
Diversity among Lasiodiplodia Species Causing Dieback, Root Rot and Leaf Spot on Fruit Trees in Egypt, and a Description of Lasiodiplodia newvalleyensis sp. nov.

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Isolation
2.2. DNA Extraction and PCR Amplification
2.3. Phylogenetic Analyses
2.4. Morphological Examination
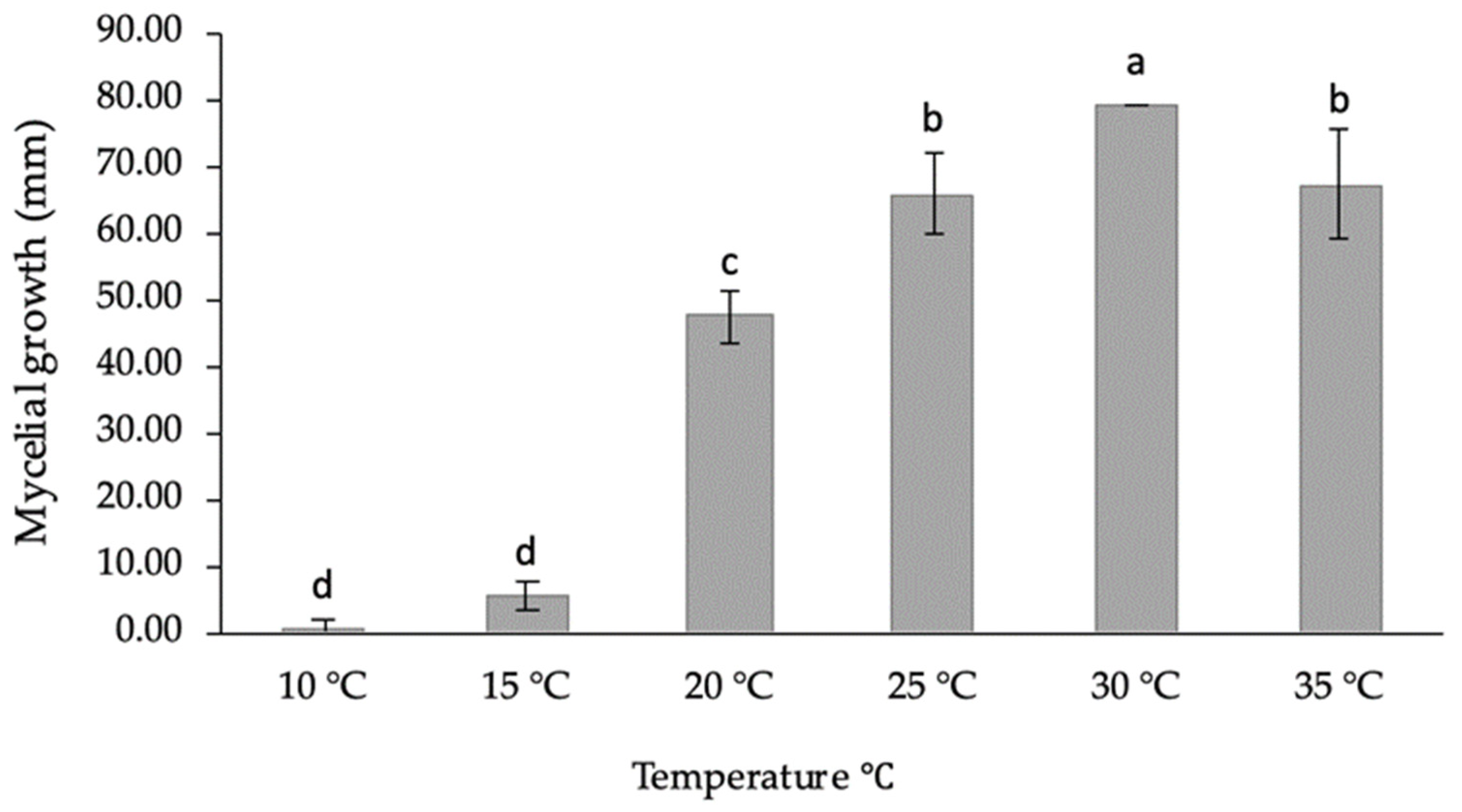
2.5. Evaluation of Temperature’s Effect on the Mycelial Growth
2.6. Pathogenicity Test on Seedlings and Leaves
2.7. Data Analysis
3. Results
3.1. Symptoms, Isolation and Frequency
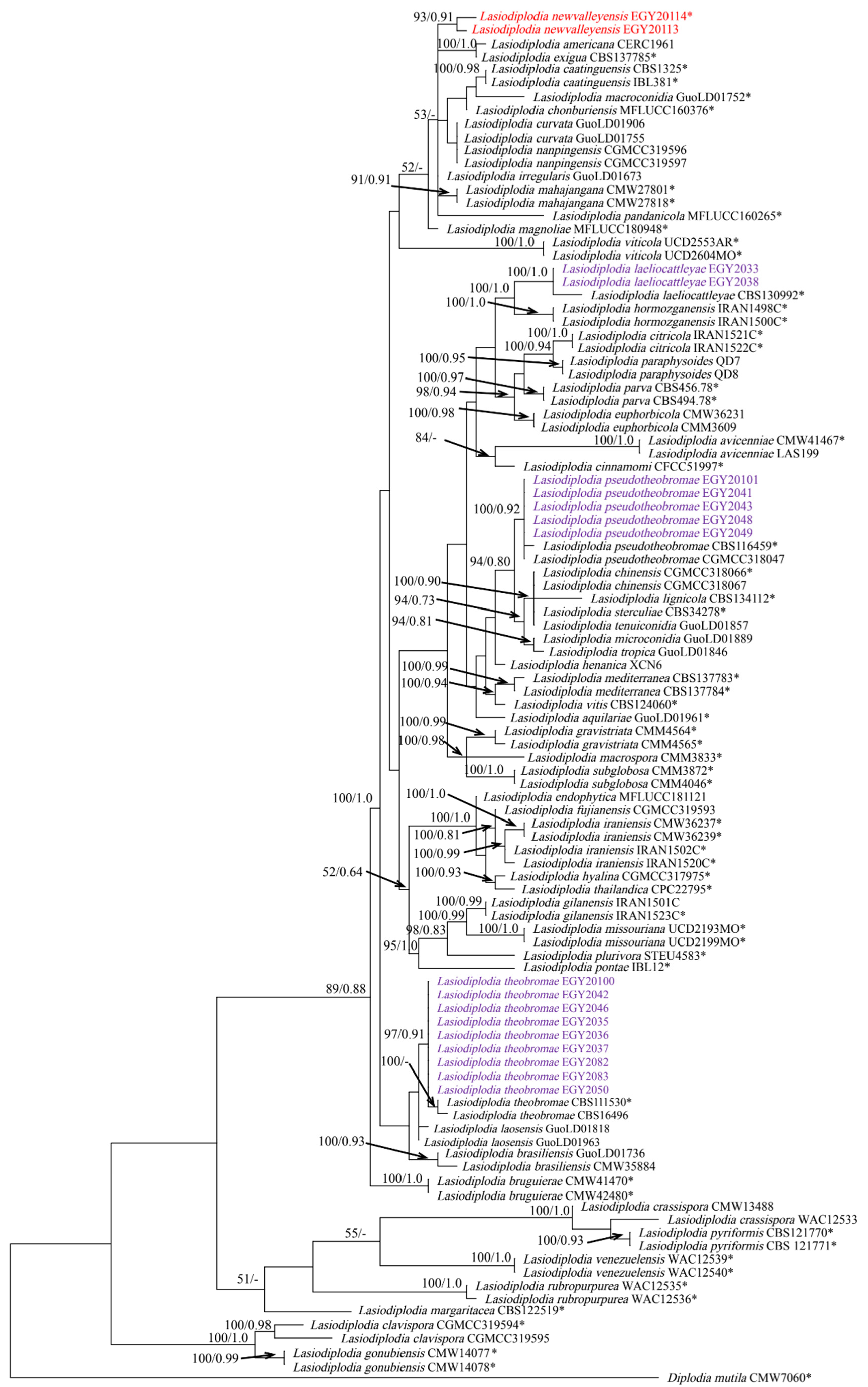
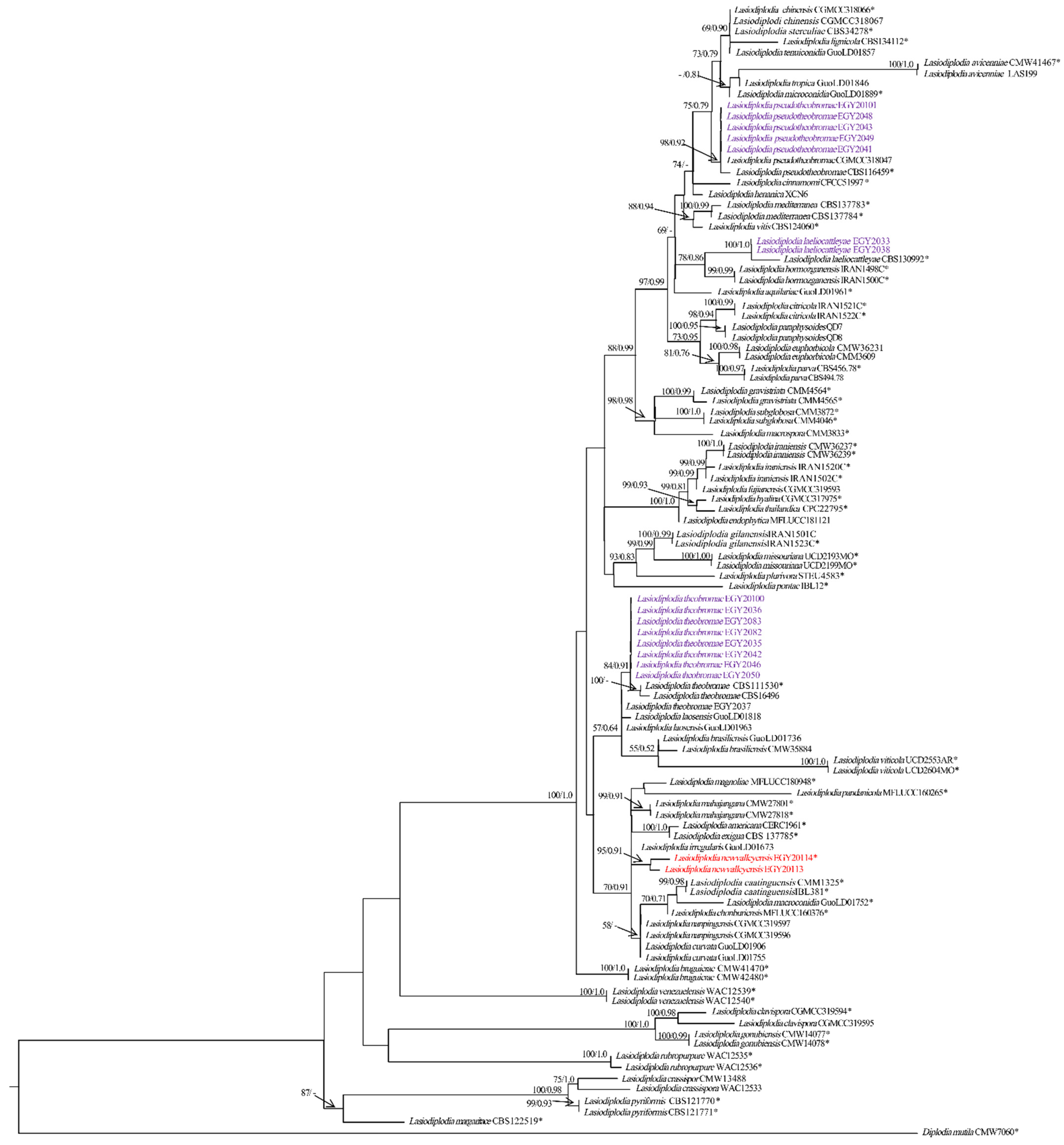
3.2. Phylogenetic Analyses
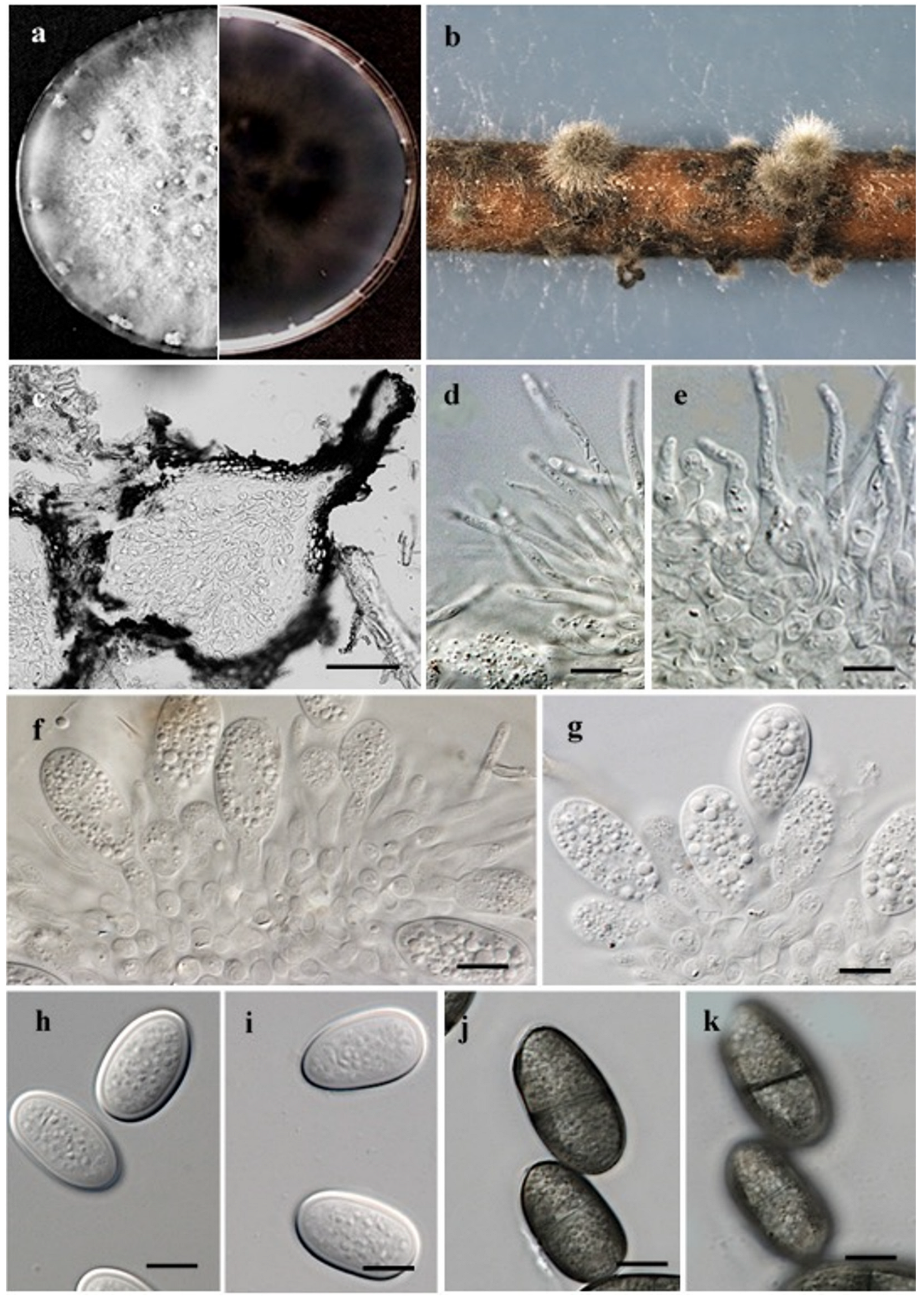
3.3. Taxonomy
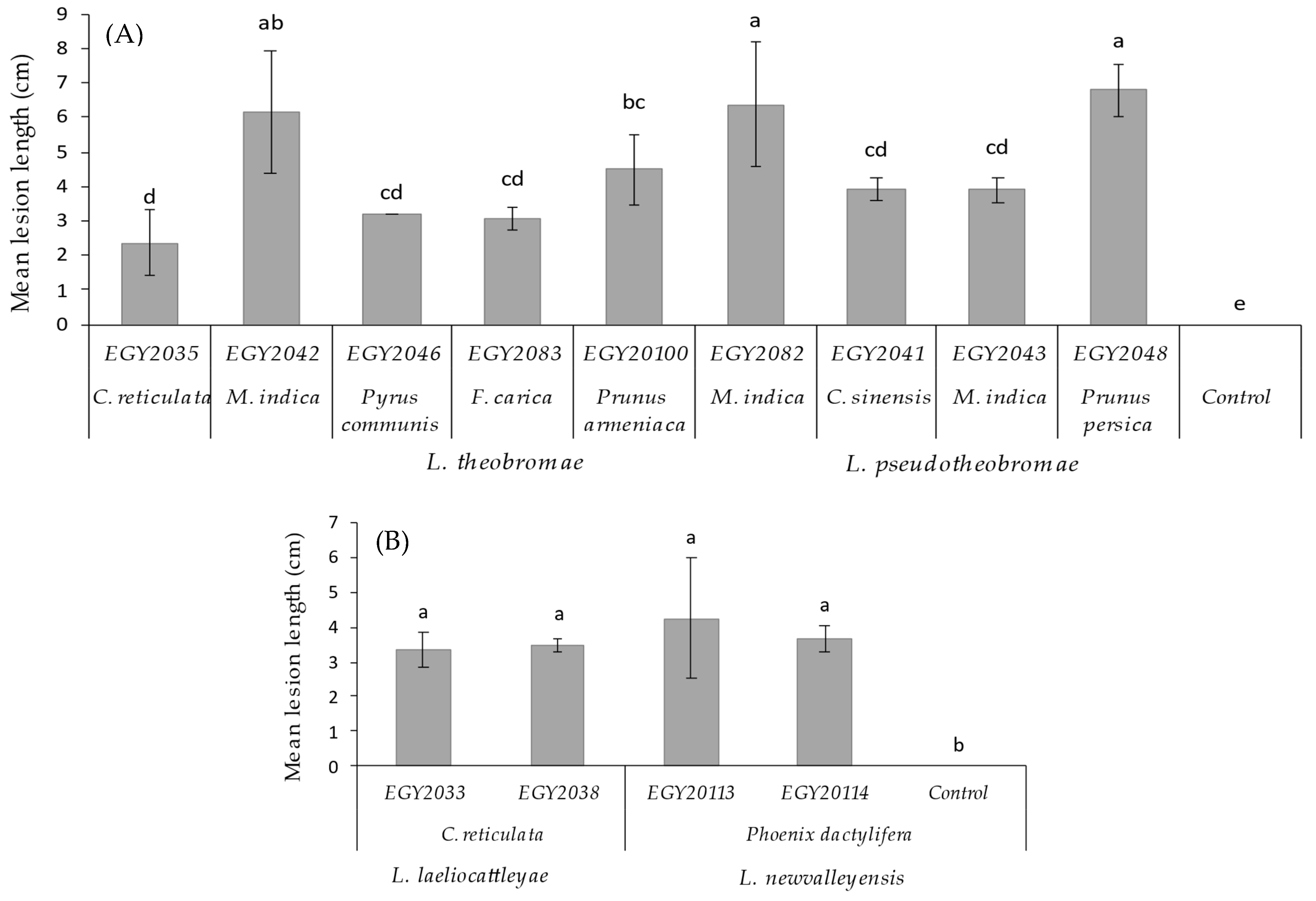
3.4. Pathogenicity Tests on Seedlings and Leaves
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Phillips, A.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2006, 55, 53–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freire, F.; Cardoso, J.; Viana, F.; Martins, M. Status of Lasiodiplodia theobromae as a plant pathogen in Brazil. Essentia 2011, 12, 53–71. [Google Scholar]
- Ismail, A.; Cirvilleri, G.; Polizzi, G.; Crous, P.; Groenewald, J.; Lombard, L. Lasiodiplodia species associated with dieback disease of mango (Mangifera indica) in Egypt. Australas. Plant Pathol. 2012, 41, 649–660. [Google Scholar] [CrossRef]
- Netto, M.S.; Assunção, I.P.; Lima, G.S.; Marques, M.W.; Lima, W.G.; Monteiro, J.H.; de Queiroz Balbino, V.; Michereff, S.J.; Phillips, A.J.; Câmara, M.P. Species of Lasiodiplodia associated with papaya stem-end rot in Brazil. Fungal Divers. 2014, 67, 127–141. [Google Scholar] [CrossRef]
- Coutinho, I.; Freire, F.; Lima, C.; Lima, J.; Gonçalves, F.; Machado, A.; Silva, A.; Cardoso, J. Diversity of genus Lasiodiplodia associated with perennial tropical fruit plants in northeastern Brazil. Plant Pathol. 2017, 66, 90–104. [Google Scholar] [CrossRef]
- Machado, A.R.; Pinho, D.B.; Pereira, O.L. Phylogeny, identification and pathogenicity of the Botryosphaeriaceae associated with collar and root rot of the biofuel plant jatropha curcas in Brazil, with a description of new species of Lasiodiplodia. Fungal Divers. 2014, 67, 231–247. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Bhoyroo, V.; Rampadarath, S.; Jeewon, R. Multigene phylogenetics and morphology reveal five novel Lasiodiplodia species associated with blueberries. Life 2021, 11, 657. [Google Scholar] [CrossRef]
- Crous, P.W.; Slippers, B.; Wingfield, M.J.; Rheeder, J.; Marasas, W.F.; Philips, A.J.; Alves, A.; Burgess, T.; Barber, P.; Groenewald, J.Z. Phylogenetic lineages in the Botryosphaeriaceae. Stud. Mycol. 2006, 55, 235–253. [Google Scholar] [CrossRef] [Green Version]
- Punithalingam, E. Plant Diseases Attributed to Botryodiplodia theobromae Pat; J. Cramer: Vaduz, Liechtenstein, 1980. [Google Scholar]
- Ragab, M.; Sabet, K.; Dawood, N. Botryodiplodia theobromae Pat. The cause of fruit rot and dieback of mango in A.R.E. Agrc. Res. Rev. Cairo 1971, 49, 81–97. [Google Scholar]
- Abdalla, M.; Safie, M.; El-Boghdady, M.; Soltan, H. Fruit coating with certain plant oils for controlling post-harvest diseases of mangoes with special reference to stem end rot. Egypt. J. Appl. Sci. 2003, 18, 116–136. [Google Scholar]
- Abd-El Ghani, H.S.; Fatouh, H.M. First record of sugar beet root rot disease caused by Botryodiplodia theobromae in Egypt. Egypt. J. Phytopathol. 2005, 33, 107–108. [Google Scholar]
- El-Goorani, M.A.; El-Meligi, M.A.; Elarosi, H.M.; Wasfy, E.H. Ceratocystis black peduncle rot of banana in Egypt/il marciume bruno del peduncolo della banana da ceratocystis in Egitto. Phytopathol. Mediter. 1972, 11, 193–194. [Google Scholar]
- Haggag, W.; Abou Rayya, M.; Kasim, N. First report of a canker disease of walnut caused by Botryodiplodia theobromae in Egypt. Plant Dis. 2007, 91, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diab, M.; Khalil, I.; Dawood, N.; El-Assiuty, E. Ear and grain rot of maize caused by Botryodiplodia theobromae pathogens in Egypt. Minufiya J. Agric. Res. 1984, 9, 129–138. [Google Scholar]
- Abo-El-Dahab, M.; El-Kazazz, S.; Shoeib, A.; El-Sheikh, M. Biochemical changes in citrus fruits infected with Botryodiplodia theobromae. J. Agric. Sci. Mansoura Univ. 1992, 7, 3524–3532. [Google Scholar]
- Haggag, W.M.; Nofal, M. Improving the biological control of Botryodiplodia disease on some annona cultivars using single or multi-bioagents in Egypt. Biol. Control 2006, 38, 341–349. [Google Scholar] [CrossRef]
- Kamhawy, M.A.; Mahrous, H.A.; Shalaby, M.S.; El-Sharabasy, S.M. Histopathology and control of Botryodiplodia theobromae rot of date palm off-shoot (Phoenix dactylifera L.) variety Zaghloul. Egypt. J. Agric. Res. 2005, 83, 1533–1546. [Google Scholar]
- Barakat, F.; Abdel Salam, A.; Abada, K.; Korra, A. In Pathogenicity of fungi associated with die-back of peach and some pome and stone fruit trees. In Proceedings of the Sixth Congress of Phytopathology, Cairo, Egypt, 5–7 March 1990; pp. 311–323. [Google Scholar]
- Leala, M.; El–Shahawy, I.; Tolba, I. Characterization of Lasiodiplodia isolates obtained from acid lime and Valencia orange in Egypt. Azhar J. Agric. Res. 2021, 46, 55–67. [Google Scholar] [CrossRef]
- Rehab, M.; Rashed, M.; Ammar, M.; El-Morsy, S. Dieback and sooty canker of ficus trees in Egypt and its control. Pak. J. Biol. Sci. 2014, 17, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Ismail, A.; Cirvilleri, G.; Lombard, L.; Crous, P.; Groenewald, J.; Polizzi, G. Characterisation of Neofusicoccum species causing mango dieback in Italy. J. Plant Pathol. 2013, 95, 549–557. [Google Scholar]
- Slippers, B.; Boissin, E.; Phillips, A.; Groenewald, J.Z.; Lombard, L.; Wingfield, M.J.; Postma, A.; Burgess, T.; Crous, P.W. Phylogenetic lineages in the Botryosphaeriales: A systematic and evolutionary framework. Stud. Mycol. 2013, 76, 31–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, Z.; He, W.; Zhang, Y. Does morphology matter in taxonomy of Lasiodiplodia? An answer from Lasiodiplodia hyalina sp. nov. Mycosphere 2017, 8, 1014–1027. [Google Scholar] [CrossRef]
- Abdollahzadeh, J.; Javadi, A.; Goltapeh, E.M.; Zare, R.; Phillips, A. Phylogeny and morphology of four new species of Lasiodiplodia from Iran. Pers.: Mol. Phylogeny Evol. Fungi 2010, 25, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slippers, B.; Crous, P.W.; Denman, S.; Coutinho, T.A.; Wingfield, B.D.; Wingfield, M.J. Combined multiple gene genealogies and phenotypic characters differentiate several species previously identified as Botryosphaeria dothidea. Mycologia 2004, 96, 83–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal rna genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA minipreparation: Version ii. Plant Mol. Biol. Rep. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous Ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the pcr to amplify conserved genes from filamentous Ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Alhudaib, K.A.; El-Ganainy, S.M.; Almaghasla, M.I.; Sattar, M.N. Characterization and control of Thielaviopsis punctulata on date palm in Saudi Arabia. Plants 2022, 11, 250. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. Mega11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. Mafft multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Swofford, D.L. Paup: Phylogenetic Analysis Using Parsimony (and Other Methods), 4.0 Beta. 2002. Available online: http://paup.csit.fsu.edu/ (accessed on 25 July 2022).
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. Mrbayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.; Pfeiffer, W.; Schwartz, T. Cipres Science Gateway Survey. Available online: http://www.phylo.org (accessed on 25 July 2022).
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. Iq-tree 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. Modelfinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Chernomor, O.; Von Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [Green Version]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. Ufboot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Snedecor, G.; Cochran, W. Statistical Methods, 7th ed.; Iowa State University Press: Ames, IA, USA, 1980. [Google Scholar]
- Linaldeddu, B.T.; Deidda, A.; Scanu, B.; Franceschini, A.; Serra, S.; Berraf-Tebbal, A.; Zouaoui Boutiti, M.; Ben Jamâa, M.; Phillips, A.J. Diversity of Botryosphaeriaceae species associated with grapevine and other woody hosts in Italy, Algeria and Tunisia, with descriptions of Lasiodiplodia exigua and Lasiodiplodia mediterranea sp. nov. Fungal Divers. 2015, 71, 201–214. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, S.; Zhao, L.; Sun, X.; He, W.; Zhang, Y.; Dai, Y.-C. Lasiodiplodia spp. Associated with Aquilaria Crassna in Laos. Mycol. Prog. 2019, 18, 683–701. [Google Scholar] [CrossRef]
- Begoude, B.; Slippers, B.; Wingfield, M.J.; Roux, J. Botryosphaeriaceae associated with Terminalia catappa in cameroon, south africa and madagascar. Mycol. Prog. 2010, 9, 101–123. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; McKenzie, E.H.; Bhat, D.J.; Phillips, A.J.; Wanasinghe, D.N.; Samarakoon, M.C.; Jayawardena, R.S.; Dissanayake, A.J.; Tennakoon, D.S. Fungal diversity notes 840–928: Micro-fungi associated with Pandanaceae. Fungal Divers. 2018, 93, 1–160. [Google Scholar] [CrossRef]
- de Silva, N.I.; Phillips, A.J.; Liu, J.-K.; Lumyong, S.; Hyde, K.D. Phylogeny and morphology of Lasiodiplodia species associated with Magnolia forest plants. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Li, G.; Liu, F.; Michailides, T.J. Novel species of Botryosphaeriaceae associated with shoot blight of pistachio. Mycologia 2015, 107, 780–792. [Google Scholar] [CrossRef] [PubMed]
- Trakunyingcharoen, T.; Lombard, L.; Groenewald, J.Z.; Cheewangkoon, R.; To-Anun, C.; Crous, P. Caulicolous Botryosphaeriales from Thailand. Pers. Mol. Phylogeny Evol. Fungi 2015, 34, 87–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalid, I.A. Pathological Studies on Lasiodiplodia theobromae the Causal Agent of Gummosis and Die-Back of Peach; Cairo University: Giza, Egypt, 2012. [Google Scholar]
- Sakalidis, M.L.; Ray, J.D.; Lanoiselet, V.; Hardy, G.E.; Burgess, T.I. Pathogenic Botryosphaeriaceae associated with Mangifera indica in the Kimberley region of Western Australia. Eur. J. Plant Pathol. 2011, 130, 379–391. [Google Scholar] [CrossRef] [Green Version]
- Marques, M.W.; Lima, N.B.; de Morais, M.A.; Michereff, S.J.; Phillips, A.J.; Câmara, M.P. Botryosphaeria, Neofusicoccum, Neoscytalidium and Pseudofusicoccum species associated with mango in Brazil. Fungal Divers. 2013, 61, 195–208. [Google Scholar] [CrossRef]
- Li, G.; Liu, F.; Li, J.; Liu, Q.; Chen, S. Characterization of Botryosphaeria dothidea and Lasiodiplodia pseudotheobromae from English Walnut in China. J. Phytopathol. 2016, 164, 348–353. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Zhang, W.; Mei, L.; Chukeatirote, E.; Yan, J.Y.; Li, X.; Hyde, K.D. Lasiodiplodia pseudotheobromae causes pedicel and peduncle discolouration of grapes in China. Australas. Plant Dis. Notes 2015, 10, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Phillips, A.; Alves, A.; Pennycook, S.; Johnston, P.; Ramaley, A.; Akulov, A.; Crous, P. Resolving the phylogenetic and taxonomic status of dark-spored teleomorph genera in the Botryosphaeriaceae. Pers. Mol. Phylogeny Evol. Fungi 2008, 21, 29–55. [Google Scholar] [CrossRef]
- Alves, A.; Crous, P.W.; Correia, A.; Phillips, A. Morphological and molecular data reveal cryptic speciation in Lasiodiplodia theobromae. Fungal Divers. 2008, 28, 1–13. [Google Scholar]
- Rodríguez-Gálvez, E.; Guerrero, P.; Barradas, C.; Crous, P.W.; Alves, A. Phylogeny and pathogenicity of Lasiodiplodia species associated with dieback of mango in Peru. Fungal Biol. 2017, 121, 452–465. [Google Scholar] [CrossRef] [PubMed]
- Rosado, A.W.C.; Machado, A.R.; Freire, F.d.C.O.; Pereira, O.L. Phylogeny, identification, and pathogenicity of Lasiodiplodia associated with postharvest stem-end rot of coconut in Brazil. Plant Dis. 2016, 100, 561–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, A.J.; Hyde, K.D.; Alves, A.; Liu, J.-K.J. Families in Botryosphaeriales: A phylogenetic, morphological and evolutionary perspective. Fungal Divers. 2019, 94, 1–22. [Google Scholar] [CrossRef]
- Huda-Shakirah, A.R.; Mohamed Nor, N.M.I.; Zakaria, L.; Leong, Y.-H.; Mohd, M.H. Lasiodiplodia theobromae as a causal pathogen of leaf blight, stem canker, and pod rot of Theobroma cacao in Malaysia. Sci. Rep. 2022, 12, 8966. [Google Scholar] [CrossRef]
- Berraf-Tebbal, A.; Mahamedi, A.E.; Aigoun-Mouhous, W.; Špetík, M.; Čechová, J.; Pokluda, R.; Baranek, M.; Eichmeier, A.; Alves, A. Lasiodiplodia mitidjana sp. nov. and other Botryosphaeriaceae species causing branch canker and dieback of Citrus sinensis in Algeria. PLoS ONE 2020, 15, e0232448. [Google Scholar] [CrossRef]
- Liang, L.; Li, H.; Zhou, L.; Chen, F. Lasiodiplodia pseudotheobromae causes stem canker of chinese hackberry in China. J. For. Res. 2020, 31, 2571–2580. [Google Scholar] [CrossRef] [Green Version]
- Pennycook, S.; Samuels, G. Botryosphaeria and Fusicoccum species associated with ripe fruit rot of Actinidia deliciosa (kiwifruit) in New Zealand. Mycotaxon 1985, 24, 445–458. [Google Scholar]
- Burgess, T.I.; Barber, P.A.; Mohali, S.; Pegg, G.; de Beer, W.; Wingfield, M.J. Three new Lasiodiplodia spp. From the tropics, recognized based on DNA sequence comparisons and morphology. Mycologia 2006, 98, 423–435. [Google Scholar] [CrossRef]
- Damm, U.; Crous, P.W.; Fourie, P.H. Botryosphaeriaceae as potential pathogens of Prunus species in South africa, with descriptions of Diplodia africana and Lasiodiplodia plurivora sp. nov. Mycologia 2007, 99, 664–680. [Google Scholar] [CrossRef]
- Xiao, X.; Wang, W.; Crous, P.; Wang, H.; Jiao, C.; Huang, F.; Pu, Z.; Zhu, Z.; Li, H. Species of Botryosphaeriaceae associated with citrus branch diseases in China. Pers. Mol. Phylogeny Evol. Fungi 2021, 47, 106–135. [Google Scholar] [CrossRef]
- Kenfaoui, J.; Lahlali, R.; Mennani, M.; Radouane, N.; Goura, K.; El Hamss, H.; El Ghadraoui, L.; Fontaine, F.; Tahiri, A.; Barka, E.A. Botryosphaeria dieback (Lasiodiplodia viticola): An imminent emerging threat to the moroccan vineyards. Plants 2022, 11, 2167. [Google Scholar] [CrossRef] [PubMed]
- Moubasher, H.A.; Balbool, B.A.; Helmy, Y.A.; Alsuhaibani, A.M.; Atta, A.A.; Sheir, D.H.; Abdel-Azeem, A.M. Insights into asparaginase from endophytic fungus Lasiodiplodia theobromae: Purification, characterization and antileukemic activity. Int. J. Environ. Res. Public Health 2022, 19, 680. [Google Scholar] [CrossRef] [PubMed]
- Salvatore, M.M.; Alves, A.; Andolfi, A. Secondary metabolites of Lasiodiplodia theobromae: Distribution, chemical diversity, bioactivity, and implications of their occurrence. Toxins 2020, 12, 457. [Google Scholar] [CrossRef]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- Pérez, C.A.; Wingfield, M.J.; Slippers, B.; Altier, N.A.; Blanchette, R.A. Endophytic and canker-associated Botryosphaeriaceae occurring on non-native eucalyptus and native Myrtaceae trees in Uruguay. Fungal Divers. 2010, 41, 53–69. [Google Scholar] [CrossRef] [Green Version]
- Nur-Shakirah, A.O.; Khadijah, M.S.; Kee, Y.J.; Huda-Shakirah, A.R.; Mohd Hafifi, A.B.; Nurul-Aliyaa, Y.A.; Chew, B.L.; Zakaria, L.; Mohamed Nor, N.M.I.; Sreeramanan, S. Characterization of Lasiodiplodia species causing leaf blight, stem rot and fruit rot of fig (Ficus carica) in Malaysia. Plant Pathol. 2022, 71, 1594–1605. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.; Leavitt, G.; Guerrero, J.; Guevara, J.; Gubler, W. Identification and pathogenicity of Lasiodiplodia theobromae and Diplodia seriata, the causal agents of bot canker disease of grapevines in Mexico. Plant Dis. 2008, 92, 519–529. [Google Scholar] [CrossRef]
- Jacobs, R. Characterisation of Botryosphaeria Species from Mango in South Africa; University of Pretoria: Pretoria, South Africa, 2002. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Host | Country | GenBank Accession Numbers | ||
|---|---|---|---|---|---|---|
| ITS | tef1-α | tub2 | ||||
| L. aquilariae | GuoLD01961 * | Aquilaria crassna | Laos | KY783442 | KY848600 | - |
| L. avicenniae | CMW 41467 * | Avocennia marina | South Africa | KP860835 | KP860680 | KP860758 |
| L. avicenniae | LAS 199 | Avocennia marina | South Africa | KU587957 | KU587947 | KU587868 |
| L. americana | CERC 1961 = CFCC 50065 * | Pistachia vera | China | KP217059 | KP217067 | KP217075 |
| L. brasiliensis | GuoLD01736 | Carica papaya | Brazil | KY783475 | KY848612 | KY848556 |
| L. brasiliensis | CMW35884 | Adansonia madagascariensis | Madagascar | KU887094 | KU886972 | KU887466 |
| L. bruguierae | CMW41470 * | Bruguiera gymnorrhiza | South Africa | KP860833 | KP860678 | KP860756 |
| L. bruguierae | CMW42480 * | Bruguiera gymnorrhiza | South Africa | KP860832 | KP860677 | KP860755 |
| L. caatinguensis | CMM1325 * | Citrus sinensis | Brazil | KT154760 | KT008006 | KT154767 |
| L. caatinguensis | IBL381 * | Spondias purpurea | Brazil | KT154757 | KT154751 | KT154764 |
| L. chinensis | CGMCC3.18066 * | Hevea brasiliensis | China | KX499899 | KX499937 | KX500012 |
| L. chinensis | CGMCC3.18067 | Sterculia lychnophora | China | KX499901 | KX499939 | KX500014 |
| L. chonburiensis | MFLUCC 16-0376 * | Pandanaceae | Thailand | MH275066 | MH412773 | MH412742 |
| L. cinnamomi | CFCC 51997 * | Cinnamomum camphora | China | MG866028 | MH236799 | MH236797 |
| L. citricola | IRAN1521C * | Citrus sp. | Iran | GU945353 | GU945339 | KU887504 |
| L. citricola | IRAN1522C * | Citrus sp. | Iran | GU945354 | GU945340 | KU887505 |
| L. clavispora | CGMCC 3.19594 * | Vaccinium uliginosum | China | MK802166 | OL773697 | MK816339 |
| L. clavispora | CGMCC 3.19595 | Vaccinium uliginosum | China | MK802165 | OL773696 | MK816338 |
| L. crassispora | CMW 13488 | Eucalyptus urophylla | Venezuela | DQ103552 | DQ103559 | KU887507 |
| L. crassispora | WAC12533 | Santalum album | Australia | DQ103550 | DQ103557 | - |
| L. curvata | GuoLD01755 | Aquilaria crassna | Laos | KY783443 | KY848601 | KY848532 |
| L. curvata | GuoLD01906 | Aquilaria crassna | Laos | KY783437 | KY84859 | KY848529 |
| L. euphorbicola | CMW36231 * | Adansonia digitata | Botswana | KU887187 | KU887063 | KU887494 |
| L. euphorbicola | CMW 3609 * | Adansonia digitata | Zimbabwe | KF234543 | KF226689 | KF254926 |
| L. endophytica | MFLUCC 18-1121 | Magnolia acuminata | China | MK501838 | MK584572 | MK550606 |
| L. exigua | IBL 104 = CBS 137785 * | Retama raetam | Tunisia | KJ638317 | KJ638336 | KU887509 |
| L. fujianensis | CGMCC3.19593 | Vaccinium uliginosum | China | MK802164 | MK887178 | MK816337 |
| L. gilanensis | IRAN 1501C | Unknown | Iran | GU945352 | GU945341 | KU887510 |
| L. gilanensis | IRAN 1523C * | Unknown | Iran | GU945351 | GU945342 | KU887511 |
| L. gonubiensis | CMW 14077 * | Syzygium cordatum | South Africa | AY639595 | DQ103566 | DQ458860 |
| L. gonubiensis | CMW 14078 * | Syzygium cordatum | South Africa | AY639594 | DQ103567 | EU673126 |
| L. gravistriata | CMM 4564 * | Anacardium humile | Brazil | KT250949 | KT250950 | - |
| L. gravistriata | CMM 4565 * | Anacardium humile | Brazil | KT250947 | KT266812 | - |
| L. henanica | XCN6 = CGMCC 3.19176 | Vaccinium uliginosum | China | MH729351 | MH729357 | MH729360 |
| L. hormozganensis | IRAN 1498C * | Mangifera indica | Iran | GU945356 | GU945344 | KU887514 |
| L. hormozganensis | IRAN 1500C * | Olea sp. | Iran | GU945355 | GU945343 | KU887515 |
| L. hyalina | CGMCC 3.17975 * | Acacia confusa | China | KX499879 | KX499917 | KX499992 |
| L. iraniensis | CMW 36237 * | Adansonia digitata | Mozambique | KU887121 | KU886998 | KU887499 |
| L. iraniensis | CMW 36239 * | Adansonia digitata | Mozambique | KU887123 | KU887000 | KU887501 |
| L. iraniensis | IRAN 1502C * | Juglans sp. | Iran | GU945347 | GU945335 | KU887517 |
| L. iraniensis | IRAN 1520C * | Salvadora persica | Iran | GU945348 | GU945336 | KU887516 |
| L. irregularis | GuoLD01673 | Aquilaria crassna | Laos | KY783472 | KY848610 | KY848553 |
| L. laeliocattleyae | CBS 130992 * | Mangifera indica | Egypt | JN814397 | JN814424 | KU887508 |
| L. laeliocattleyae | EGY2033 | Citrus reticulata | Egypt | ON392181 | OP080238 | OP080255 |
| L. laeliocattleyae | EGY2038 | Citrus reticulata | Egypt | ON392185 | OP080242 | OP080259 |
| L. laosensis | GuoLD01818 | Aquilaria crassna | Laos | KY783471 | KY848609 | KY848552 |
| L. laosensis | GuoLD01963 | Aquilaria crassna | Laos | KY783450 | KY848603 | KY848536 |
| L. lignicola | CBS 134112 * | dead wood | Thailand | JX646797 | KU887003 | JX646845 |
| L. macroconidica | GuoLD01752 * | Aquilaria crassna | Laos | KY783438 | KY848597 | KY848530 |
| L. macrospora | CMM3833 * | Jatropha curcas | Brazil | KF234557 | KF226718 | KF254941 |
| L. magnoliae | MFLUCC18-0948 * | Magnolia candolii | China | MK499387 | MK568537 | MK521587 |
| L. mahajangana | CMW 27801 * | Terminalia catappa | Madagascar | FJ900595 | FJ900641 | FJ900630 |
| L. mahajangana | CMW 27818 * | Terminalia catappa | Madagascar | FJ900596 | FJ900642 | FJ900631 |
| L. margaritacea | CBS 122519 * | Adansonia gibbosa | Australia | EU144050 | EU144065 | KU887520 |
| L. mediterranea | CBS 137783 * | Quercus ilex | Italy | KJ638312 | KJ638331 | KU887521 |
| L. mediterranea | CBS 137784 * | Vitis vinifera | Italy | KJ638311 | KJ638330 | KU887522 |
| L. microcondia | GuoLD01889 | Aquilaria crassna | Laos | KY783441 | KY848614 | - |
| L. missouriana | UCD 2193MO * | Vitis vinifera | USA | HQ288225 | HQ288267 | HQ288304 |
| L. missouriana | UCD 2199MO * | Vitis vinifera | USA | HQ288226 | HQ288268 | HQ288305 |
| L. nanpingensis | CGMCC3.19597 | Vaccinium uliginosum | China | MK802168 | OL773699 | MK816341 |
| L. nanpingensis | CGMCC319596 | Vaccinium uliginosum | China | MK802168 | OL773698 | MK816340 |
| L. newvalleyensis | EGY20113 * | Phoenix dactylifera | Egypt | ON392175 | OP080253 | OP080271 |
| L. newvalleyensis | EGY20114 * | Phoenix dactylifera | Egypt | ON392180 | OP080254 | OP080272 |
| L. pandanicola | MFLUCC 16-0265 * | Pandanaceae | Thailand | MH275068 | MH412774 | - |
| L. paraphysoides | CGMCC 3.19174 = QD7 | Vaccinium uliginosum | China | MH729349 | MH729355 | MH729358 |
| L. paraphysoides | CGMCC 3.19175 = QD8 | Vaccinium uliginosum | China | MH729350 | MH729356 | MH729359 |
| L. parva | CBS 456.78 * | Cassava field-soil | USA | EF622083 | EF622063 | KU887523 |
| L. parva | CBS 494.78 | Cassava field-soil | USA | EF622084 | EF622064 | EU673114 |
| L. plurivora | STE-U 4583 */CBS 121103 | Vitis vinifera | South Africa | AY343482 | EF445396 | KU887525 |
| L. pontae | IBL12 = CMM1277 * | Spondias purpurea | Brazil | KT151794 | KT151791 | KT151797 |
| L. pseudotheobromae | CBS 116459 * | Gmelina arborea | Costa Rica | EF622077 | EF622057 | EU673111 |
| L. pseudotheobromae | CGMCC 3.18047 | Pteridium aquilinum | China | KX499876 | KX499914 | KX499989 |
| L. pseudotheobromae | EGY2041 | Citrus sinensis | Egypt | ON392168 | OP080243 | OP080260 |
| L. pseudotheobromae | EGY2043 | Mangifera indica | Egypt | ON392170 | OP080245 | OP080262 |
| L. pseudotheobromae | EGY2048 | Prunus persica | Egypt | ON392172 | OP080247 | OP080264 |
| L. pseudotheobromae | EGY2049 | Mangifera indica | Egypt | ON392173 | OP080248 | OP080265 |
| L. pseudotheobromae | EGY20101 | Mangifera indica | Egypt | ON392179 | OP080252 | OP080270 |
| L. pyriformis | CBS 121770 * | Acacia mellifera | Namibia | EU101307 | EU101352 | KU887527 |
| L. pyriformis | CBS 121771 * | Acacia mellifera | Namibia | EU101308 | EU101353 | KU887528 |
| L. rubropurpurea | WAC 12535 * | Eucalyptus grandis | Australia | DQ103553 | DQ103571 | EU673136 |
| L. rubropurpurea | WAC 12536 * | Eucalyptus grandis | Australia | DQ103554 | DQ103572 | KU887530 |
| L. sterculiae | CBS342.78 * | Sterculia oblonga | Germany | KX464140 | KX464634 | KX464908 |
| L. subglobosa | CMM3872 * | Jatropha curcas | Brazil | KF234558 | KF226721 | KF254942 |
| L. subglobosa | CMM4046 * | Jatropha curcas | Brazil | KF234560 | KF226723 | KF254944 |
| L. tenuiconidia | GuoLD01857 | Aquilaria crassna | Laos | KY783466 | KY848619 | KY848586 |
| L. thailandica | CPC22795 * | Albizia chinensis | China | KJ193637 | KJ193681 | KY751301 |
| L. theobromae | CBS 111530 * | Unknown | Unknown | EF622074 | EF622054 | KU887531 |
| L. theobromae | CBS 164.96 | Fruit on coral reef coast | Papua New Guinea | AY640255 | AY640258 | KU887532 |
| L. theobromae | EGY2035 | Citrus reticulata | Egypt | ON392182 | OP080239 | OP080256 |
| L. theobromae | EGY2036 | Citrus reticulata | Egypt | ON392183 | OP080240 | OP080257 |
| L. theobromae | EGY2037 | Citrus reticulata | Egypt | ON392184 | OP080241 | OP080258 |
| L. theobromae | EGY2042 | Mangifera indica | Egypt | ON392169 | OP080244 | OP080261 |
| L. theobromae | EGY2046 | Pyrus communis | Egypt | ON392171 | OP080246 | OP080263 |
| L. theobromae | EGY2050 | Pyrus communis | Egypt | ON392174 | OP080249 | OP080266 |
| L. theobromae | EGY2082 | Mangifera indica | Egypt | ON392176 | OP080237 | OP080267 |
| L. theobromae | EGY2083 | Ficus carica | Egypt | ON392177 | OP080250 | OP080268 |
| L. theobromae | EGY20100 | Prunus armeniaca | Egypt | ON392178 | OP080251 | OP080269 |
| L. tropica | GuoLD01846 | Aquilaria crassna | Laos | KY783454 | KY848616 | KY848540 |
| L. venezuelensis | WAC 12539 * | Acacia mangium | Venezuela | DQ103547 | DQ103568 | KU887533 |
| L. venezuelensis | WAC 12540 * | Acacia mangium | Venezuela | DQ103548 | DQ103569 | KU887534 |
| L. viticola | UCD 2553AR * | Vitis sp. | USA | HQ288227 | HQ288269 | HQ288306 |
| L. viticola | UCD 2604MO * | Vitis sp. | USA | HQ288228 | HQ288270 | HQ288307 |
| L. vitis | CBS 124060 * | Vitis vinifera | Italy | KX464148 | KX464642 | KX464917 |
| Diplodia mutila | CMW 7060 * | Fraxinus excelsior | Netherlands | AY236955 | AY236904 | AY236933 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Ganainy, S.M.; Ismail, A.M.; Iqbal, Z.; Elshewy, E.S.; Alhudaib, K.A.; Almaghasla, M.I.; Magistà, D. Diversity among Lasiodiplodia Species Causing Dieback, Root Rot and Leaf Spot on Fruit Trees in Egypt, and a Description of Lasiodiplodia newvalleyensis sp. nov. J. Fungi 2022, 8, 1203. https://doi.org/10.3390/jof8111203
El-Ganainy SM, Ismail AM, Iqbal Z, Elshewy ES, Alhudaib KA, Almaghasla MI, Magistà D. Diversity among Lasiodiplodia Species Causing Dieback, Root Rot and Leaf Spot on Fruit Trees in Egypt, and a Description of Lasiodiplodia newvalleyensis sp. nov. Journal of Fungi. 2022; 8(11):1203. https://doi.org/10.3390/jof8111203
Chicago/Turabian StyleEl-Ganainy, Sherif Mohamed, Ahmed Mahmoud Ismail, Zafar Iqbal, Eman Said Elshewy, Khalid A. Alhudaib, Mustafa I. Almaghasla, and Donato Magistà. 2022. "Diversity among Lasiodiplodia Species Causing Dieback, Root Rot and Leaf Spot on Fruit Trees in Egypt, and a Description of Lasiodiplodia newvalleyensis sp. nov." Journal of Fungi 8, no. 11: 1203. https://doi.org/10.3390/jof8111203