
Taxonomy and Phylogeny of Meruliaceae with Descriptions of Two New Species from China



Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological Studies
2.2. DNA Extraction, PCR, and Sequencing
2.3. Phylogenetic Analyses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sample | GenBank Accession No. | References | |
|---|---|---|---|---|
| ITS | nLSU | |||
| Aurantiopileus mayaensi | JV 1504/128 | KT156706 | — | — |
| A. mayaensi | TJB10228 | HM772140 | HM772139 | [18] |
| Aurantiporus croceus | Miettinen-16483 | KY948745 | KY948901 | [2] |
| A. roseus | Dai 13573 | KJ698635 | KJ698639 | [19] |
| Ceriporiopsis alboaurantia | Cui 4136 | KF845955 | KF845948 | [20] |
| C. alboaurantia | Cui 2877 | KF845954 | KF845947 | [20] |
| C. fimbriata | Cui 1671 | KJ698634 | KJ698638 | [19] |
| C. fimbriata | Dai 11672 | KJ698633 | KJ698637 | [19] |
| C. gilvescens | BRNM 710166 | FJ496684 | FJ496684 | [21] |
| C. gilvescens | BRNM 667882 | FJ496685 | FJ496719 | [21] |
| C. guidella | HUBO 7659 | FJ496687 | FJ496722 | [21] |
| C. kunmingensis | CLZhao 152 | KX081072 | KX081074 | [22] |
| C. kunmingensis | CLZhao 153 | KX081073 | KX081075 | [22] |
| C. lagerheimii | 58240 | KX008365 | KX081077 | [23] |
| C. pseudoplacenta | PRM 899297 | JN592497 | JN592504 | [24] |
| C. pseudoplacenta | PRM 899300 | JN592498 | JN592505 | [24] |
| C. semisupina | Cui 10222 | KF845956 | KF845949 | [20] |
| C. semisupina | Cui 7971 | KF845957 | KF845950 | [20] |
| Climacodon septentrionalis | AFTOL-767 | AY854082 | AY684165 | [25] |
| C. septentrionalis | RLG-6890-Sp | KP135344 | — | [26] |
| Crustodontia chrysocreas | HHB-3946 | KP135357 | — | [26] |
| C. chrysocreas | HHB-6333-Sp | KP135358 | KP135263 | [26] |
| C. nigrodontea | CLZhao 2758 | MT896824 | — | [5] |
| C. nigrodontea | CLZhao 2445 | MT896821 | MT896818 | [27] |
| C. sp. | KUC20121123-24 | KJ668482 | — | [28] |
| C. tongxiniana | CLZhao 2255 | MT020773 | MT020751 | [27] |
| C. tongxiniana | CLZhao 2316 | MT020774 | MT020752 | [27] |
| Geesterania carneola | MCW 388/12 | KY174999 | KY174999 | [29] |
| G. davidii | MCW 396/12 | KY174998 | KY174998 | [29] |
| Hermanssonia centrifuga | CBS 125890 | MH864088 | MH875547 | [30] |
| H. centrifuga | HHB-9239-Sp | KP135380 | KP135262 | [26] |
| H. fimbriata | Dai 23266 | ON135436 | ON135440 | Present study |
| H. fimbriata | Dai 23305 | ON135437 | ON135441 | Present study |
| H. fimbriata | Dai 23306 | ON135438 | ON135442 | Present study |
| Hydnophanerochaete odontoidea | CLZhao 3882 | MH784919 | MH784929 | [31] |
| H. odontoidea | CLZhao 4036 | MH784927 | MH784937 | [31] |
| Hydnophlebia chrysorhiza | FD-282 | KP135338 | KP135217 | [26] |
| H. chrysorhiza | HHB-18767 | KP135337 | — | [26] |
| Hyphoderma mutatum | HHB-15479-Sp | KP135296 | KP135221 | [26] |
| H. setigerum | FD-312 | KP135297 | KP135222 | [26] |
| Lilaceophlebia livida | FCUG 2189 | AF141624 | AF141624 | [21] |
| L. livida | FCUG 1290 | HQ153414 | — | [32] |
| L. subserialis | FCUG 1434 | AF141631 | AF141631 | — |
| Luteochaete subglobosa | CLZhao 3639 | MK881898 | MK881788 | [33] |
| L. subglobosa | CLZhao 3475 | MK881897 | MK881787 | [33] |
| Luteoporia albomarginata | Dai 15229 | KU598873 | KU598878 | [34] |
| L. albomarginata | GC 1702-1 | LC379003 | LC379155 | [35] |
| L. citriniporia | Dai 19507 | MT872218 | MT872216 | [36] |
| L. citriniporia | Dai 19622 | MT872219 | MT872217 | [36] |
| L. lutea | CHWC 1506-68 | MZ636997 | MZ637157 | [3] |
| L. lutea | GC 1409-1 | MZ636998 | MZ637158 | [3] |
| Mycoacia aurea | DLL 2011263 | KJ140747 | — | [1] |
| M. aurea | RLG-5075-Sp | KY948759 | MZ637161 | [2,3] |
| M. aurea | DLL2011_100 | KJ140614 | — | [37] |
| M. fuscoatra | HHB 15354T | KP135367 | — | [26] |
| M. cf. kurilensis | WEI 18-312 | MZ637001 | MZ637162 | [3] |
| M. cf. kurilensis | WEI 18-324 | MZ637002 | MZ637163 | [3] |
| M. fuscoatra | KHL 13275 | JN649352 | JN649352 | [21] |
| M. nothofagi | HHB 12067 | KP135370 | — | [26] |
| M. nothofagi | KHL 13750 | GU480000 | GU480000 | [21] |
| Mycoaciella bispora | EL13_99 | — | AY586692 | [38] |
| M. efibulata | WEI 19-057 | MZ637012 | MZ637172 | [3] |
| M. efibulata | WEI 16-172 | MZ637011 | MZ637171 | [3] |
| Odoria alborubescens | BP106943 | MG097864 | MG097867 | [39] |
| O. alborubescens | BRNU 627479 | JQ821319 | JQ821318 | [40] |
| Pappia fissilis | 814 | HQ728291 | HQ729001 | [41] |
| P. fissilis | BRNM 699803 | HQ728292 | HQ729002 | [41] |
| Phlebia acanthocystis | KUC20131001-33 | KJ668484 | KJ668337 | [26] |
| P. acanthocystis | FP150571 | KY948767 | KY948844 | [2] |
| P. acerina | FD 301 | KP135378 | — | [2] |
| P. acerina | HHB 11146 | KP135372 | — | [26] |
| P. austroasiana | Dai 17556 | ON135439 | ON135443 | Present study |
| P. austroasiana | E8898A | KJ654590 | — | [42] |
| P. brevispora | HHB 7030 | KP135387 | — | [26] |
| P. brevispora | FBCC1463 | LN611135 | LN611135 | [43] |
| P. floridensis | HHB 7175 | KP135384 | — | [26] |
| P. floridensis | HHB-9905-Sp | KP135383 | KP135264 | [26] |
| P. fuscotuberculata | CLZhao 10227 | MT020759 | MT020737 | [27] |
| P. fuscotuberculata | CLZhao 10239 | MT020760 | MT020738 | [27] |
| P. hydnoidea | HHB-1993-Sp | KY948778 | KY948853 | [2] |
| P. lindtneri | GB-1027 | AB210076 | — | [44] |
| P. lindtneri | GB-501 | KY948772 | KY948847 | [2] |
| P. ludoviciana | HHB-8715-Sp | KY948770 | KY948846 | [2] |
| P. ludoviciana | FD-427 | KP135342 | — | [26] |
| P. nantahaliensis | HHB-2816-Sp | KY948777 | KY948852 | [2] |
| P. radiata | CBS 285.56 | MH857642 | MH869187 | [30] |
| P. radiata | AFTOL-484 | AY854087 | AF287885 | [25] |
| P. radiata | UBC: F19726 | HQ604797 | HQ604797 | [1] |
| P. rufa | FBCC297 | LN611092 | LN611092 | [43] |
| P. rufa | HHB-14924 | KP135374 | — | [26] |
| P. serialis | FCUG 2868 | HQ153429 | — | [32] |
| P. serialis | UC2023146 | KP814195 | — | [33] |
| P. setulosa | PH 11749 | GU461312 | — | [1] |
| P. setulosa | HHB-6891-Sp | KP135382 | KP135267 | [26] |
| P. setulosa | AH31879 | GQ259417 | GQ259417 | [45] |
| P. subochracea I | KGN 162/95 | EU118656 | EU118656 | [46] |
| P. subochracea II | FBCC295 | LN611116 | LN611116 | [43] |
| P. subochracea II | HHB-8494-Sp | KY948768 | KY948845 | [2] |
| P. tomentopileata | CLZhao 9563 | MT020765 | MT020743 | [27] |
| P. tomentopileata | CLZhao 9515 | MT020764 | MT020742 | [27] |
| P. tremellosa | ES 20082 | JX109859 | JX109859 | [1] |
| P. tremellosa | CBS 217.56 | MH857589 | MH869138 | [30] |
| Phlebiporia bubalina | Dai 13168 | KC782526 | KC782528 | [47] |
| P. bubalina | Dai 15179 | KY131843 | KY131902 | [48] |
| Sarcodontia uda | FP-101544-Sp | KP135361 | KP135232 | [26] |
| Sarcodontia uda | USDA Kropp 1 | KY948764 | — | [2] |
| Scopuloides hydnoides | FP-150473 | KP135355 | KP135284 | [26] |
| S. hydnoides | WEI 17-569 | MZ637085 | MZ637283 | [3] |
| Stereophlebia tuberculata | FCUG 3157 | HQ153427 | — | [32] |
| S. tuberculata | Wu 1708-107 | MZ637089 | MZ637286 | [3] |
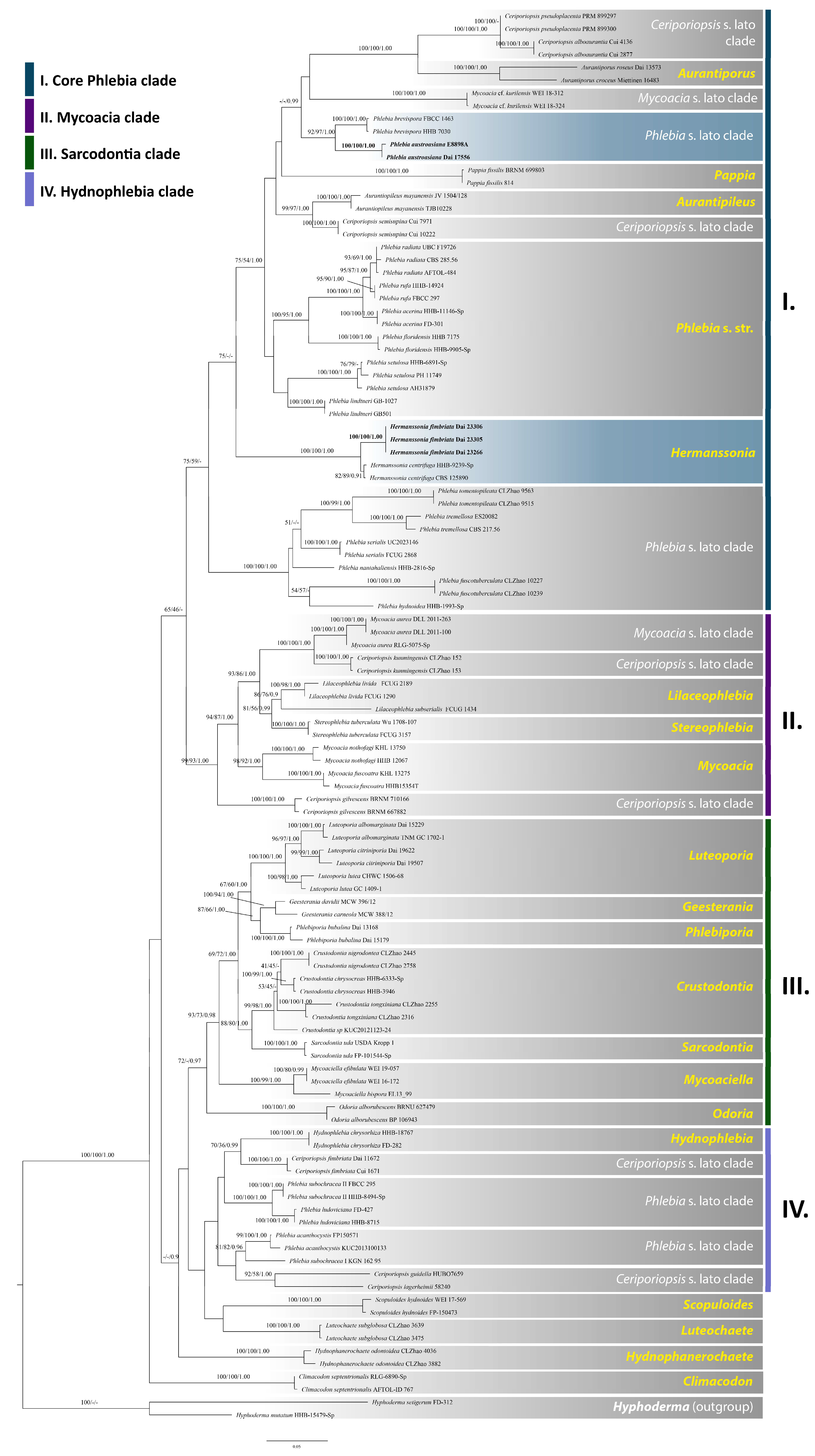
3. Results
3.1. Phylogenetic Analyses
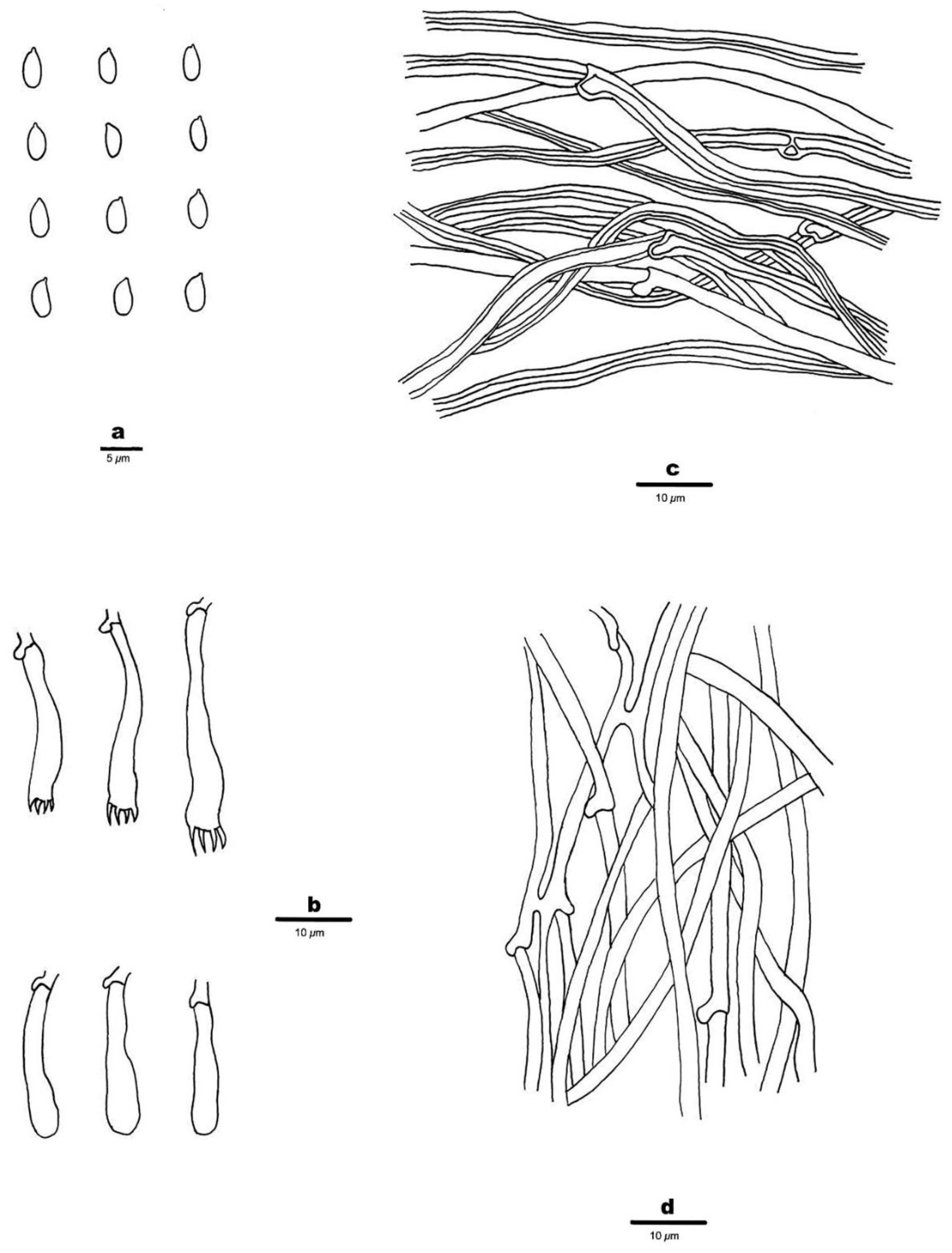
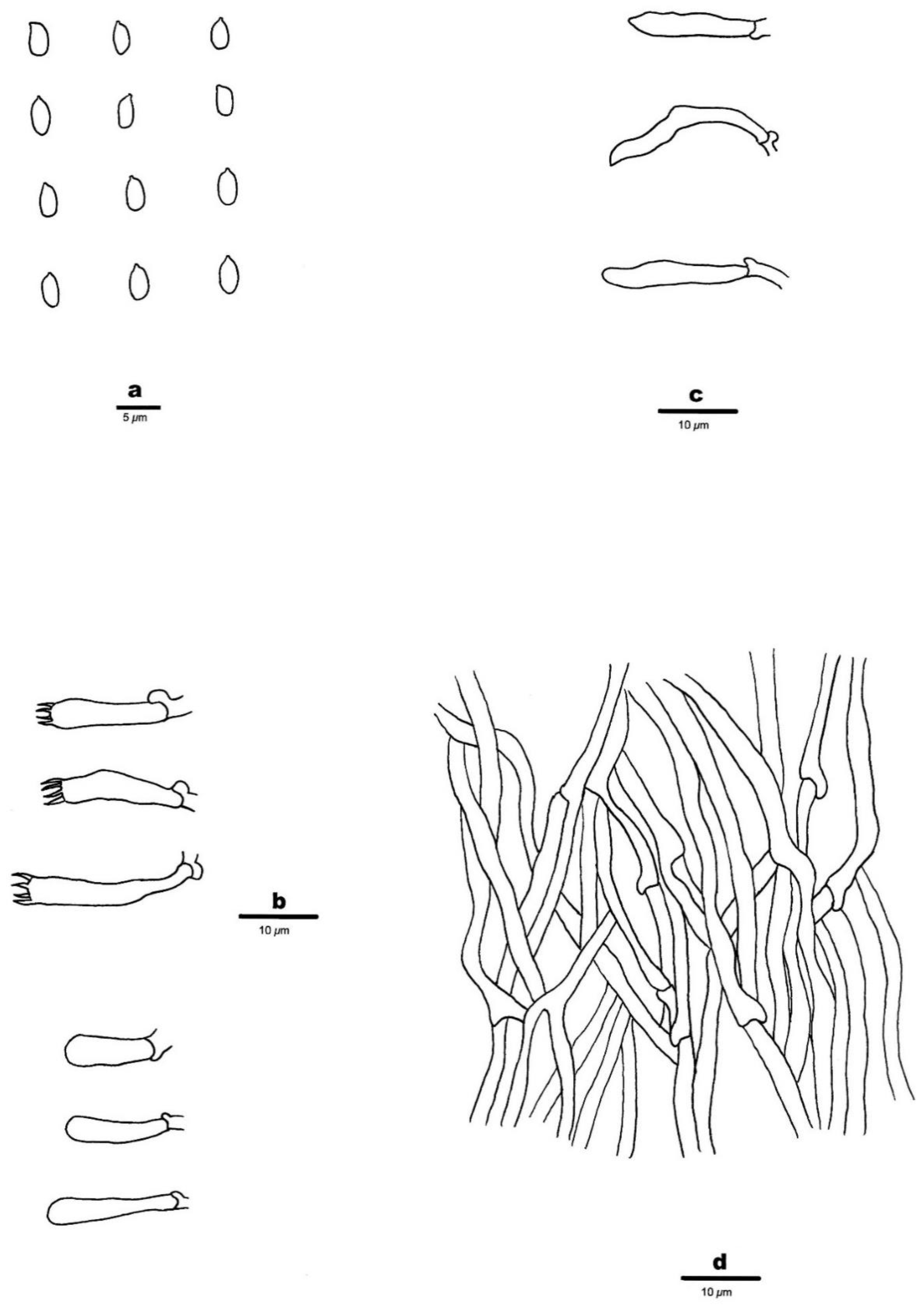
3.2. Taxonomy
- 1.
- 2.
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Binder, M.; Hibbett, D.S.; Larsson, K.H.; Larsson, E.; Langer, E.; Langer, G. The phylogenetic distribution of resupinate forms across the major clades of mushroom-forming fungi (Homobasidiomycetes). Syst. Biodivers. 2005, 3, 113–157. [Google Scholar] [CrossRef]
- Justo, A.; Miettinen, O.; Floudas, D.; Ortiz-Santana, B.; Sjökvist, E.; Lindner, D.; Nakasone, K.K.; Niemelä, T.; Larsson, K.H.; Ryvarden, L. A revised family-level classification of the Polyporales (Basidiomycota). Fungal Biol. 2017, 121, 798–824. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Chen, C.Y.; Wu, S.H. Species diversity, taxonomy and multi-gene phylogeny of phlebioid clade (Phanerochaetaceae, Irpicaceae, Meruliaceae) of Polyporales. Fungal Divers. 2021, 111, 337–442. [Google Scholar] [CrossRef]
- Fries, E.M. Epicrisis systematis mycologici, seu Synopsis Hymenomycetum. xiv, 610. Syst. Mycol. 1821, 1, 1838. [Google Scholar]
- Huang, R.X.; Luo, K.Y.; Zhao, C.L. Phlebia nigrodontea sp. nov. in Meruliaceae (Polyporales) with a black hymenial surface. Phytotaxa 2020, 458, 195–206. [Google Scholar] [CrossRef]
- de Sousa Lira, C.R.; Chikowski, R.d.S.; de Lima, V.X.; Gibertoni, T.B.; Larsson, K.H. Allophlebia, a new genus to accomodate Phlebia ludoviciana (Agaricomycetes, Polyporales). Mycol. Prog. 2022, 21, 47. [Google Scholar] [CrossRef]
- Zmitrovich, I.V. Conspectus systematis Polyporacearum v. 1.0. Folia Cryptogam. Petropolitana 2018, 6, 3–145. [Google Scholar] [CrossRef]
- Wu, F.; Zhou, L.W.; Vlasák, J.; Dai, Y.C. Global diversity and systematics of Hymenochaetaceae with poroid hymenophore. Fungal Divers. 2022, 113, 1–192. [Google Scholar] [CrossRef]
- Petersen, J.H. Farvekort. The Danish Mycological Society’s Colour-Chart; Foreningen til Svampekundskabens Fremme: Greve, Italy, 1996; pp. 1–6. [Google Scholar]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff; New York Botanical Garden’s Virtual Herbarium: New York, NY, USA, 2018; Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 15 January 2021).
- Li, H.J.; Cui, B.K.; Dai, Y.C. Taxonomy and multi-gene phylogeny of Datronia (Polyporales, Basidiomycota). Persoonia 2014, 32, 170–182. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gefand, D.H., Sninsky, J.J., White, J.T., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.L.; Cui, B.K.; Song, J.; Dai, Y.C. Fragiliporiaceae, a new family of Polyporales (Basidiomycota). Fungal Divers. 2015, 70, 115–126. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. Bioedit: A user–friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.70. Available online: https://www.mesquiteproject.org/ (accessed on 18 April 2022).
- Ginns, J.; Lindner, D.L.; Baronia, T.J.; Ryvarden, L. Aurantiopileus mayanensis a new genus and species of polypore (Polyporales, Basidiomycota) from Belize with connections to existing Asian species. N. Am. Fungi 2010, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.L.; Wu, F.; Liu, H. A phylogenetic and taxonomic study on Ceriporiopsis s. str. (Polyporales) in China. Nova Hedwig. 2015, 101, 403–417. [Google Scholar] [CrossRef]
- Zhao, C.L.; Cui, B.K. Phylogeny and taxonomy of Ceriporiopsis (Polyporales) with descriptions of two new species from southern China. Phytotaxa 2014, 164, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Tomšovský, M.; Menkis, A.; Vasaitis, R. Phylogenetic relationships in European Ceriporiopsis species inferred from nuclear and mitochondrial ribosomal DNA sequences. Fungal Biol. 2010, 114, 350–358. [Google Scholar] [CrossRef]
- Zhao, C.L.; Wu, Z.Q. Ceriporiopsis kunmingensis sp. nov. (Polyporales, Basidiomycota) evidenced by morphological characters and phylogenetic analysis. Mycol. Prog. 2017, 16, 93–100. [Google Scholar] [CrossRef]
- Zhao, C.L.; Ren, G.J.; Wu, F. A new species of Hyphodermella (Polyporales, Basidiomycota) with a poroid hymenophore. Mycoscience 2017, 58, 452–456. [Google Scholar] [CrossRef]
- Vlasák, J.; Vlasák, J., Jr.; Ryvarden, L. Four new polypore species from the western United States. Mycotaxon 2012, 119, 217–231. [Google Scholar] [CrossRef]
- Lutzoni, F.; Kauff, F.; Cox, C.J.; McLaughlin, D.; Celio, G.; Dentinger, B.; Padamsee, M.; Hibbett, D.; James, T.Y.; Baloch, E.; et al. Assembling the fungal tree of life: Progress, classification, and evolution of subcellular traits. Am. J. Bot. 2004, 91, 1446–1480. [Google Scholar] [CrossRef]
- Floudas, D.; Hibbett, D.S. Revisiting the taxonomy of Phanerochaete (Polyporales, Basidiomycota) using a four gene dataset and extensive ITS sampling. Fungal Biol. 2015, 119, 679–719. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.X.; Zhao, C.L. Three new species of Phlebia (Polyporales, Basidiomycota) based on the evidence from morphology and DNA sequence data. Mycol. Prog. 2020, 19, 753–767. [Google Scholar] [CrossRef]
- Jang, Y.; Jang, S.; Lee, J.; Lee, H.; Lim, Y.W.; Kim, C.; Kim, J.J. Diversity of wood-inhabiting polyporoid and corticioid fungi in Odaesan National Park, Korea. Mycobiology 2016, 44, 217–236. [Google Scholar] [CrossRef] [Green Version]
- Westphalen, M.C.; Rajchenberg, M.; Tomšovský, M.; Gugliotta, A.M. A re-evaluation of Neotropical Junghuhnia s. lat. (Polyporales, Basidiomycota) based on morphological and multigene analyses. Pers. Mol. Phylogeny Evol. Fungi 2018, 41, 130. [Google Scholar] [CrossRef] [PubMed]
- Vu, D.; Groenewald, M.; De Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Verkley, G.J.M. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Ma, X.; Xu, T.M.; Zhao, C.L. Phlebia ailaoshanensis sp. nov. (Polyporales, Basidiomycota) evidenced by morphological characters and phylogenetic analyses. Phytotaxa 2018, 373, 184–196. [Google Scholar] [CrossRef]
- Ghobad-Nejhad, M.; Hallenberg, N. Multiple evidence for recognition of Phlebia tuberculata, a more widespread segregate of Phlebia livida (Polyporales, Basidiomycota). Mycol. Prog. 2012, 11, 27–35. [Google Scholar] [CrossRef]
- Huang, R.X.; Luo, K.Y.; Ma, R.X.; Zhao, C.L. Morphological and molecular identification of Phlebia wuliangshanensis sp. nov. in China. Mycotaxon 2020, 135, 103–117. [Google Scholar] [CrossRef]
- Wu, F.; Yuan, Y.; Chen, J.J.; He, S.H. Luteoporia albomarginata gen. et sp. nov. (Meruliaceae, Basidiomycota) from tropical China. Phytotaxa 2016, 263, 31–41. [Google Scholar] [CrossRef]
- Chen, C.C.; Wu, S.H.; Chen, C.Y. Hydnophanerochaete and two new genera of phanerochaetoid fungi (Polyporales, Basidiomycota) from East Asia. MycoKeys 2018, 39, 75–96. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.B.; Yuan, Y. Luteoporia citriniporia sp. nov. (Polyporales, Basidiomycota), evidenced by morphological characters and phylogenetic analysis. Phytotaxa 2020, 461, 31–39. [Google Scholar] [CrossRef]
- Brazee, N.J.; Lindner, D.L.; D’Amato, A.W.; Fraver, S.; Forrester, J.A.; Mladeno, D.J. Disturbance and diversity of wood-inhabiting fungi: Effects of canopy gaps and downed woody debris. Biodivers. Conserv. 2014, 23, 2155–2172. [Google Scholar] [CrossRef]
- Larsson, K.H.; Larsson, E.; Kõljalg, U. High phylogenetic diversity among corticioid homobasidiomycetes. Mycol. Res. 2004, 108, 983–1002. [Google Scholar] [CrossRef]
- Papp, V.; Dima, B. New systematic position of Aurantiporus alborubescens (Meruliaceae, Basidiomycota), a threatened old-growth forest polypore. Mycol. Prog. 2017, 17, 319–332. [Google Scholar] [CrossRef]
- Dvořák, D.; Běťák, J.; Tomšovský, M. Aurantiporus alborubescens (Basidiomycota, Polyporales)–first record in the Carpathians and notes on its systematic position. Czech Mycol. 2014, 66, 71–84. [Google Scholar] [CrossRef] [Green Version]
- Tomšovský, M. Delimitation of an almost forgotten species Spongipellis litschaueri (Polyporales, Basidiomycota) and its taxonomic position within the genus. Mycol. Prog. 2012, 11, 415–424. [Google Scholar] [CrossRef]
- Glen, M.; Yuskianti, V.; Puspitasari, D.; Francis, A.; Agustini, L.; Rimbawanto, A.; Indrayadi, H.; Gafur, A.; Mohammed, C.L. Identification of basidiomycete fungi in Indonesian hardwood plantations by DNA barcoding. For. Pathol. 2014, 44, 496–508. [Google Scholar] [CrossRef]
- Kuuskeri, J.; Mäkelä, M.R.; Isotalo, J.; Oksanen, I.; Lundell, T. Lignocellulose-converting enzyme activity profiles correlate with molecular systematics and phylogeny grouping in the incoherent genus Phlebia (Polyporales, Basidiomycota). BMC Microbiol. 2015, 15, 217. [Google Scholar] [CrossRef] [Green Version]
- Kamei, I.; Suhara, H.; Kondo, R. Phylogenetical approach to isolation of white-rot fungi capable of degrading polychlorinated dibenzo-p-dioxin. Appl. Microbiol. Biotechnol. 2005, 69, 358–366. [Google Scholar] [CrossRef]
- Moreno, G.; Blanco, M.N.; Checa, J.; Platas, G.; Peláez, F. Taxonomic and phylogenetic revision of three rare irpicoid species within the Meruliaceae. Mycol. Prog. 2011, 10, 481–491. [Google Scholar] [CrossRef]
- Larsson, K.H. Re-thinking the classification of corticioid fungi. Mycol. Res. 2007, 111, 1040–1063. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.J.; Cui, B.K. Phlebiporia bubalina gen. et. sp. nov. (Meruliaceae, Polyporales) from Southwest China with a preliminary phylogeny based on rDNA sequences. Mycol. Prog. 2014, 13, 563–573. [Google Scholar] [CrossRef]
- Wu, F.; Chen, J.J.; Ji, X.H.; Vlasák, J.; Dai, Y.C. Phylogeny and diversity of the morphologically similar polypore genera Rigidoporus, Physisporinus, Oxyporus, and Leucophellinus. Mycologia 2017, 109, 749–765. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. PAUP: Phylogenetic Analysis Using Parsimony Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Felsenstein, J. Confidence intervals on phylogenetics: An approach using bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML Version 8: A tool for phylogenetic analyses and post analyses of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.A.; Holder, M.T.; Vos, R.; Midford, P.E.; Liebowitz, T.; Chan, L.; Hoover, P.; Warnow, T. The CIPRES Portals. CIPRES. Available online: http://www.phylo.org/sub_sections/portal (accessed on 4 August 2009).
- Ronquist, F.; Teslenko, M.; Mark, P.; Avres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes3.2: Efficient Bayesian phylogenetic inference and model choice, across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre: Uppsala, Sweden, 2004. [Google Scholar]
- Rambaut, A. Molecular Evolution, Phylogenetics and Epidemiology. FigTree ver. 1.4.4 Software. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 18 April 2022).
- Salo, P.; Niemela, T.; Salo, U. Suomen Sieniopas; Kasvimuseo/WSOY: Helsinik, Finland, 2005; pp. 1–512. [Google Scholar]
- Karsten, P.A. Symbolae ad mycologiam Fennicam 8. Medd. Af Soc. Pro Fauna Et Flora Fenn. 1881, 6, 7–13. [Google Scholar]
- Pilát, A. Additamenta ad floram Sibiriae Asiaeque orientalis mycologicam. Pars Secunda. Bull. Trimest. Société Mycol. Fr. 1933, 49, 256–339. [Google Scholar]
- Cooke, W.B. The genus Phlebia. Mycologia 1956, 48, 386–405. [Google Scholar] [CrossRef]
- Bernicchia, A.; Gorjón, S.P. Corticiaceae s.l. Fungi Europaei 12; Candusso Edizioni: Origgio, Italy, 2010; 1007p. [Google Scholar]
- Nakasone, K.K.; Eslyn, W.E. A new species, Phlebia brevispora, a cause of internal decay in utility poles. Mycologia 1981, 73, 803–810. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.-B.; Zhang, J.-L.; Papp, V.; Dai, Y.-C. Taxonomy and Phylogeny of Meruliaceae with Descriptions of Two New Species from China. J. Fungi 2022, 8, 501. https://doi.org/10.3390/jof8050501
Liu Z-B, Zhang J-L, Papp V, Dai Y-C. Taxonomy and Phylogeny of Meruliaceae with Descriptions of Two New Species from China. Journal of Fungi. 2022; 8(5):501. https://doi.org/10.3390/jof8050501
Chicago/Turabian StyleLiu, Zhan-Bo, Jun-Li Zhang, Viktor Papp, and Yu-Cheng Dai. 2022. "Taxonomy and Phylogeny of Meruliaceae with Descriptions of Two New Species from China" Journal of Fungi 8, no. 5: 501. https://doi.org/10.3390/jof8050501