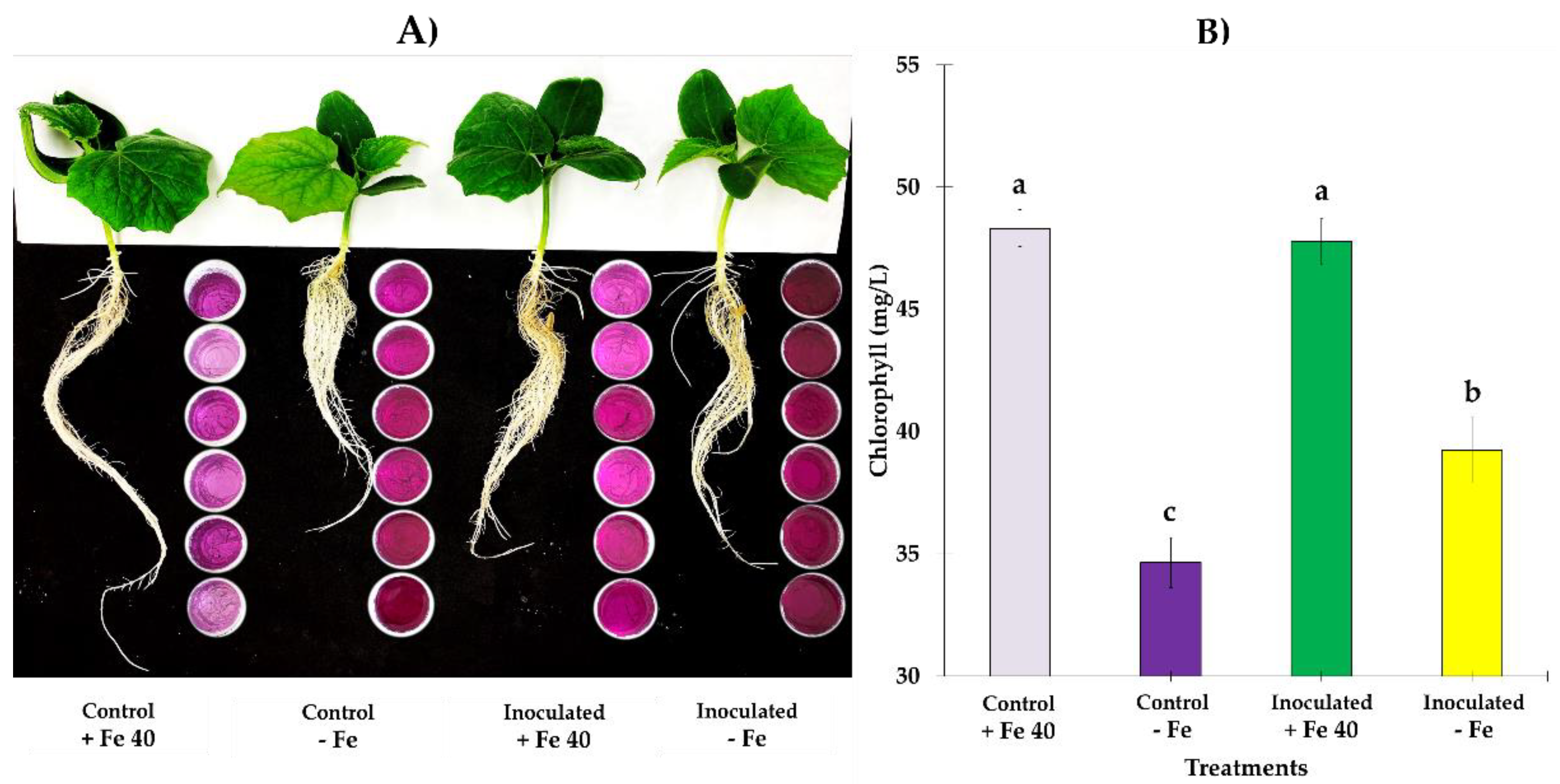
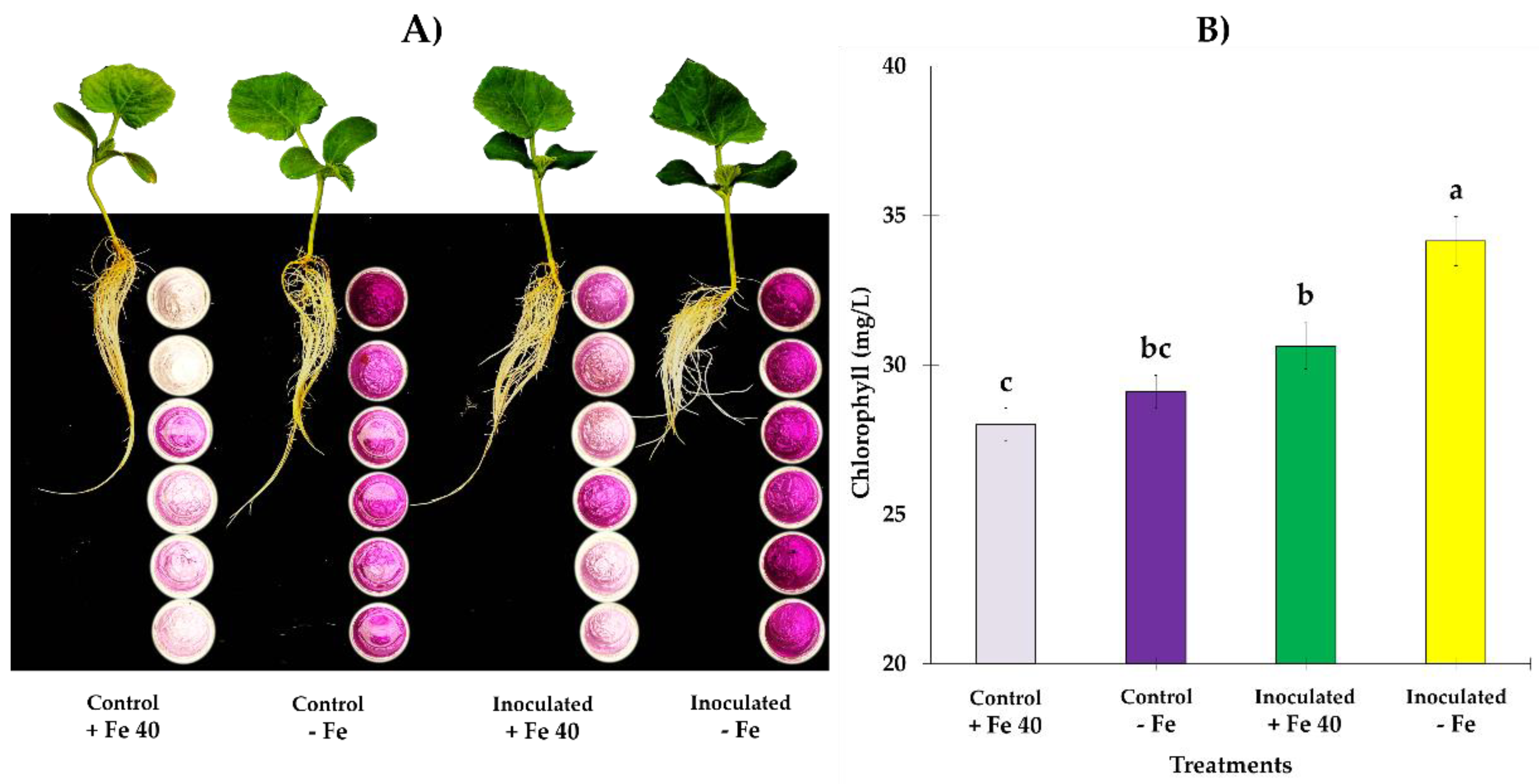
3.2. Total Dry Matter and Fe Content in Dry Matter and Substrate
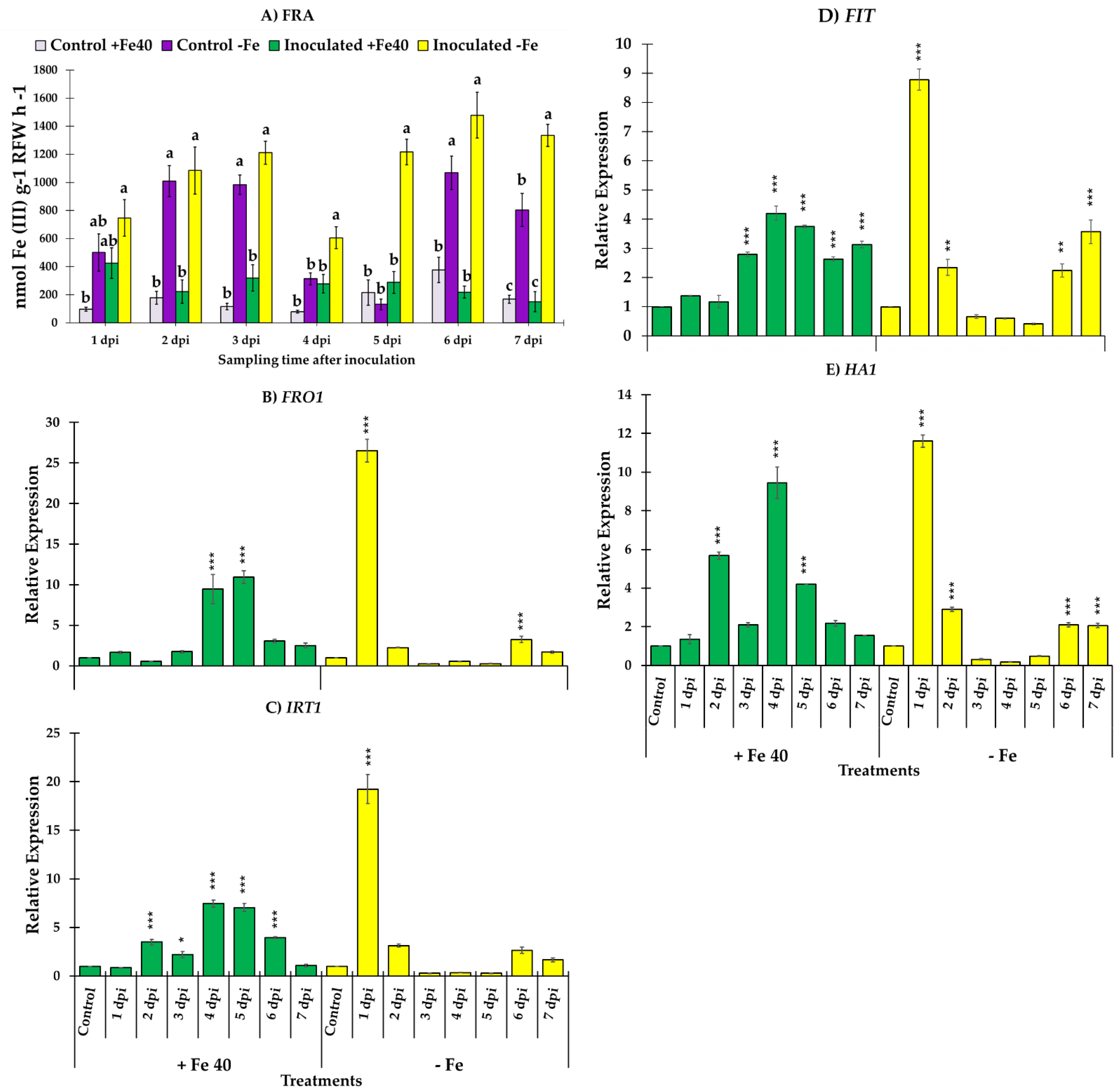
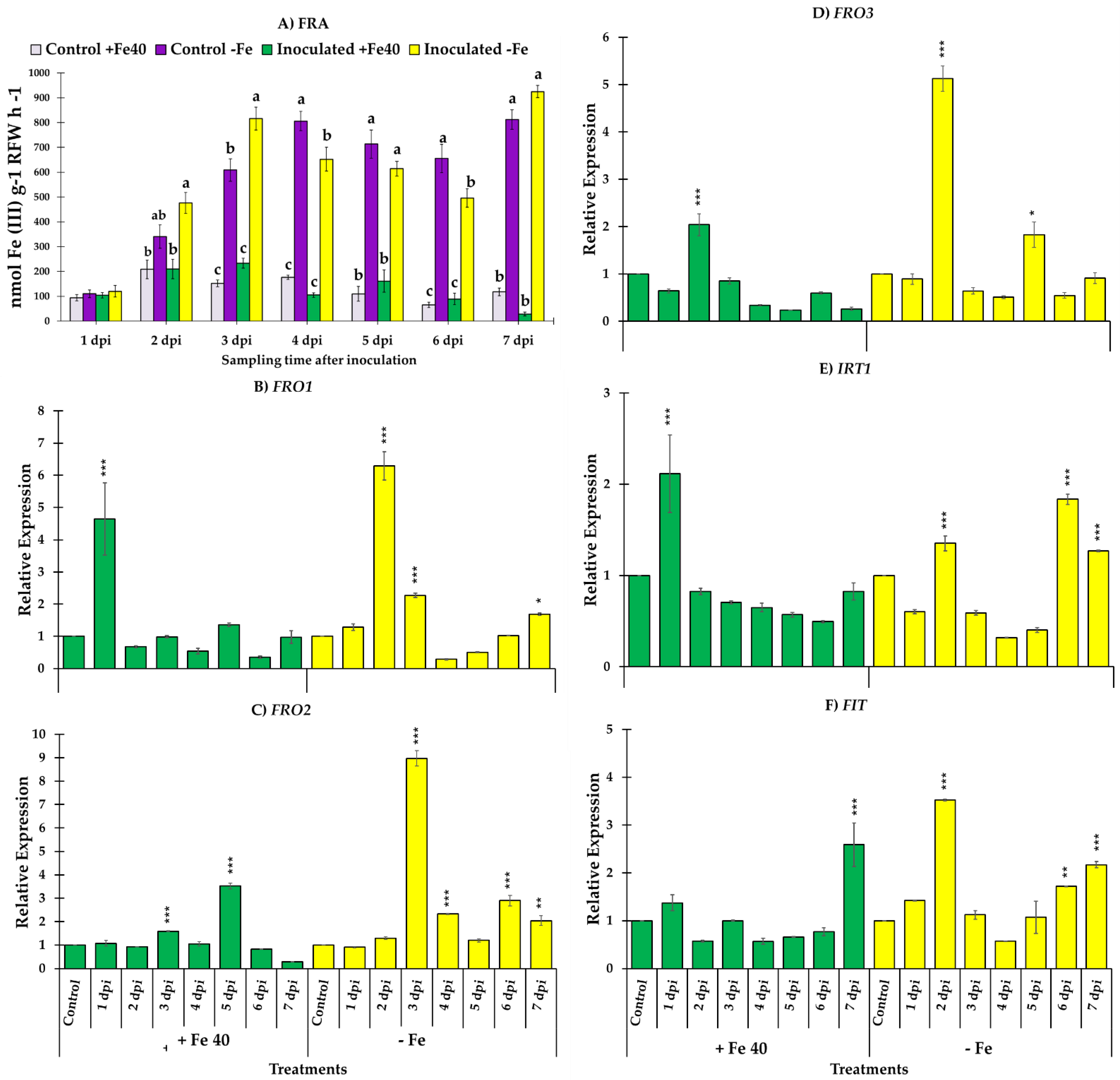
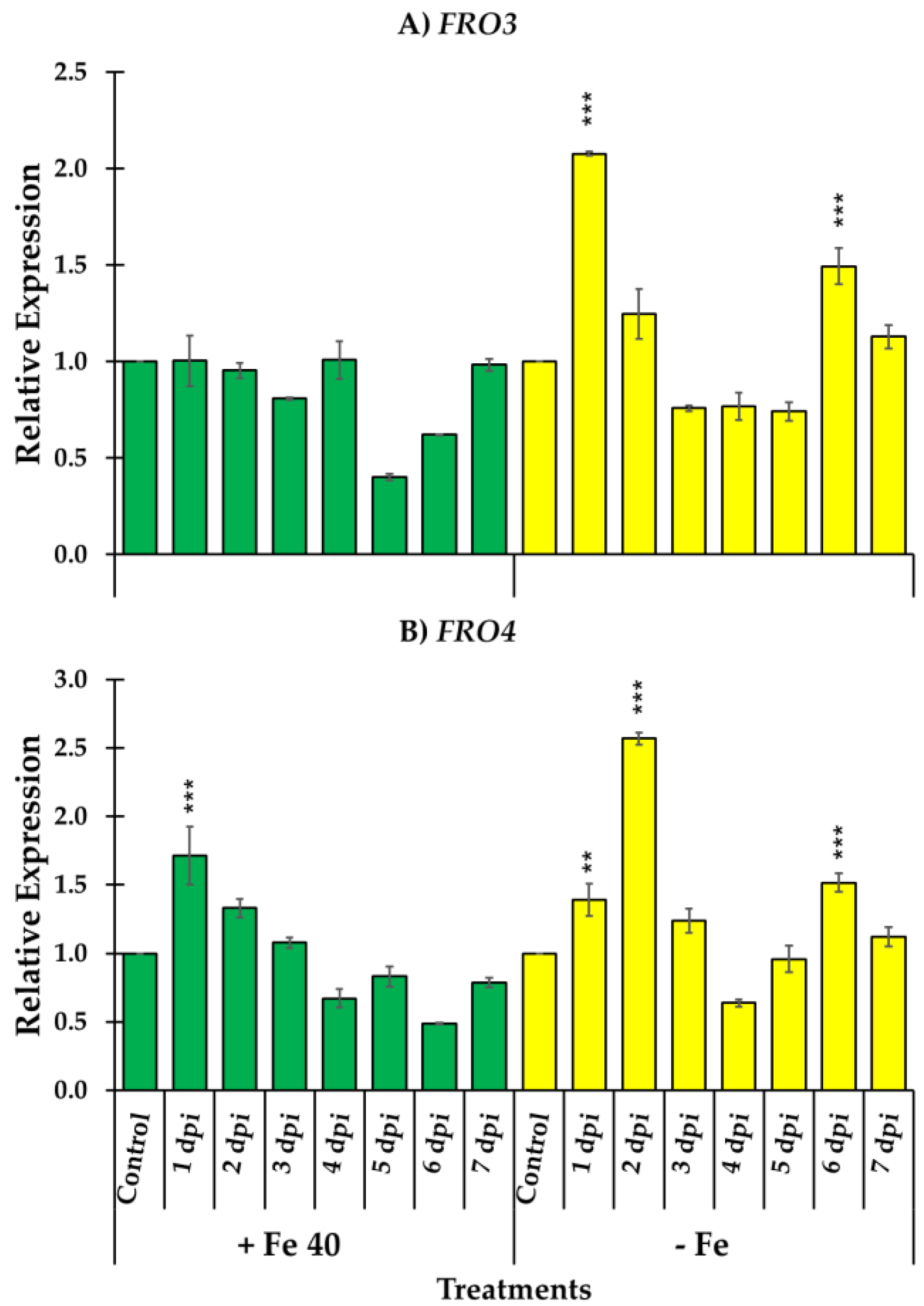
3.3. Ferric Reductase Activity and Genes Responsible of the Reduction and Transport of Iron

4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hu, S.; Bidochka, M.J. Root Colonization by Endophytic Insect-Pathogenic Fungi. J. Appl. Microbiol. 2019, 130, 570–581. [Google Scholar] [CrossRef]
- Quesada-Moraga, E. Entomopathogenic Fungi as Endophytes: Their Broader Contribution to IPM and Crop Production. Biocontrol Sci. Technol. 2020, 30, 864–877. [Google Scholar] [CrossRef]
- Nosheen, S.; Ajmal, I.; Song, Y. Microbes as Biofertilizers, a Potential Approach for Sustainable Crop Production. Sustainability 2021, 13, 1868. [Google Scholar] [CrossRef]
- Branine, M.; Bazzicalupo, A.; Branco, S. Biology and Applications of Endophytic Insect-Pathogenic Fungi. PLoS Pathog. 2019, 15, e1007831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega, F.E. The Use of Fungal Entomopathogens as Endophytes in Biological Control: A Review. Mycologia 2018, 110, 4–30. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L.; Bohra, N.; Rajput, V.D.; Quiroz-Figueroa, F.R.; Singh, R.K.; Marques, G. Advances in Entomopathogen Isolation: A Case of Bacteria and Fungi. Microorganisms 2021, 9, 16. [Google Scholar] [CrossRef] [PubMed]
- Barelli, L.; Moonjely, S.; Behie, S.W.; Bidochka, M.J. Fungi with Multifunctional Lifestyles: Endophytic Insect Pathogenic Fungi. Plant Mol. Biol. 2016, 90, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rodríguez, A.R.; Raya-Díaz, S.; Zamarreño, Á.M.; García-Mina, J.M.; del Campillo, M.C.; Quesada-Moraga, E. An Endophytic Beauveria bassiana Strain Increases Spike Production in Bread and Durum Wheat Plants and Effectively Controls Cotton Leafworm (Spodoptera littoralis) Larvae. Biol. Control 2018, 116, 90–102. [Google Scholar] [CrossRef]
- Raya-Díaz, S.; Sánchez-Rodríguez, A.R.; Segura-Fernández, J.M.; del Campillo, M.D.C.; Quesada-Moraga, E. Entomopathogenic Fungi-Based Mechanisms for Improved Fe Nutrition in Sorghum Plants Grown on Calcareous Substrates. PLoS ONE 2017, 12, e0185903. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Rodríguez, A.R.; Barrón, V.; del Campillo, M.C.; Quesada-Moraga, E. The Entomopathogenic Fungus Metarhizium brunneum: A Tool for Alleviating Fe Chlorosis. Plant Soil 2016, 406, 295–310. [Google Scholar] [CrossRef]
- Barra-Bucarei, L.; González, M.G.; Iglesias, A.F.; Aguayo, G.S.; Peñalosa, M.G.; Vera, P.V. Beauveria bassiana Multifunction as an Endophyte: Growth Promotion and Biologic Control of Trialeurodes vaporariorum, (Westwood) (Hemiptera: Aleyrodidae) in Tomato. Insects 2020, 11, 591. [Google Scholar] [CrossRef] [PubMed]
- Raya–Díaz, S.; Quesada–Moraga, E.; Barrón, V.; del Campillo, M.C.; Sánchez–Rodríguez, A.R. Redefining the Dose of the Entomopathogenic Fungus Metarhizium brunneum (Ascomycota, Hypocreales) to Increase Fe Bioavailability and Promote Plant Growth in Calcareous and Sandy Soils. Plant Soil 2017, 418, 387–404. [Google Scholar] [CrossRef]
- Romera, F.J.; García, M.J.; Lucena, C.; Martínez-Medina, A.; Aparicio, M.A.; Ramos, J.; Alcántara, E.; Angulo, M.; Pérez-Vicente, R. Induced Systemic Resistance (ISR) and Fe Deficiency Responses in Dicot Plants. Front. Plant Sci. 2019, 10, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraemer, S.M. Iron Oxide Dissolution and Solubility in the Presence of Siderophores. Aquat. Sci. 2004, 66, 3–18. [Google Scholar] [CrossRef] [Green Version]
- Krasnoff, S.B.; Keresztes, I.; Donzelli, B.G.G.; Gibson, D.M. Metachelins, Mannosylated and N-Oxidized Coprogen-Type Siderophores from Metarhizium robertsii. J. Nat. Prod. 2014, 77, 1685–1692. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal Chemical Assay for the Detection and Determination of Siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Winkelmann, G. Ecology of Siderophores with Special Reference to the Fungi. BioMetals 2007, 20, 379. [Google Scholar] [CrossRef]
- García, M.J.; Romera, F.J.; Lucena, C.; Alcántara, E.; Pérez-Vicente, R. Ethylene and the Regulation of Physiological and Morphological Responses to Nutrient Deficiencies. Plant Physiol. 2015, 169, 51–60. [Google Scholar] [CrossRef] [Green Version]
- García, M.J.; Angulo, M.; García, C.; Lucena, C.; Alcántara, E.; Pérez-Vicente, R.; Romera, F.J. Influence of Ethylene Signaling in the Crosstalk Between Fe, S, and P Deficiency Responses in Arabidopsis thaliana. Front. Plant Sci. 2021, 12, 643585. [Google Scholar] [CrossRef]
- Gattullo, C.E.; Pii, Y.; Allegretta, I.; Medici, L.; Cesco, S.; Mimmo, T.; Terzano, R. Iron Mobilization and Mineralogical Alterations Induced by Iron-Deficient Cucumber Plants (Cucumis sativus L.) in a Calcareous Soil. Pedosphere 2018, 28, 59–69. [Google Scholar] [CrossRef]
- Rodríguez-Celma, J.; Schmidt, W. Reduction-Based Iron Uptake Revisited. Plant Signal. Behav. 2013, 8, e26116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fourcroy, P.; Sisó-Terraza, P.; Sudre, D.; Savirón, M.; Reyt, G.; Gaymard, F.; Abadía, A.; Abadia, J.; Álvarez-Fernández, A.; Briat, J. Involvement of the <scp>ABCG</Scp> 37 Transporter in Secretion of Scopoletin and Derivatives by Arabidopsis Roots in Response to Iron Deficiency. New Phytol. 2014, 201, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Schmid, N.B.; Giehl, R.F.H.; Doll, S.; Mock, H.-P.; Strehmel, N.; Scheel, D.; Kong, X.; Hider, R.C.; von Wiren, N. Feruloyl-CoA 6’-Hydroxylase1-Dependent Coumarins Mediate Iron Acquisition from Alkaline Substrates in Arabidopsis. Plant Physiol. 2014, 164, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Sisó-Terraza, P.; Luis-Villarroya, A.; Fourcroy, P.; Briat, J.-F.; Abadía, A.; Gaymard, F.; Abadía, J.; Álvarez-Fernández, A. Accumulation and Secretion of Coumarinolignans and Other Coumarins in Arabidopsis thaliana Roots in Response to Iron Deficiency at High PH. Front. Plant Sci. 2016, 7, 1711. [Google Scholar] [CrossRef] [Green Version]
- Sisó-Terraza, P.; Rios, J.J.; Abadía, J.; Abadía, A.; Álvarez-Fernández, A. Flavins Secreted by Roots of Iron-deficient Beta vulgaris Enable Mining of Ferric Oxide via Reductive Mechanisms. New Phytol. 2016, 209, 733–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sisó-Terraza, P. Metabolitos Secundarios Exudados Por Raíces de Plantas de Estrategia I En Respuesta a La Deficiencia de Hierro: Caracterización, Transporte y Función. Ph.D. Thesis, Universitat de Lleida, Lleida, Spain, 2017. [Google Scholar]
- Tsai, H.H.; Schmidt, W. Mobilization of Iron by Plant-Borne Coumarins. Trends Plant Sci. 2017, 22, 538–548. [Google Scholar] [CrossRef]
- Römheld, V.; Marschner, H. Evidence for a Specific Uptake System for Iron Phytosiderophores in Roots of Grasses. Plant Physiol. 1986, 80, 175–180. [Google Scholar] [CrossRef] [Green Version]
- Landsberg, E. Function of Rhizodermal Transfer Cells in the Fe Stress Response Mechanism of Capsicum annuum L. Plant Physiol. 1986, 82, 511–517. [Google Scholar] [CrossRef] [Green Version]
- Lucena, C.; Romera, F.J.; García, M.J.; Alcántara, E.; Pérez-Vicente, R. Ethylene Participates in the Regulation of Fe Deficiency Responses in Strategy I Plants and in Rice. Front. Plant Sci. 2015, 6, 1056. [Google Scholar] [CrossRef] [Green Version]
- Venuti, S.; Zanin, L.; Marroni, F.; Franco, A.; Morgante, M.; Pinton, R.; Tomasi, N. Physiological and Transcriptomic Data Highlight Common Features between Iron and Phosphorus Acquisition Mechanisms in White Lupin Roots. Plant Sci. 2019, 285, 110–121. [Google Scholar] [CrossRef]
- García, M.J.; Lucena, C.; Romera, F.J.; Alcántara, E.; Pérez-Vicente, R. Ethylene and Nitric Oxide Involvement in the Up-Regulation of Key Genes Related to Iron Acquisition and Homeostasis in Arabidopsis. J. Exp. Bot. 2010, 61, 3885–3899. [Google Scholar] [CrossRef] [PubMed]
- García, M.J.; Suárez, V.; Romera, F.J.; Alcántara, E.; Pérez-Vicente, R. A New Model Involving Ethylene, Nitric Oxide and Fe to Explain the Regulation of Fe-Acquisition Genes in Strategy I Plants. Plant Physiol. Biochem. 2011, 49, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Nishizawa, N.K. Iron Uptake, Translocation, and Regulation in Higher Plants. Annu. Rev. Plant Biol. 2012, 63, 131–152. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Robe, K.; Gaymard, F.; Izquierdo, E.; Dubos, C. The Transcriptional Control of Iron Homeostasis in Plants: A Tale of BHLH Transcription Factors? Front. Plant Sci. 2019, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, B.; Bauer, P. FIT, a Regulatory Hub for Iron Deficiency and Stress Signaling in Roots, and FIT-Dependent and -Independent Gene Signatures. J. Exp. Bot. 2020, 71, 1694–1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brumbarova, T.; Bauer, P.; Ivanov, R. Molecular Mechanisms Governing Arabidopsis Iron Uptake. Trends Plant Sci. 2015, 20, 124–133. [Google Scholar] [CrossRef]
- Palmer, C.M.; Hindt, M.N.; Schmidt, H.; Clemens, S.; Guerinot, M. lou MYB10 and MYB72 Are Required for Growth under Iron-Limiting Conditions. PLoS Genet. 2013, 9, e1003953. [Google Scholar] [CrossRef] [Green Version]
- Zamioudis, C.; Hanson, J.; Pieterse, C.M.J. β-Glucosidase BGLU42 Is a MYB72-Dependent Key Regulator of Rhizobacteria-Induced Systemic Resistance and Modulates Iron Deficiency Responses in Arabidopsis Roots. New Phytol. 2014, 204, 368–379. [Google Scholar] [CrossRef]
- Zamioudis, C.; Korteland, J.; van Pelt, J.A.; Hamersveld, M.; Dombrowski, N.; Bai, Y.; Hanson, J.; van Verk, M.C.; Ling, H.; Schulze-Lefert, P.; et al. Rhizobacterial Volatiles and Photosynthesis-related Signals Coordinate MYB72 Expression in Arabidopsis Roots during Onset of Induced Systemic Resistance and Iron-deficiency Responses. Plant J. 2015, 84, 309–322. [Google Scholar] [CrossRef] [Green Version]
- Fomina, M.; Hillier, S.; Charnock, J.M.; Melville, K.; Alexander, I.J.; Gadd, G.M. Role of Oxalic Acid Overexcretion in Transformations of Toxic Metal Minerals by Beauveria caledonica. Appl. Environ. Microbiol. 2005, 71, 371–381. [Google Scholar] [CrossRef]
- Ghosh, S.K.; Banerjee, S.; Sengupta, C. Siderophore Production by Antagonistic Fungi. Bioassay, Characterization and Estimation of Siderophores from Some Important Antagonistic Fungi. J. Biopestic. 2017, 10, 105–112. [Google Scholar] [CrossRef]
- Jin, C.W.; Ye, Y.Q.; Zheng, S.J. An Underground Tale: Contribution of Microbial Activity to Plant Iron Acquisition via Ecological Processes. Ann. Bot. 2014, 113, 7–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dijkstra, F.A.; Carrillo, Y.; Pendall, E.; Morgan, J.A. Rhizosphere Priming: A Nutrient Perspective. Front. Microbiol. 2013, 4, 216. [Google Scholar] [CrossRef] [Green Version]
- Resquín-Romero, G.; Garrido-Jurado, I.; Delso, C.; Ríos-Moreno, A.; Quesada-Moraga, E. Transient Endophytic Colonizations of Plants Improve the Outcome of Foliar Applications of Mycoinsecticides against Chewing Insects. J. Invertebr. Pathol. 2016, 136, 23–31. [Google Scholar] [CrossRef]
- Garrido-Jurado, I.; Resquín-Romero, G.; Amarilla, S.P.; Ríos-Moreno, A.; Carrasco, L.; Quesada-Moraga, E. Transient Endophytic Colonization of Melon Plants by Entomopathogenic Fungi after Foliar Application for the Control of Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae). J. Pest Sci. 2017, 90, 319–330. [Google Scholar] [CrossRef]
- Srimathi, K.; Suji, H.A. Siderophores Detection by Using Blue Agar CAS Assay Methods. Int. J. Sci. Res. Biol. Sci. 2018, 5, 180–185. [Google Scholar] [CrossRef] [Green Version]
- Andrews, M.Y.; Santelli, C.M.; Duckworth, O.W. Digital Image Quantification of Siderophores on Agar Plates. Data Brief 2016, 6, 890–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Mas, N.; Cuenca-Medina, M.; Gutiérrez-Sánchez, F.; Quesada-Moraga, E. Bottom-up Effects of Endophytic Beauveria bassiana on Multitrophic Interactions between the Cotton Aphid, Aphis gossypii, and Its Natural Enemies in Melon. J. Pest Sci. 2019, 92, 1271–1281. [Google Scholar] [CrossRef] [Green Version]
- Quesada-Moraga, E.; Yousef-Naef, M.; Garrido-Jurado, I. Advances in the Use of Entomopathogenic Fungi as Biopesticides in Suppressing Crop Insect Pests. In Biopesticides for Sustainable Agriculture; Birch, N.N., Glare, T., Eds.; Burleigh Dodds Science Publishing: Cambridge, UK, 2020; pp. 63–98. [Google Scholar]
- Olsen, S.R.; Cole, S.V.; Watanabe, F.S.; Dean, L.A. Circular No. 939 Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; USDA: Washington, DC, USA, 1954; pp. 1–19.
- Zasoski, R.J.; Burau, R.G. A Rapid Nitric-perchloric Acid Digestion Method for Multi-element Tissue Analysis. Commun. Soil Sci. Plant Anal. 1977, 8, 425–436. [Google Scholar] [CrossRef]
- Bianchini, M.R.; Eyherabide, G.A. Técnicas de Mineralización Para La Determinación de Macronutrientes En Muestras de Raíz de Zanahoria (Daucus carota L.). Rev. Fac. Agron. 1998, 103, 191–195. [Google Scholar]
- Lucena, C.; Waters, B.M.; Romera, F.J.; García, M.J.; Morales, M.; Alcántara, E.; Pérez-Vicente, R. Ethylene Could Influence Ferric Reductase, Iron Transporter, and H +-ATPase Gene Expression by Affecting FER (or FER-like) Gene Activity. J. Exp. Bot. 2006, 57, 4145–4154. [Google Scholar] [CrossRef] [PubMed]
- Römheld, V.; Marschner, H. Iron Deficiency Stress Induced Morphological and Physiological Changes in Root Tips of Sunflower. Physiol. Plant 1981, 53, 354–360. [Google Scholar] [CrossRef]
- García, M.J.; Angulo, M.; Romera, F.J.; Lucena, C.; Pérez-Vicente, R. A Shoot Derived Long Distance Iron Signal May Act Upstream of the IMA Peptides in the Regulation of Fe Deficiency Responses in Arabidopsis thaliana Roots. Front. Plant Sci. 2022, 13, 971773. [Google Scholar] [CrossRef]
- García, M.J.; Romera, F.J.; Stacey, M.G.; Stacey, G.; Villar, E.; Alcántara, E.; Pérez-Vicente, R. Shoot to Root Communication Is Necessary to Control the Expression of Iron-Acquisition Genes in Strategy I Plants. Planta 2013, 237, 65–75. [Google Scholar] [CrossRef]
- Angulo, M.; García, M.J.; Alcántara, E.; Pérez-Vicente, R.; Romera, F.J. Comparative Study of Several Fe Deficiency Responses in the Arabidopsis thaliana Ethylene Insensitive Mutants Ein2-1 and Ein2-5. Plants 2021, 10, 262. [Google Scholar] [CrossRef]
- Hashemi, L.; Golparvar, A.R.; Nasr-Esfahani, M.; Golabadi, M. Expression Analysis of Defense-Related Genes in Cucumber (Cucumis Sativus L.) against Phytophthora melonis. Mol. Biol. Rep. 2020, 47, 4933–4944. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar] [CrossRef]
- Aparicio, M.A.; Lucena, C.; García, M.J.; Ruiz-Castilla, F.J.; Jiménez-Adrián, P.; López-Berges, M.S.; Prieto, P.; Alcántara, E.; Pérez-Vicente, R.; Ramos, J.; et al. The Nonpathogenic Strain of Fusarium Oxysporum FO12 Induces Fe Deficiency Responses in Cucumber (Cucumis sativus L.) Plants. Planta 2023, 257, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Waters, B.M.; McInturf, S.A.; Amundsen, K. Transcriptomic and Physiological Characterization of the Fefe Mutant of Melon (Cucumis melo) Reveals New Aspects of Iron-Copper Crosstalk. New Phytol. 2014, 203, 1128–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moonjely, S.; Barelli, L.; Bidochka, M.J. Insect Pathogenic Fungi as Endophytes. Adv. Genet. 2016, 94, 107–135. [Google Scholar] [CrossRef] [PubMed]
- Brunner-Mendoza, C.; Reyes-Montes, M.d.R.; Moonjely, S.; Bidochka, M.J.; Toriello, C. A Review on the Genus Metarhizium as an Entomopathogenic Microbial Biocontrol Agent with Emphasis on Its Use and Utility in Mexico. Biocontrol Sci. Technol. 2019, 29, 83–102. [Google Scholar] [CrossRef]
- Yousef, M.; Alba-Ramírez, C.; Jurado, I.G.; Mateu, J.; Díaz, S.R.; Valverde-García, P.; Quesada-Moraga, E. Metarhizium brunneum (Ascomycota; Hypocreales) Treatments Targeting Olive Fly in the Soil for Sustainable Crop Production. Front. Plant Sci. 2018, 9, 1. [Google Scholar] [CrossRef] [Green Version]
- Stone, L.B.L.; Bidochka, M.J. The Multifunctional Lifestyles of Metarhizium: Evolution and Applications. Appl. Microbiol. Biotechnol. 2020, 104, 9935–9945. [Google Scholar] [CrossRef]
- Agbessenou, A.; Akutse, K.S.; Yusuf, A.A.; Ekesi, S.; Subramanian, S.; Khamis, F.M. Endophytic Fungi Protect Tomato and Nightshade Plants against Tuta Absoluta (Lepidoptera: Gelechiidae) through a Hidden Friendship and Cryptic Battle. Sci. Rep. 2020, 10, 22195. [Google Scholar] [CrossRef] [PubMed]
- Moonjely, S.; Bidochka, M.J. Generalist and Specialist Metarhizium Insect Pathogens Retain Ancestral Ability to Colonize Plant Roots. Fungal Ecol. 2019, 41, 209–217. [Google Scholar] [CrossRef]
- Miranda-Fuentes, P.; Yousef-Yousef, M.; Valverde-García, P.; Rodríguez-Gómez, I.M.; Garrido-Jurado, I.; Quesada-Moraga, E. Entomopathogenic Fungal Endophyte-Mediated Tritrophic Interactions between Spodoptera littoralis and Its Parasitoid Hyposoter didymator. J. Pest Sci. 2021, 94, 933–945. [Google Scholar] [CrossRef]
- Quesada-Moraga, E.; Garrido-Jurado, I.; Yousef-Yousef, M.; González-Mas, N. Multitrophic Interactions of Entomopathogenic Fungi in BioControl. BioControl 2022, 67, 457–472. [Google Scholar] [CrossRef]
- Sánchez-Montesinos, B.; Diánez, F.; Moreno-Gavíra, A.; Gea, F.J.; Santos, M. Role of Trichoderma aggressivum f. Europaeum as Plant-growth Promoter in Horticulture. Agronomy 2020, 10, 1004. [Google Scholar] [CrossRef]
- Pal, S.; Ghosh, S.K. Phosphate Solubilization Study of Entomopathogenic Fungi Beauveria bassiana (Bals.) Vuill. and Its Effects on Brinjal Plant (Solanum melongena L.) Growth. In Biotechnology and Nature; Dulay, D., Sovan, R., Gopal, C.B., Eds.; Kabitika: Paschim Msdinipur, India, 2018; pp. 109–121. [Google Scholar]
- Naeem, U.; Haq, I.; Afzaal, M.; Qazi, A.; Yasar, A.; Tabinda, A.; Mahfooz, Y.; Naz, A.U.; Awan, H. Investigating the Effect of Aspergillus niger Inoculated Press Mud (Biofertilizer) on the Potential of Enhancing Maize (Zea mays L.) Yield, Potassium Use Efficiency and Potassium Agronomic Efficiency. Cereal Res. Commun. 2022, 50, 157–170. [Google Scholar] [CrossRef]
- Baron, N.C.; Costa, N.T.A.; Mochi, D.A.; Rigobelo, E.C. First Report of Aspergillus sydowii and Aspergillus brasiliensis as Phosphorus Solubilizers in Maize. Ann. Microbiol. 2018, 68, 863–870. [Google Scholar] [CrossRef]
- Baron, N.C.; de Souza Pollo, A.; Rigobelo, E.C. Purpureocillium lilacinum and Metarhizium marquandii as Plant Growth-Promoting Fungi. PeerJ 2020, 8, e9005. [Google Scholar] [CrossRef]
- Marschner, H.; Römheld, V.; Kissel, M. Different Strategies in Higher Plants in Mobilization and Uptake of Iron. J. Plant Nutr. 1986, 9, 695–713. [Google Scholar] [CrossRef]
- Brown, J.C.; Jolley, V.D. Strategy I and Strategy II Mechanisms Affecting Iron Availability to Plants May Be Established Too Narrow or Limited. J. Plant Nutr. 1988, 11, 1077–1098. [Google Scholar] [CrossRef]
- Marschner, H.; Römheld, V. Strategies of Plants for Acquisition of Iron. Plant Soil 1994, 165, 261–274. [Google Scholar] [CrossRef]
- Ivanov, R.; Brumbarova, T.; Bauer, P. Fitting into the Harsh Reality: Regulation of Iron-Deficiency Responses in Dicotyledonous Plants. Mol. Plant 2012, 5, 27–42. [Google Scholar] [CrossRef] [Green Version]
- García, M.J.; Lucena, C.; Romera, F.J. Ethylene and Nitric Oxide Involvement in the Regulation of Fe and P Deficiency Responses in Dicotyledonous Plants. Int. J. Mol. Sci. 2021, 22, 4904. [Google Scholar] [CrossRef]
- Naranjo-Arcos, M.A.; Bauer, P. Iron Nutrition, Oxidative Stress, and Pathogen Defense. In Nutritional Deficiency; Erkekoglu, P., Kocer-Gumusel, B., Eds.; InTechOpen: London, UK, 2016; pp. 63–98. [Google Scholar]
- Vera, D.F.; Perez, H.; Hernando, V. Isolation of Phosphate Solubilizer Fungi from Araza Rhizosphere. Acta Biol. Colomb. 2002, 7, 33. [Google Scholar]
- Zhao, L.; Wang, Y.; Kong, S. Effects of Trichoderma asperellum and Its Siderophores on Endogenous Auxin in Arabidopsis Thaliana under Iron-Deficiency Stress. Int. Microbiol. 2020, 23, 501–509. [Google Scholar] [CrossRef]
- Pii, Y.; Borruso, L.; Brusetti, L.; Crecchio, C.; Cesco, S.; Mimmo, T. The Interaction between Iron Nutrition, Plant Species and Soil Type Shapes the Rhizosphere Microbiome. Plant Physiol. Biochem. 2016, 99, 39–48. [Google Scholar] [CrossRef]
- Zhou, C.; Guo, J.; Zhu, L.; Xiao, X.; Xie, Y.; Zhu, J.; Ma, Z.; Wang, J. Paenibacillus polymyxa BFKC01 Enhances Plant Iron Absorption via Improved Root Systems and Activated Iron Acquisition Mechanisms. Plant Physiol. Biochem. 2016, 105, 162–173. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, Y.; Xie, X.; Kim, M.-S.; Dowd, S.E.; Paré, P.W. A Soil Bacterium Regulates Plant Acquisition of Iron via Deficiency-Inducible Mechanisms. Plant J. 2009, 58, 568–577. [Google Scholar] [CrossRef]
- Kabir, A.H.; Debnath, T.; Das, U.; Prity, S.A.; Haque, A.; Rahman, M.M.; Parvez, M.S. Arbuscular Mycorrhizal Fungi Alleviate Fe-Deficiency Symptoms in Sunflower by Increasing Iron Uptake and Its Availability along with Antioxidant Defense. Plant Physiol. Biochem. 2020, 150, 254–262. [Google Scholar] [CrossRef]
- Rahman, M.A.; Parvin, M.; Das, U.; Ela, E.J.; Lee, S.-H.; Lee, K.-W.; Kabir, A.H. Arbuscular Mycorrhizal Symbiosis Mitigates Iron (Fe)-Deficiency Retardation in Alfalfa (Medicago sativa L.) Through the Enhancement of Fe Accumulation and Sulfur-Assisted Antioxidant Defense. Int. J. Mol. Sci. 2020, 21, 2219. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Wang, F.; Zhang, Y.; Zhang, J. Involvement of Trichoderma asperellum Strain T6 in Regulating Iron Acquisition in Plants. J. Basic Microbiol. 2014, 54, S115–S124. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; van Wees, S.C.M.; Pieterse, C.M.J. Airborne Signals from Trichoderma Fungi Stimulate Iron Uptake Responses in Roots Resulting in Priming of Jasmonic Acid-Dependent Defences in Shoots of Arabidopsis thaliana and Solanum lycopersicum. Plant Cell Environ. 2017, 40, 2691–2705. [Google Scholar] [CrossRef] [Green Version]
- Lucena, C.; Alcalá-Jiménez, M.T.; Romera, F.J.; Ramos, J. Several Yeast Species Induce Iron Deficiency Responses in Cucumber Plants (Cucumis sativus L.). Microorganisms 2021, 9, 2603. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Fungal Species | Origin | Agroecosystem | Habitat | GenBank Accession Number | Spanish Type Culture Collection Accession Number |
|---|---|---|---|---|---|---|
| EABb 04/01-Tip | B. bassiana | Ecija (Sevilla, Spain) | Opium poppy crop | Insect (Iraella luteipes) | FJ972963 | 20744 |
| EABb 01/33-Su | B. bassiana | El Bosque (Cadiz, Spain) | Traditional olive orchard | Soil | FJ972969 | 21149 |
| EAMa 01/58–Su | M. brunneum | Hinojosa del Duque (Córdoba, Spain) | Wheat crop | Soil | JN900390 | 20764 |
| Gene | Gene Function/Name | Accession No. | Reference | Sequence | Species | Tissue |
|---|---|---|---|---|---|---|
| FRO1 | Ferric reductase oxidase | AY590765 | [61] | F: ATACGGCCCTGTTTCCACTT R: GGGTTTTGTTGTGGTGGGAA | C. sativus | Roots |
| FRO1 | Ferric reductase oxidase | [62] | F: TCACAGCGATTTAGAACCAGA R: GCCTTCGAGGGAAACTTGAA | C. melo | Roots | |
| FRO2 | Ferric reductase oxidase | [62] | F: TCTATCTAATCCATGTGGGAGTAGC R: AACAGCGCCAGAAGGAAGAT | C. melo | Roots | |
| FRO3 | Ferric reductase oxidase | [62] | F: CGAAGGCTGAAGTATAAACCAAC R: ACCTTGTCCATGACTCATCACA | C. melo | Roots/Shoots | |
| FRO4 | Ferric reductase oxidase | [62] | F: CACCGTCGAATTGGTCCT R: TGGACTCGACGACACACTGAA | C. melo | Roots/Shoots | |
| IRT1 | Iron-Regulated Transporter1 | AY590764 | [61] | F: GCAGGTATCATTCTCGCCAC R: ATCATAGCAACGAAGCCCGA | C. sativus | Roots |
| IRT1 | Iron-Regulated Transporter1 | [62] | F: ATCCCAATGTTGCACCCGGATAGA R: AAACCGGTGGCGAGAATGATACCT | C. melo | Roots | |
| HA1 | ATPase | AJ703810 | [61] | F: GGGATGGGCTGGTGTAGTTTG R: TTCTTGGTCGTAAAGGCGGT | C. sativus | Roots |
| FIT | Induced Transcription Factor | [62] | F: GACATCAACGATCAATTTGAG R: CGATCCTCGATCAAGCAA | C. melo/C. sativus | Roots | |
| Actin * | Actina | XM_004136807 | [61] | F: AACCCAAAGGCAAACAGGGA R: TCCGACCACTGGCATAGAGA | C. melo/C. sativus | Roots/Shoots |
| Cyclo * | Cyclophilin | NM_001280769 | [61] | F: ATTTCCTATTTGCGTGTGTTGTT R: GTAGCATAAACCATGACCCATAATA | C. melo/C. sativus | Roots/Shoots |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Espinoza, F.; Quesada-Moraga, E.; García del Rosal, M.J.; Yousef-Yousef, M. Entomopathogenic Fungi-Mediated Solubilization and Induction of Fe Related Genes in Melon and Cucumber Plants. J. Fungi 2023, 9, 258. https://doi.org/10.3390/jof9020258
García-Espinoza F, Quesada-Moraga E, García del Rosal MJ, Yousef-Yousef M. Entomopathogenic Fungi-Mediated Solubilization and Induction of Fe Related Genes in Melon and Cucumber Plants. Journal of Fungi. 2023; 9(2):258. https://doi.org/10.3390/jof9020258
Chicago/Turabian StyleGarcía-Espinoza, Fabián, Enrique Quesada-Moraga, María José García del Rosal, and Meelad Yousef-Yousef. 2023. "Entomopathogenic Fungi-Mediated Solubilization and Induction of Fe Related Genes in Melon and Cucumber Plants" Journal of Fungi 9, no. 2: 258. https://doi.org/10.3390/jof9020258