
Comparative Mitogenomic Analysis Reveals Intraspecific, Interspecific Variations and Genetic Diversity of Medical Fungus Ganoderma

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequencing, Assembly, and Annotation of Mitogenomes
2.2. Sequence Analysis
2.3. Intron Analysis
2.4. Phylogenetic Analysis
3. Results
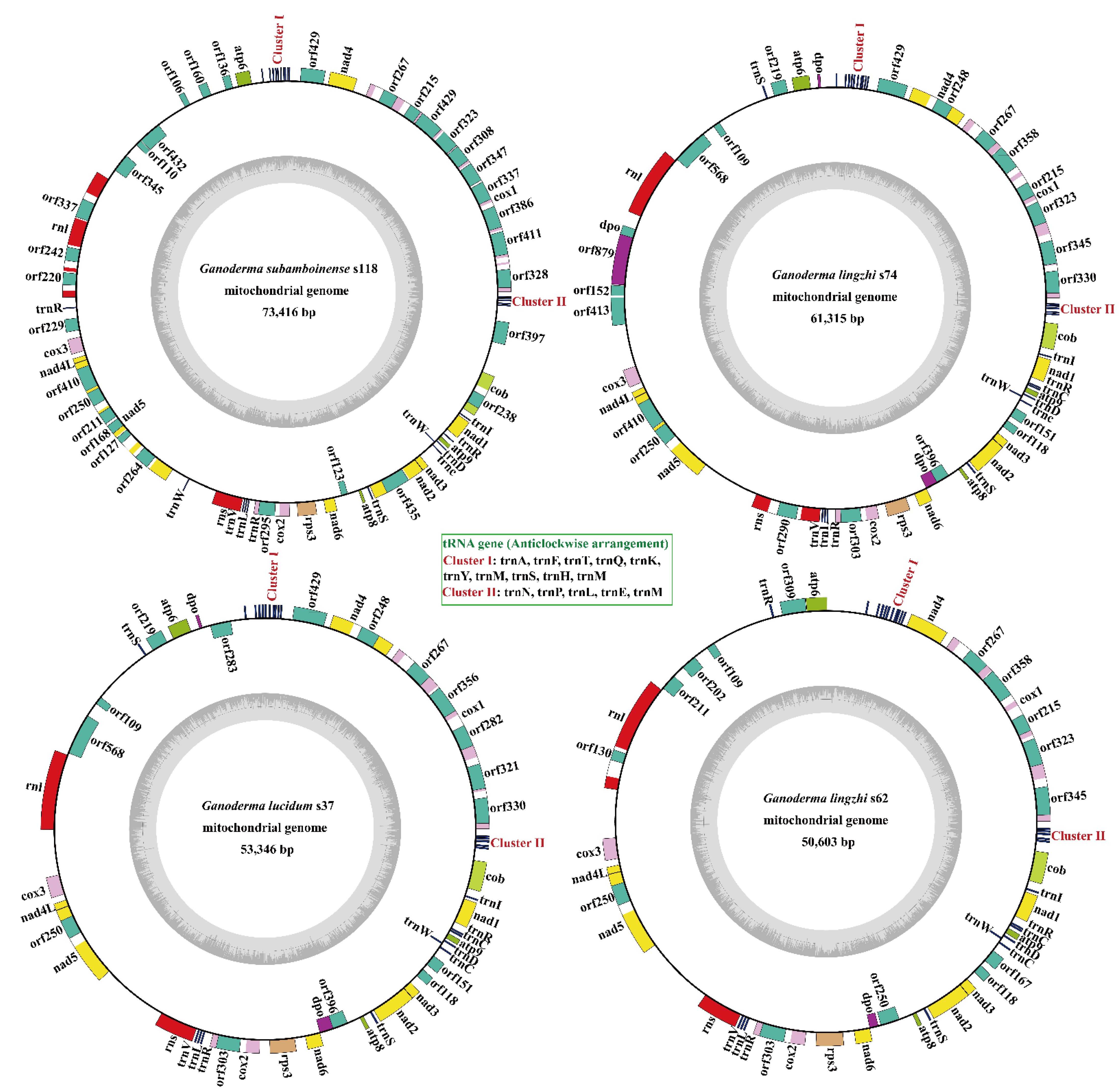
3.1. Characterization of the Four Ganoderma Mitogenomes
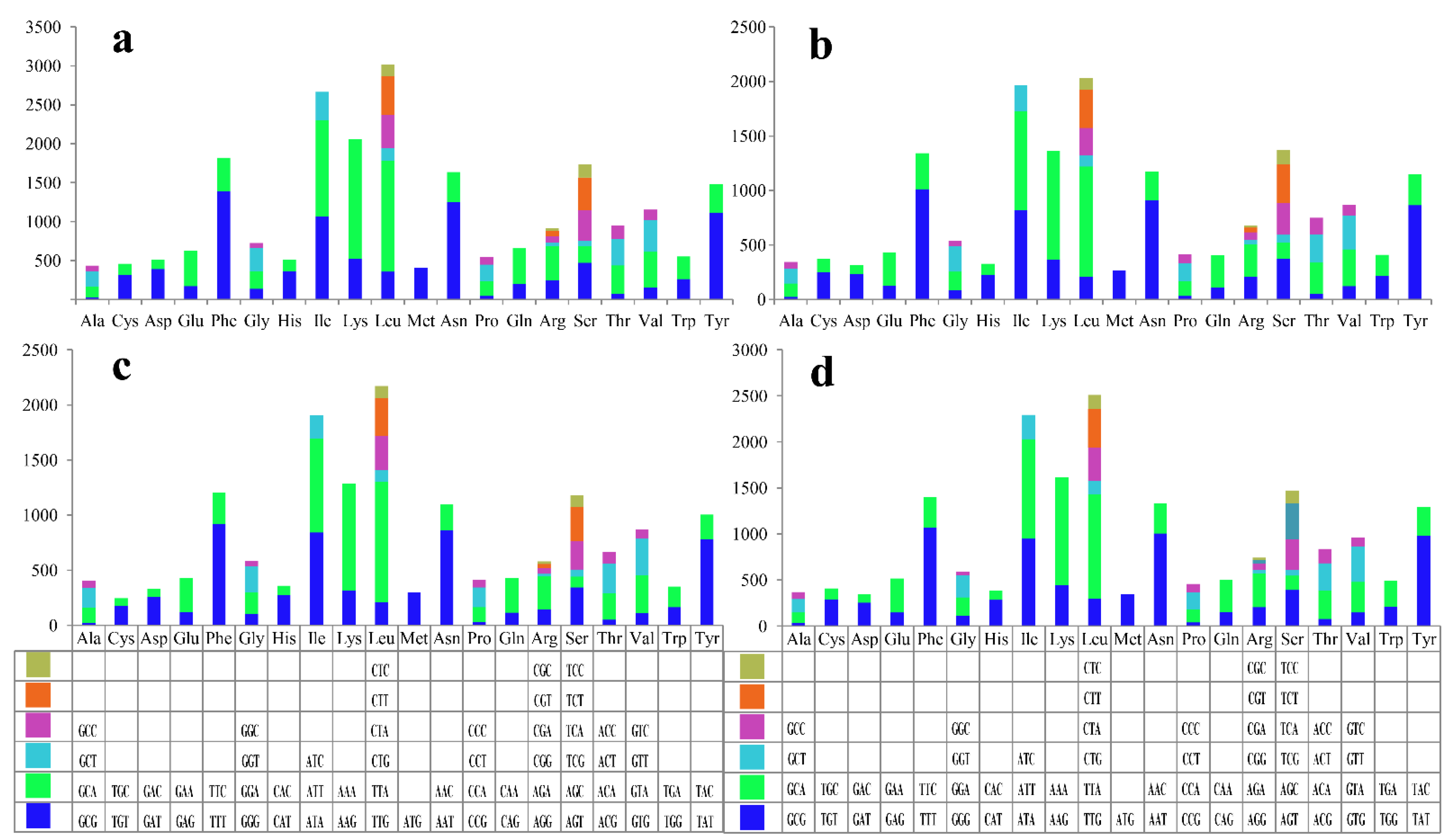
3.2. Codon Usage Analysis
3.3. Repetitive Sequences Analysis
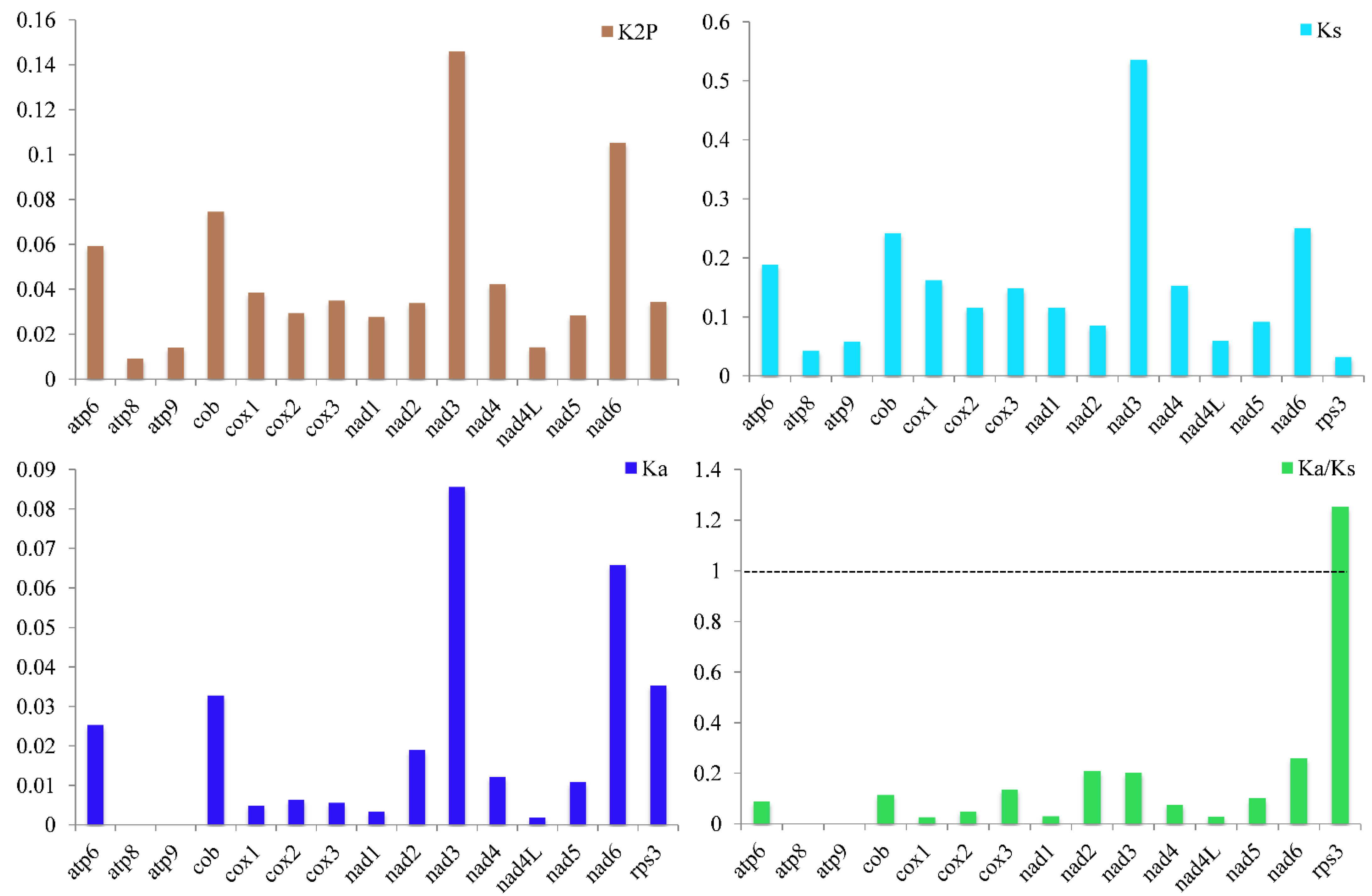
3.4. Variation, Genetic Distance, and Evolutionary Rates of Core Genes
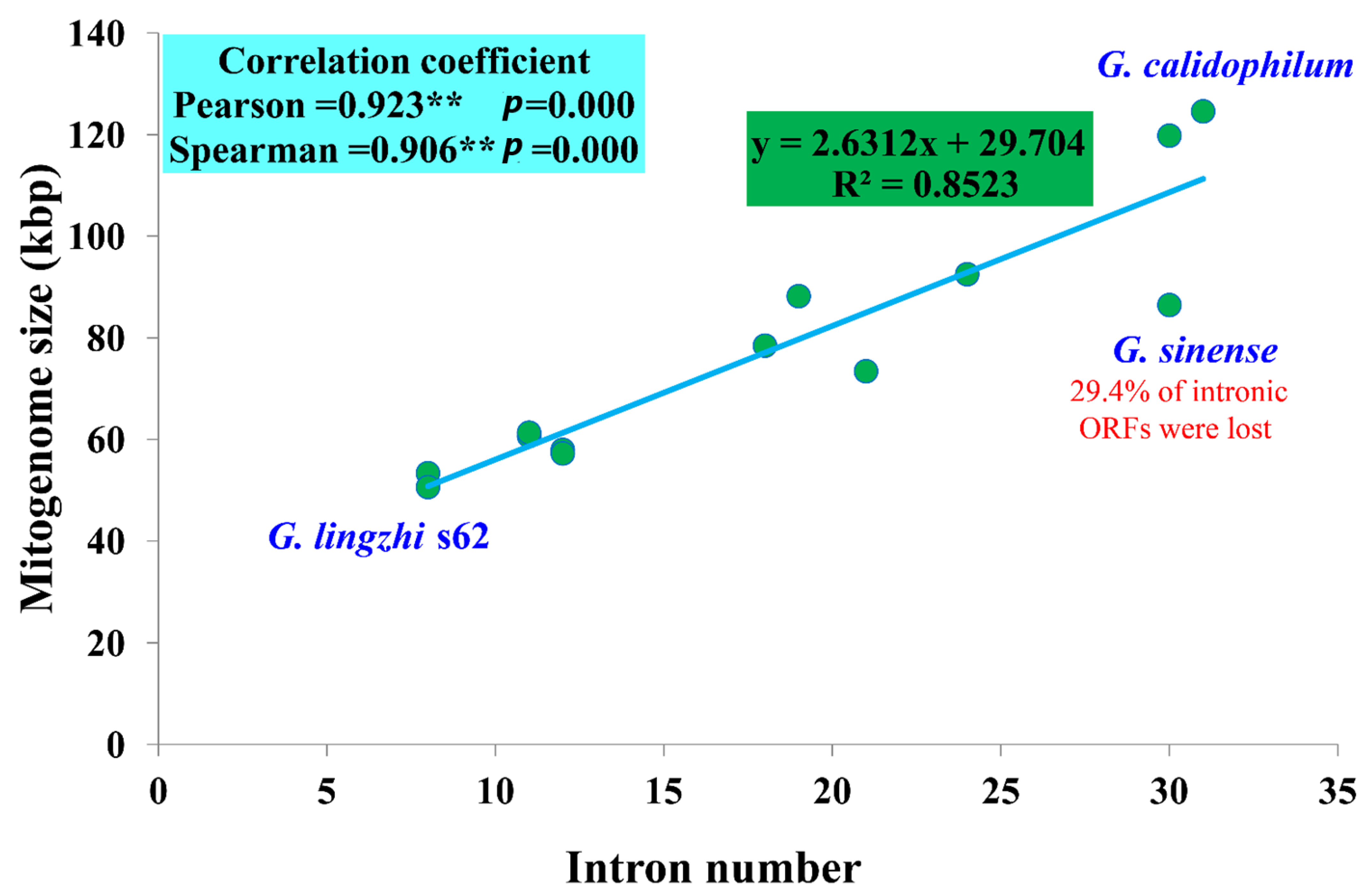
3.5. Intron Dynamic Analysis
3.6. Comparative Mitogenomic Analysis
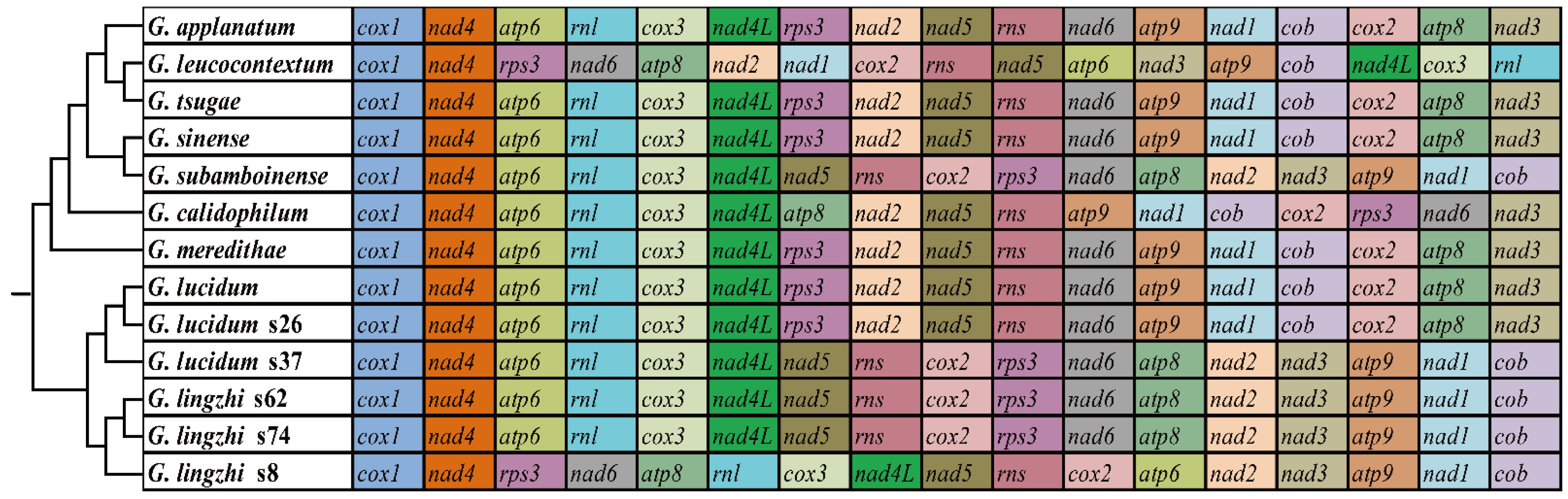
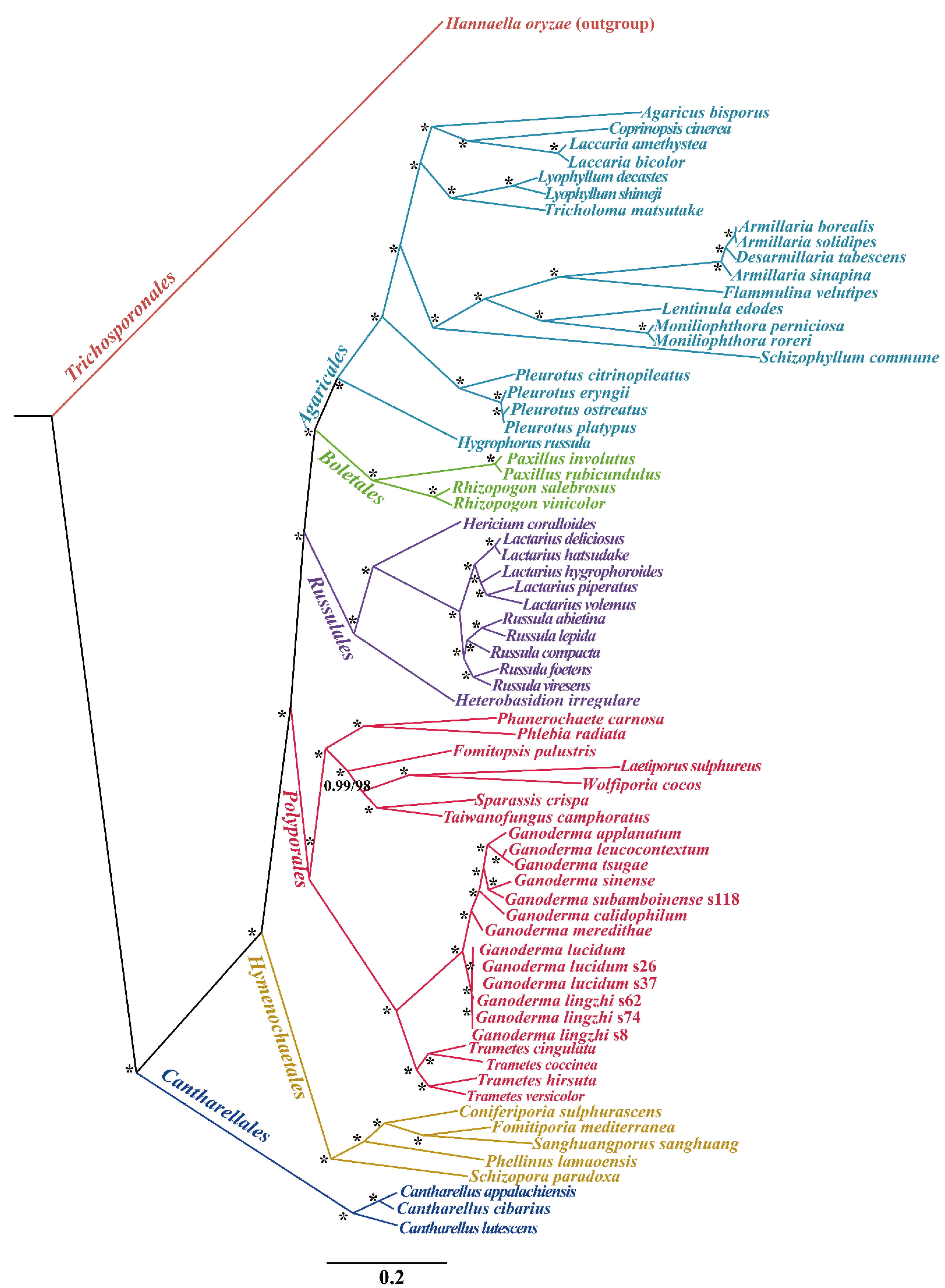
3.7. Gene Arrangement and Phylogenetic Analyses
4. Discussion
4.1. Mitogenome Size Variation in Ganoderma
4.2. Mitogenome Content Evolution in Ganoderma
4.3. Dynamics of Introns in Ganoderma
4.4. Gene Rearrangements and Phylogenetic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Kwon, O.-C.; Park, Y.-J.; Kim, H.-I.; Kong, W.-S.; Cho, J.-H.; Lee, C.-S. Taxonomic Position and Species Identity of the Cultivated Yeongji ‘Ganoderma lucidum’ in Korea. Mycobiology 2016, 44, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jargalmaa, S.; Eimes, J.A.; Park, M.S.; Park, J.Y.; Oh, S.-Y.; Lim, Y.W. Taxonomic evaluation of selected Ganoderma species and database sequence validation. PeerJ 2017, 5, e3596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kües, U.; Nelson, D.R.; Liu, C.; Yu, G.-J.; Zhang, J.; Li, J.; Wang, X.-C.; Sun, H. Genome analysis of medicinal Ganoderma spp. with plant-pathogenic and saprotrophic life-styles. Phytochemistry 2015, 114, 18–37. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.-F.; Xing, J.-H.; He, X.-L.; Wu, D.-M.; Song, C.-G.; Liu, S.; Vlasák, J.; Gates, G.; Gibertoni, T.; Cui, B.-K. Species diversity, systematic revision and molecular phylogeny of Ganodermataceae (Polyporales, Basidiomycota) with an emphasis on Chinese collections. Stud. Mycol. 2022, 101, 287–415. [Google Scholar] [CrossRef]
- Wang, X.-C.; Xi, R.-J.; Li, Y.; Wang, N.-M.; Yao, Y.-J. The Species Identity of the Widely Cultivated Ganoderma, ‘G. lucidum’ (Ling-zhi), in China. PLoS ONE 2012, 7, e40857. [Google Scholar] [CrossRef] [Green Version]
- Dhillon, B.; Hamelin, R.C.; Rollins, J.A. Transcriptional profile of oil palm pathogen, Ganoderma boninense, reveals activation of lignin degradation machinery and possible evasion of host immune response. BMC Genom. 2021, 22, 326. [Google Scholar] [CrossRef]
- Hidayati, N.; Glen, M.; Nurrohmah, S.H.; Rimbawanto, A.; Mohammed, C.L. Ganoderma steyaertanum as a root-rot pathogen of forest trees. For. Pathol. 2014, 44, 460–471. [Google Scholar] [CrossRef]
- Page, D.E.; Glen, M.; Puspitasari, D.; Rimbawanto, A.; Ratkowsky, D.; Mohammed, C. Sexuality and mating types of Ganoderma philippii, Ganoderma mastoporum and Ganoderma australe, three basidiomycete fungi with contrasting ecological roles in south-east Asian pulpwood plantations. Australas. Plant Pathol. 2017, 47, 83–94. [Google Scholar] [CrossRef]
- Francis, A.; Beadle, C.; Puspitasari, D.; Irianto, R.; Agustini, L.; Rimbawanto, A.; Gafur, A.; Hardiyanto, E.; Junarto, N.; Hidyati, B.; et al. Disease progression in plantations of Acacia mangium affected by red root rot (Ganoderma philippii). For. Pathol. 2014, 44, 447–459. [Google Scholar] [CrossRef]
- Bishop, K.S.; Kao, C.H.; Xu, Y.; Glucina, M.P.; Paterson, R.R.M.; Ferguson, L.R. From 2000 years of Ganoderma lucidum to recent developments in nutraceuticals. Phytochemistry 2015, 114, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Paterson, R.R.M. Ganoderma—A therapeutic fungal biofactory. Phytochemistry 2006, 67, 1985–2001. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Xu, J.; Sun, C.; Zhou, S.; Xu, H.; Nelson, D.R.; Qian, J.; Song, J.; Luo, H.; Xiang, L.; et al. Chromosome-level genome map provides insights into diverse defense mechanisms in the medicinal fungus Ganoderma sinense. Sci. Rep. 2015, 5, 11087. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.S.; Asatiani, M.D.; Sharvit, L.E.; Trabelcy, B.; Barseghyan, G.S.; Wasser, S.P. Chemical Composition and Medicinal Value of the New Ganoderma tsugae var. jannieae CBS-120304 Medicinal Higher Basidiomycete Mushroom. Int. J. Med. Mushrooms 2015, 17, 735–747. [Google Scholar] [CrossRef]
- Chien, R.C.; Tsai, S.Y.; Lai, E.Y.; Mau, J.L. Antiproliferative Activities of Hot Water Extracts from Culinary-Medicinal Mush-rooms, Ganoderma tsugae and Agrocybe cylindracea (Higher Basidiomycetes) on Cancer Cells. Int. J. Med. Mushrooms 2015, 17, 453–462. [Google Scholar] [CrossRef]
- Dai, Y.-C.; Zhou, L.-W.; Hattori, T.; Cao, Y.; Stalpers, J.A.; Ryvarden, L.; Buchanan, P.; Oberwinkler, F.; Hallenberg, N.; Liu, P.-G.; et al. Ganoderma Lingzhi (Polyporales, Basidiomycota): The scientific binomial for the widely cultivated medicinal fungus Lingzhi. Mycol. Prog. 2017, 16, 1051–1055. [Google Scholar] [CrossRef]
- Dai, Y.C.; Yang, Z.L.; Cui, B.K.; Wu, G.; Yuan, H.S.; Zhou, L.W.; He, S.H.; Ge, Z.W.; Wu, F.; Wei, Y.L.; et al. Diversity and sys-tematics of the important macrofungi in Chinese forests. Mycosystema 2021, 40, 770–805. [Google Scholar] [CrossRef]
- Gao, X.; Qi, J.; Ho, C.-T.; Li, B.; Mu, J.; Zhang, Y.; Hu, H.; Mo, W.; Chen, Z.; Xie, Y. Structural characterization and immunomodulatory activity of a water-soluble polysaccharide from Ganoderma leucocontextum fruiting bodies. Carbohydr. Polym. 2020, 249, 116874. [Google Scholar] [CrossRef]
- Zheng, S.; Zhu, N.; Shi, C. Genomic data mining approaches for the discovery of anticancer peptides from Ganoderma sinense. Phytochemistry 2020, 179, 112466. [Google Scholar] [CrossRef]
- Liao, B.; Chen, X.; Han, J.; Dan, Y.; Wang, L.; Jiao, W.; Song, J.; Chen, S. Identification of commercial Ganoderma (Lingzhi) species by ITS2 sequences. Chin. Med. 2015, 10, 22. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.-M.; Wu, S.-H.; Yao, Y.-J. Clarification of the Concept of Ganoderma orbiforme with High Morphological Plasticity. PLoS ONE 2014, 9, e98733. [Google Scholar] [CrossRef]
- Cao, Y.; Wu, S.-H.; Dai, Y.-C. Species clarification of the prize medicinal Ganoderma mushroom “Lingzhi”. Fungal Divers. 2012, 56, 49–62. [Google Scholar] [CrossRef]
- Cameron, S.L. Insect Mitochondrial Genomics: Implications for Evolution and Phylogeny. Annu. Rev. Èntomol. 2014, 59, 95–117. [Google Scholar] [CrossRef] [Green Version]
- Yuan, M.-L.; Zhang, Q.-L.; Guo, Z.-L.; Wang, J.; Shen, Y.-Y. Comparative mitogenomic analysis of the superfamily Pentatomoidea (Insecta: Hemiptera: Heteroptera) and phylogenetic implications. BMC Genomics 2015, 16, 460. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhang, J.; Chen, H.; Chen, X.; Lan, J.; Liu, C. Complete Mitochondrial Genome of the Medicinal Mushroom Ganoderma lucidum. PLoS ONE 2013, 8, e72038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Xiang, D.; Wan, Y.; Wu, Q.; Wu, X.; Ma, C.; Song, Y.; Zhao, G.; Huang, W. The complete mitochondrial genomes of five important medicinal Ganoderma species: Features, evolution, and phylogeny. Int. J. Biol. Macromol. 2019, 139, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-C.; Shao, J.; Liu, C. The complete mitochondrial genome of the medicinal fungus Ganoderma applanatum (Polyporales, Basidiomycota). Mitochondrial DNA Part A 2015, 27, 2813–2814. [Google Scholar] [CrossRef]
- Wang, X.-C.; Wu, K.; Chen, H.; Shao, J.; Zhang, N.; Chen, X.; Lan, J.; Liu, C. The complete mitochondrial genome of the white-rot fungus Ganoderma meredithiae (Polyporales, Basidiomycota). Mitochondrial DNA Part A 2015, 27, 4197–4198. [Google Scholar] [CrossRef] [PubMed]
- Latorre-Pellicer, A.; Moreno-Loshuertos, R.; Lechuga-Vieco, A.V.; Sánchez-Cabo, F.; Torroja, C.; Acín-Pérez, R.; Calvo, E.; Aix, E.; González-Guerra, A.; Logan, A.; et al. Mitochondrial and nuclear DNA matching shapes metabolism and healthy ageing. Nature 2016, 535, 561–565. [Google Scholar] [CrossRef]
- Boore, J.L. Animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef] [Green Version]
- Cieslak, M.; Pruvost, M.; Benecke, N.; Hofreiter, M.; Morales, A.; Reissmann, M.; Ludwig, A. Origin and History of Mitochondrial DNA Lineages in Domestic Horses. PLoS ONE 2010, 5, e15311. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Ren, Y.; Shi, X.; Peng, L.; Zhao, J.; Song, Y.; Zhao, G. Comparative Mitochondrial Genome Analysis of Two Ectomycorrhizal Fungi (Rhizopogon) Reveals Dynamic Changes of Intron and Phylogenetic Relationships of the Subphylum Agaricomycotina. Int. J. Mol. Sci. 2019, 20, 5167. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Cheng, J.; Ren, Y.; Liao, W.; Li, Q. The First Mitochondrial Genome for Geastrales (Sphaerobolus stellatus) Reveals Intron Dynamics and Large-Scale Gene Rearrangements of Basidiomycota. Front. Microbiol. 2020, 11, 1970. [Google Scholar] [CrossRef]
- Zhang, J.-Y.; Zhang, L.-P.; Yu, D.-N.; Storey, K.B.; Zheng, R.-Q. Complete mitochondrial genomes of Nanorana taihangnica and N. yunnanensis (Anura: Dicroglossidae) with novel gene arrangements and phylogenetic relationship of Dicroglossidae. BMC Evol. Biol. 2018, 18, 26. [Google Scholar] [CrossRef] [Green Version]
- Barr, C.M.; Neiman, M.; Taylor, D.R. Inheritance and recombination of mitochondrial genomes in plants, fungi and animals. New Phytol. 2005, 168, 39–50. [Google Scholar] [CrossRef]
- Freel, K.C.; Friedrich, A.; Schacherer, J. Mitochondrial genome evolution in yeasts: An all-encompassing view. FEMS Yeast Res. 2015, 15, fov023. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wang, Q.; Jin, X.; Chen, Z.; Xiong, C.; Li, P.; Liu, Q.; Huang, W. Characterization and comparative analysis of six complete mitochondrial genomes from ectomycorrhizal fungi of the Lactarius genus and phylogenetic analysis of the Agaricomycetes. Int. J. Biol. Macromol. 2018, 121, 249–260. [Google Scholar] [CrossRef]
- Li, Q.; Wang, Q.; Chen, C.; Jin, X.; Chen, Z.; Xiong, C.; Li, P.; Zhao, J.; Huang, W. Characterization and comparative mitogenomic analysis of six newly sequenced mitochondrial genomes from ectomycorrhizal fungi (Russula) and phylogenetic analysis of the Agaricomycetes. Int. J. Biol. Macromol. 2018, 119, 792–802. [Google Scholar] [CrossRef]
- Li, Q.; Wu, P.; Li, L.; Feng, H.; Tu, W.; Bao, Z.; Xiong, C.; Gui, M.; Huang, W. The first eleven mitochondrial genomes from the ectomycorrhizal fungal genus (Boletus) reveal intron loss and gene rearrangement. Int. J. Biol. Macromol. 2021, 172, 560–572. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-J.; Yang, X.-Q.; Zhang, S.; Humber, R.A.; Xu, J. Genomic analyses reveal low mitochondrial and high nuclear diversity in the cyclosporin-producing fungus Tolypocladium inflatum. Appl. Microbiol. Biotechnol. 2017, 101, 8517–8531. [Google Scholar] [CrossRef]
- Zhang, Y.-J.; Zhang, H.-Y.; Liu, X.-Z.; Zhang, S. Mitochondrial genome of the nematode endoparasitic fungus Hirsutella vermicola reveals a high level of synteny in the family Ophiocordycipitaceae. Appl. Microbiol. Biotechnol. 2017, 101, 3295–3304. [Google Scholar] [CrossRef]
- Sandor, S.; Zhang, Y.; Xu, J. Fungal mitochondrial genomes and genetic polymorphisms. Appl. Microbiol. Biotechnol. 2018, 102, 9433–9448. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, J. Mitochondrial Genome Polymorphisms in the Human Pathogenic Fungus Cryptococcus neoformans. Front. Microbiol. 2020, 11, 706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Wang, X.-N.; Zhang, X.-L.; Liu, X.-Z.; Zhang, Y.-J. Complete mitochondrial genome of the endophytic fungus Pestalotiopsis fici: Features and evolution. Appl. Microbiol. Biotechnol. 2017, 101, 1593–1604. [Google Scholar] [CrossRef]
- Li, Q.; Yang, L.; Xiang, D.; Wan, Y.; Wu, Q.; Huang, W.; Zhao, G. The complete mitochondrial genomes of two model ectomycorrhizal fungi (Laccaria): Features, intron dynamics and phylogenetic implications. Int. J. Biol. Macromol. 2019, 145, 974–984. [Google Scholar] [CrossRef]
- Huang, W.; Feng, H.; Tu, W.; Xiong, C.; Jin, X.; Li, P.; Wang, X.; Li, Q. Comparative Mitogenomic Analysis Reveals Dynamics of Intron Within and Between Tricholoma Species and Phylogeny of Basidiomycota. Front. Genet. 2021, 12, 534871. [Google Scholar] [CrossRef]
- Chen, C.; Khaleel, S.S.; Huang, H.; Wu, C.H. Software for pre-processing Illumina next-generation sequencing short read sequences. Source Code Biol. Med. 2014, 9, 8. [Google Scholar] [CrossRef]
- Schubert, M.; Lindgreen, S.; Orlando, L. AdapterRemoval v2: Rapid adapter trimming, identification, and read merging. BMC Res. Notes 2016, 9, 88. [Google Scholar] [CrossRef] [Green Version]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: De novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 2017, 45, e18. [Google Scholar] [CrossRef] [Green Version]
- Valach, M.; Burger, G.; Gray, M.; Lang, B.F. Widespread occurrence of organelle genome-encoded 5S rRNAs including permuted molecules. Nucleic Acids Res. 2014, 42, 13764–13777. [Google Scholar] [CrossRef] [Green Version]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenet. Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef]
- Li, Q.; Ren, Y.; Xiang, D.; Shi, X.; Zhao, J.; Peng, L.; Zhao, G. Comparative mitogenome analysis of two ectomycorrhizal fungi (Paxillus) reveals gene rearrangement, intron dynamics, and phylogeny of basidiomycetes. IMA Fungus 2020, 11, 12. [Google Scholar] [CrossRef]
- Lohse, M.; Drechsel, O.; Bock, R. OrganellarGenomeDRAW (OGDRAW): A tool for the easy generation of high-quality custom graphical maps of plastid and mitochondrial genomes. Curr. Genet. 2007, 52, 267–274. [Google Scholar] [CrossRef]
- Wang, X.; Song, A.; Wang, F.; Chen, M.; Li, X.; Li, Q.; Liu, N. The 206 kbp mitochondrial genome of Phanerochaete carnosa reveals dynamics of introns, accumulation of repeat sequences and plasmid-derived genes. Int. J. Biol. Macromol. 2020, 162, 209–219. [Google Scholar] [CrossRef]
- Caspermeyer, J. MEGA Evolutionary Software Re-Engineered to Handle Today’s Big Data Demands. Mol. Biol. Evol. 2016, 33, 1887. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Stothard, P. The sequence manipulation suite: JavaScript programs for analyzing and formatting protein and DNA sequences. Biotechniques 2000, 28, 1102. [Google Scholar] [CrossRef] [Green Version]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Li, Q.; Fu, R.; Wang, J.; Deng, G.; Chen, X.; Lu, D. Comparative mitochondrial genome analysis reveals intron dynamics and gene rearrangements in two Trametes species. Sci. Rep. 2021, 11, 2569. [Google Scholar] [CrossRef]
- Martin, F.N.; Bensasson, D.; Tyler, B.M.; Boore, J.L. Mitochondrial genome sequences and comparative genomics of Phytophthora ramorum and P. sojae. Curr. Genet. 2007, 51, 285–296. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Luo, Q.; Ren, Y.; Luo, Z.; Liao, W.; Wang, X.; Li, Q. Panorama of intron dynamics and gene rearrangements in the phylum Basidiomycota as revealed by the complete mitochondrial genome of Turbinellus floccosus. Appl. Microbiol. Biotechnol. 2021, 105, 2017–2032. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Férandon, C.; Moukha, S.; Callac, P.; Benedetto, J.-P.; Castroviejo, M.; Barroso, G. The Agaricus bisporus cox1 Gene: The Longest Mitochondrial Gene and the Largest Reservoir of Mitochondrial Group I Introns. PLoS ONE 2010, 5, e14048. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, L.; Feng, H.; Tu, W.; Bao, Z.; Xiong, C.; Wang, X.; Qing, Y.; Huang, W. Characterization of the Complete Mitochondrial Genome of Basidiomycete Yeast Hannaella oryzae: Intron Evolution, Gene Rearrangement, and Its Phylogeny. Front. Microbiol. 2021, 12, 646567. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Briefings Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Vaidya, G.; Lohman, D.L.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New Methods for Selecting Partitioned Models of Evolution for Molecular and Morphological Phylogenetic Analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Yang, M.; Chen, C.; Xiong, C.; Jin, X.; Pu, Z.; Huang, W. Characterization and phylogenetic analysis of the complete mitochondrial genome of the medicinal fungus Laetiporus sulphureus. Sci. Rep. 2018, 8, 9104. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Costa, G.G.; Cabrera, O.G.; Tiburcio, R.A.; Medrano, F.; Carazzolle, M.F.; Thomazella, D.P.; Schuster, S.C.; Carlson, J.E.; Guiltinan, M.J.; Bailey, B.A.; et al. The mitochondrial genome of Moniliophthora roreri, the frosty pod rot pathogen of cacao. Fungal Biol. 2012, 116, 551–562. [Google Scholar] [CrossRef]
- Al-Reedy, R.M.; Malireddy, R.; Dillman, C.B.; Kennell, J.C. Comparative analysis of Fusarium mitochondrial genomes reveals a highly variable region that encodes an exceptionally large open reading frame. Fungal Genet. Biol. 2012, 49, 2–14. [Google Scholar] [CrossRef]
- Basse, C.W. Mitochondrial inheritance in fungi. Curr. Opin. Microbiol. 2010, 13, 712–719. [Google Scholar] [CrossRef]
- Deng, Y.; Hsiang, T.; Li, S.; Lin, L.; Wang, Q.; Chen, Q.; Xie, B.; Ming, R. Comparison of the Mitochondrial Genome Sequences of Six Annulohypoxylon stygium Isolates Suggests Short Fragment Insertions as a Potential Factor Leading to Larger Genomic Size. Front. Microbiol. 2018, 9, 2079. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.; Bao, Z.; Tu, W.; Li, L.; Xiong, C.; Jin, X.; Li, P.; Gui, M.; Huang, W.; Li, Q. The mitogenomes of two saprophytic Boletales species (Coniophora) reveals intron dynamics and accumulation of plasmid-derived and non-conserved genes. Comput. Struct. Biotechnol. J. 2020, 19, 401–414. [Google Scholar] [CrossRef]
- Adams, K.L.; Palmer, J.D. Evolution of mitochondrial gene content: Gene loss and transfer to the nucleus. Mol. Phylogenet. Evol. 2003, 29, 380–395. [Google Scholar] [CrossRef]
- Adams, K.L.; Qiu, Y.-L.; Stoutemyer, M.; Palmer, J.D. Punctuated evolution of mitochondrial gene content: High and variable rates of mitochondrial gene loss and transfer to the nucleus during angiosperm evolution. Proc. Natl. Acad. Sci. USA 2002, 99, 9905–9912. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Jia, L.; Wang, M.; Yang, H.; Chen, M.; Li, X.; Liu, H.; Li, Q.; Liu, N. The complete mitochondrial genome of medicinal fungus Taiwanofungus camphoratus reveals gene rearrangements and intron dynamics of Polyporales. Sci. Rep. 2020, 10, 16500. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Y.; Yao, W.; Shen, J.; Chen, M.; Gao, M.; Ren, J.; Li, Q.; Liu, N. The 256 kb mitochondrial genome of Clavaria fumosa is the largest among phylum Basidiomycota and is rich in introns and intronic ORFs. IMA Fungus 2020, 11, 26. [Google Scholar] [CrossRef]
- Li, Q.; He, X.; Ren, Y.; Xiong, C.; Jin, X.; Peng, L.; Huang, W. Comparative Mitogenome Analysis Reveals Mitochondrial Genome Differentiation in Ectomycorrhizal and Asymbiotic Amanita Species. Front. Microbiol. 2020, 11, 1382. [Google Scholar] [CrossRef]
- Turmel, M.; Côté, V.; Otis, C.; Mercier, J.P.; Gray, M.; Lonergan, K.M.; Lemieux, C. Evolutionary transfer of ORF-containing group I introns between different subcellular compartments (chloroplast and mitochondrion). Mol. Biol. Evol. 1995, 12, 533–545. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Chen, C.; Xiong, C.; Jin, X.; Chen, Z.; Huang, W. Comparative mitogenomics reveals large-scale gene rearrangements in the mitochondrial genome of two Pleurotus species. Appl. Microbiol. Biotechnol. 2018, 102, 6143–6153. [Google Scholar] [CrossRef]
- Lavrov, D.V.; Boore, J.L.; Brown, W.M. Complete mtDNA Sequences of Two Millipedes Suggest a New Model for Mitochondrial Gene Rearrangements: Duplication and Nonrandom Loss. Mol. Biol. Evol. 2002, 19, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Zhong, L.; Wang, M.; Li, D.; Tang, S.; Zhang, T.; Bian, W.; Chen, X. Complete mitochondrial genome of Odontobutis haifengensis (Perciformes, Odontobutiae): A unique rearrangement of tRNAs and additional non-coding regions identified in the genus Odontobutis. Genomics 2018, 110, 382–388. [Google Scholar] [CrossRef]
- Aguileta, G.; de Vienne, D.M.; Ross, O.N.; Hood, M.E.; Giraud, T.; Petit, E.; Gabaldón, T. High Variability of Mitochondrial Gene Order among Fungi. Genome Biol. Evol. 2014, 6, 451–465. [Google Scholar] [CrossRef] [Green Version]
- Hseu, R.S.; Wang, H.H.; Wang, H.F.; Moncalvo, J.M. Differentiation and grouping of isolates of the Ganoderma lucidum com-plex by random amplified polymorphic DNA-PCR compared with grouping on the basis of internal transcribed spacer se-quences. Appl. Environ. Microbiol. 1996, 62, 1354–1363. [Google Scholar] [CrossRef] [Green Version]
- Kuok, Q.-Y.; Yeh, C.-Y.; Su, B.-C.; Hsu, P.-L.; Ni, H.; Liu, M.-Y.; Mo, F.-E. The triterpenoids of Ganoderma tsugae prevent stress-induced myocardial injury in mice. Mol. Nutr. Food Res. 2013, 57, 1892–1896. [Google Scholar] [CrossRef]
- Hsu, W.-H.; Qiu, W.-L.; Tsao, S.-M.; Tseng, A.-J.; Lu, M.-K.; Hua, W.-J.; Cheng, H.-C.; Hsu, H.-Y.; Lin, T.-Y. Effects of WSG, a polysaccharide from Ganoderma lucidum, on suppressing cell growth and mobility of lung cancer. Int. J. Biol. Macromol. 2020, 165, 1604–1613. [Google Scholar] [CrossRef]
- Tel-Çayan, G.; Muhammad, A.; Deveci, E.; Duru, M.E.; Öztürk, M. Isolation, structural characterization, and biological activities of galactomannans from Rhizopogon luteolus and Ganoderma adspersum mushrooms. Int. J. Biol. Macromol. 2020, 165, 2395–2403. [Google Scholar] [CrossRef]
- Jiang, L.; Zhao, L.; Cheng, D.; Zhu, L.; Zhang, M.; Ruan, Q.; Chen, W. The complete mitochondrial genome sequence of the Sichuan Digging Frog, Kaloula rugifera (Anura: Microhylidae) and its phylogenetic implications. Gene 2017, 626, 367–375. [Google Scholar] [CrossRef]
- Li, W.; Wang, Z.; Che, Y. The Complete Mitogenome of the Wood-Feeding Cockroach Cryptocercus meridianus (Blattodea: Cryptocercidae) and Its Phylogenetic Relationship among Cockroach Families. Int. J. Mol. Sci. 2017, 18, 12397. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Bao, Z.; Tang, K.; Feng, H.; Tu, W.; Li, L.; Han, Y.; Cao, M.; Zhao, C. First two mitochondrial genomes for the order Filobasidiales reveal novel gene rearrangements and intron dynamics of Tremellomycetes. IMA Fungus 2022, 13, 7. [Google Scholar] [CrossRef] [PubMed]
- Binder, M.; Justo, A.; Riley, R.; Salamov, A.; Lopez-Giraldez, F.; Sjökvist, E.; Copeland, A.; Foster, B.; Sun, H.; Larsson, E.; et al. Phylogenetic and phylogenomic overview of the Polyporales. Mycologia 2013, 105, 1350–1373. [Google Scholar] [CrossRef] [Green Version]
- Justo, A.; Miettinen, O.; Floudas, D.; Ortiz-Santana, B.; Sjökvist, E.; Lindner, D.; Nakasone, K.; Niemelä, T.; Larsson, K.-H.; Ryvarden, L.; et al. A revised family-level classification of the Polyporales (Basidiomycota). Fungal Biol. 2017, 121, 798–824. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wu, S.; Ma, X.; Chen, W.; Zhang, J.; Duan, S.; Gao, Y.; Kui, L.; Huang, W.; Wu, P.; et al. The Genome Sequences of 90 Mushrooms. Sci. Rep. 2018, 8, 9982. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PCGs | G. lucidum | G. lingzhi | Function | ||||
|---|---|---|---|---|---|---|---|
| GL | s26 | s37 | s62 | s74 | s8 | ||
| orf109 | * | * | * | * | * | * | RNA polymerase |
| orf115 | / | / | / | * | / | / | DNA polymerase |
| orf118 | / | / | / | * | * | / | GIY-YIG endonuclease |
| orf151 | * | * | *(orf167) | * | * | hypothetical protein | |
| orf152 | / | / | / | / | * | / | hypothetical protein |
| orf190 | / | / | * | / | * | / | DNA polymerase |
| orf192 | * | * | / | / | / | * | hypothetical protein |
| orf202 | / | / | / | * | / | / | RNA polymerase |
| orf211 | / | / | * | / | / | RNA polymerase | |
| orf219 | / | / | * | / | * | / | LAGLIDADG endonuclease |
| orf250 | / | / | / | * | / | / | DNA polymerase |
| orf283 | * | / | * | / | / | / | DNA polymerase |
| orf294 | * | * | / | / | / | * | hypothetical protein |
| orf309 | / | / | / | * | / | / | LAGLIDADG endonuclease |
| orf396 | * | * | * | / | * | * | DNA polymerase |
| orf413 | / | / | / | * | / | hypothetical protein | |
| orf429 | *(orf439) | * | * | / | * | * | hypothetical protein |
| orf568 | * | * | * | / | * | * | RNA polymerase |
| orf744 | / | / | / | / | * | / | DNA polymerase |
| orf879 | / | / | / | / | * | / | DNA polymerase |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Zhang, T.; Li, L.; Bao, Z.; Tu, W.; Xiang, P.; Wu, Q.; Li, P.; Cao, M.; Huang, W. Comparative Mitogenomic Analysis Reveals Intraspecific, Interspecific Variations and Genetic Diversity of Medical Fungus Ganoderma. J. Fungi 2022, 8, 781. https://doi.org/10.3390/jof8080781
Li Q, Zhang T, Li L, Bao Z, Tu W, Xiang P, Wu Q, Li P, Cao M, Huang W. Comparative Mitogenomic Analysis Reveals Intraspecific, Interspecific Variations and Genetic Diversity of Medical Fungus Ganoderma. Journal of Fungi. 2022; 8(8):781. https://doi.org/10.3390/jof8080781
Chicago/Turabian StyleLi, Qiang, Ting Zhang, Lijiao Li, Zhijie Bao, Wenying Tu, Peng Xiang, Qian Wu, Ping Li, Mei Cao, and Wenli Huang. 2022. "Comparative Mitogenomic Analysis Reveals Intraspecific, Interspecific Variations and Genetic Diversity of Medical Fungus Ganoderma" Journal of Fungi 8, no. 8: 781. https://doi.org/10.3390/jof8080781