
A Novel Gammapartitivirus That Causes Changes in Fungal Development and Multi-Stress Tolerance to Important Medicinal Fungus Cordyceps chanhua
 , and
, and 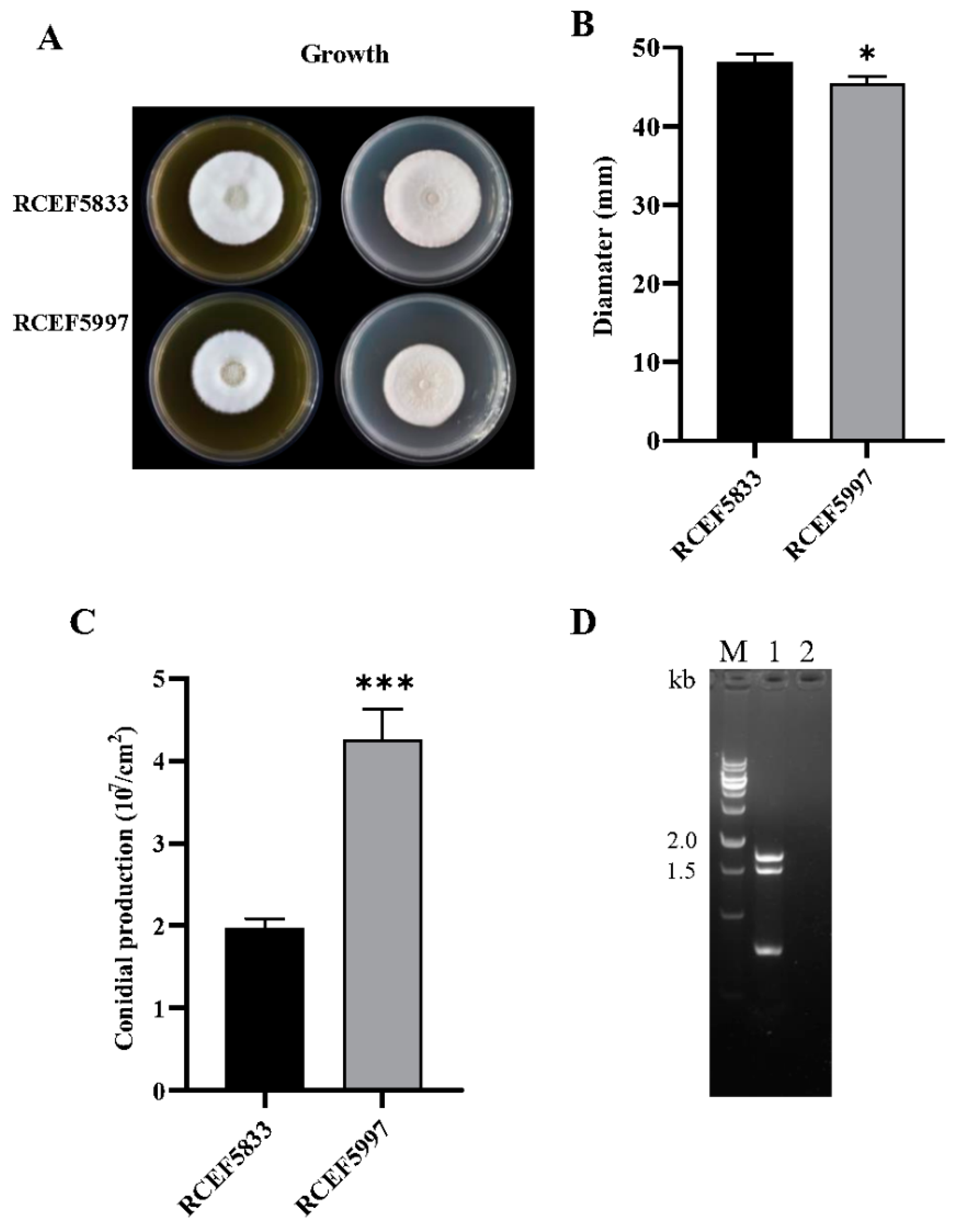{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolates and Culture
2.2. dsRNA Extraction and Purification
2.3. RNA-Sequencing, Molecular Cloning, and Sequence Analysis
2.4. Horizontal Transmission of CchPV1
2.5. Phenotype Assays
2.6. Statistical Analysis
3. Results
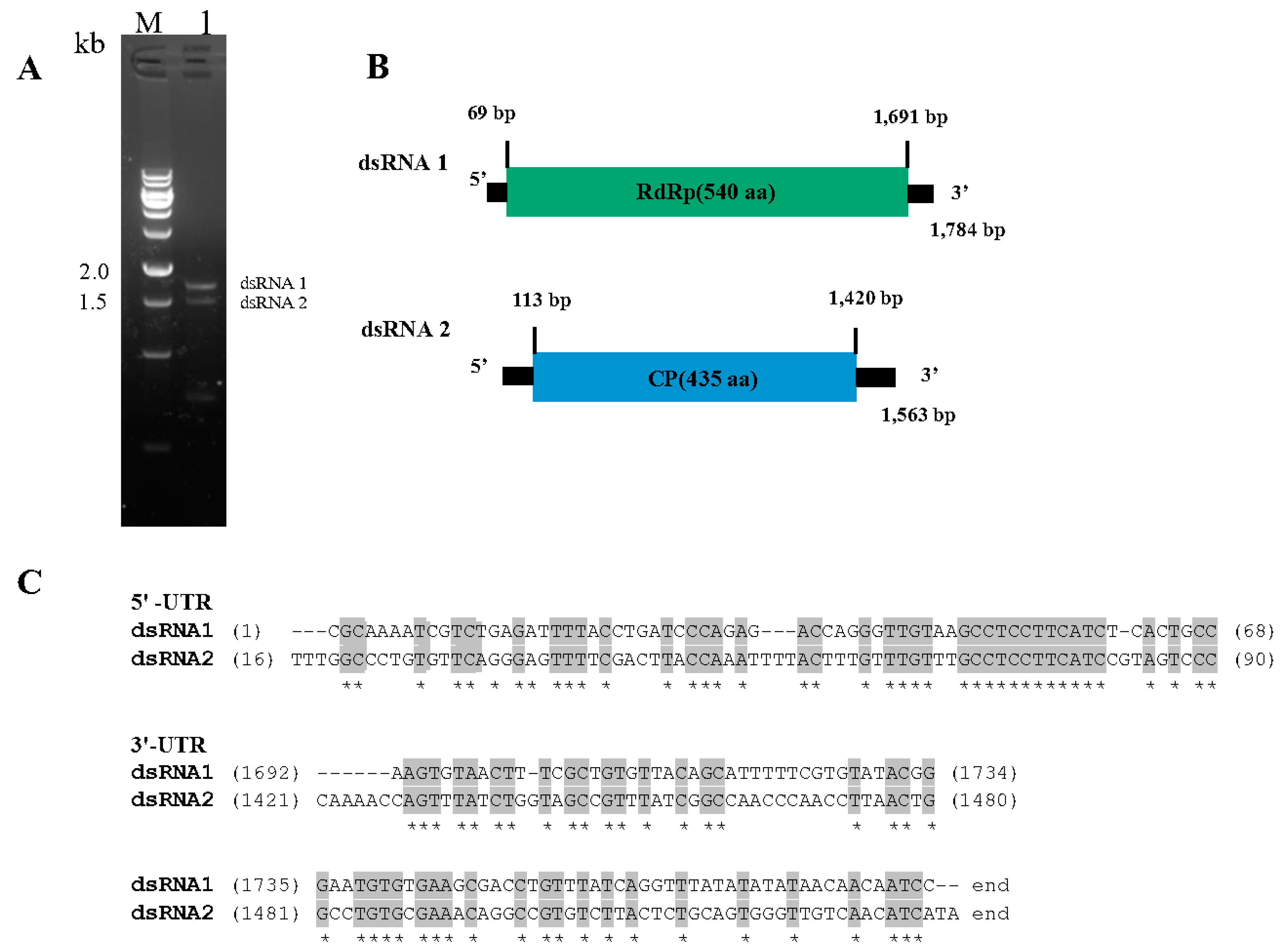
3.1. dsRNA Segments in C. chanhua RCEF5997
3.2. Genome Structure of CchPV1
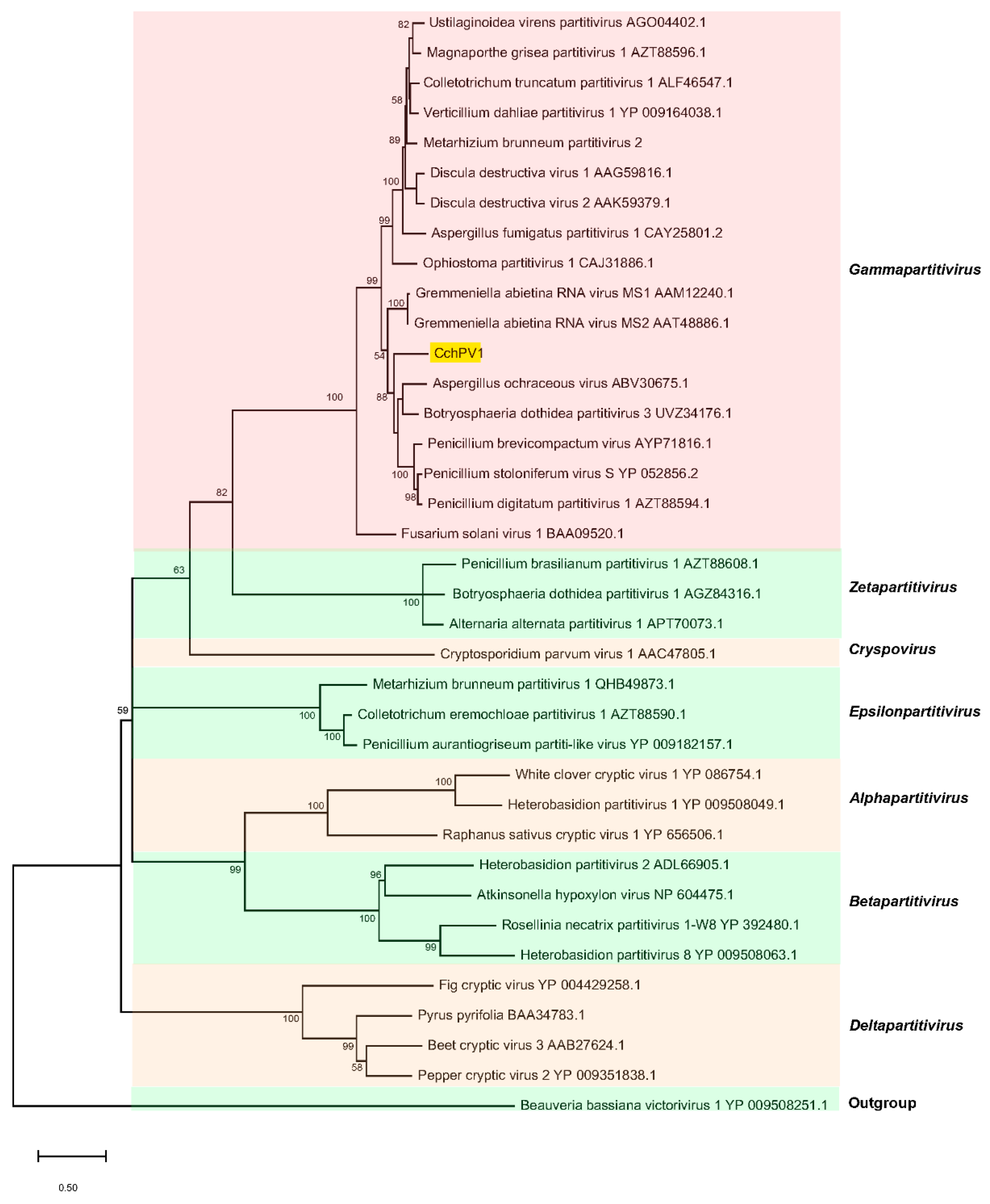
3.3. Taxonomic and Phylogenetic Position of CchPV1
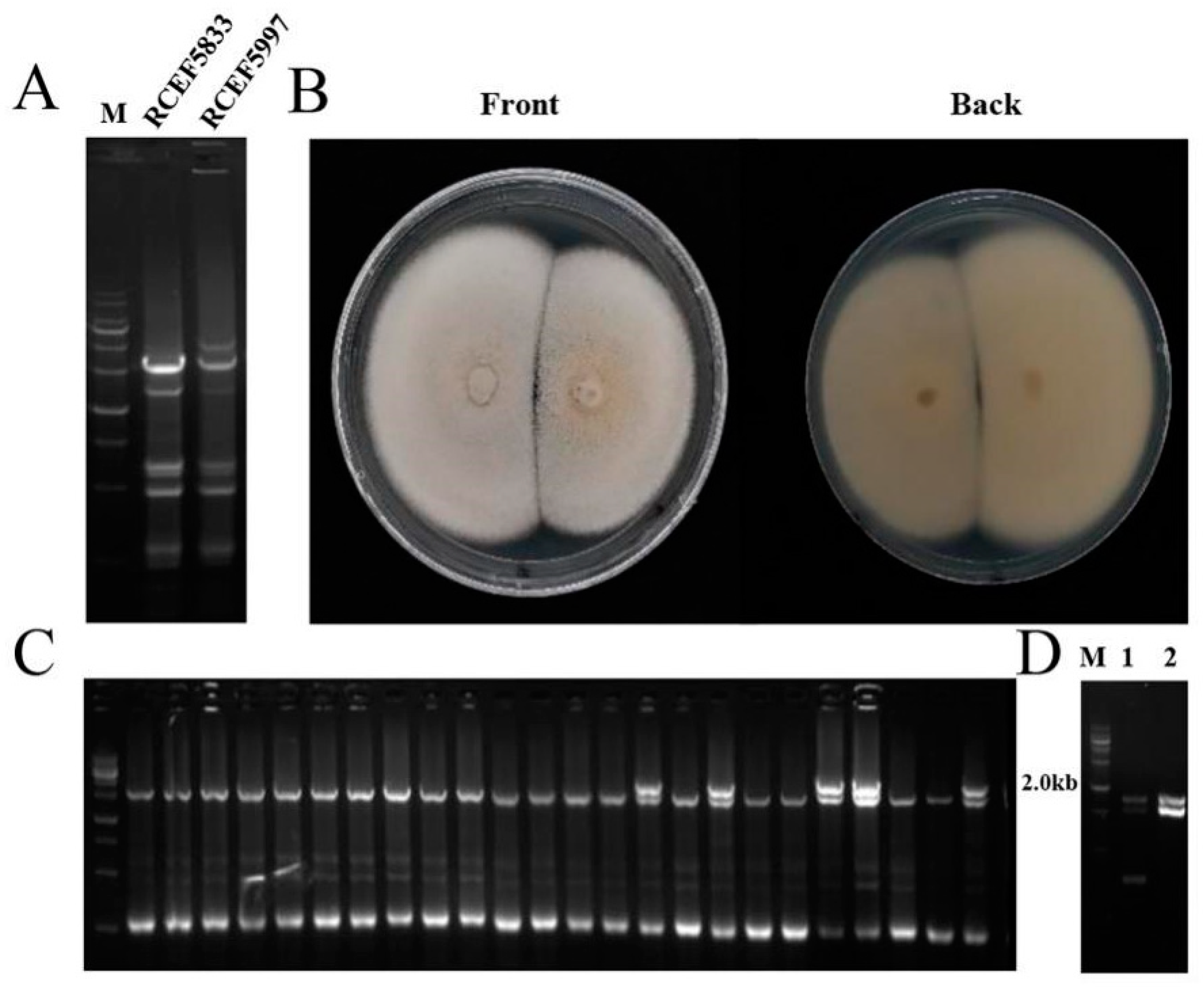
3.4. Horizontal Transmission of CchPV1 from C. chanhua RCEF5997
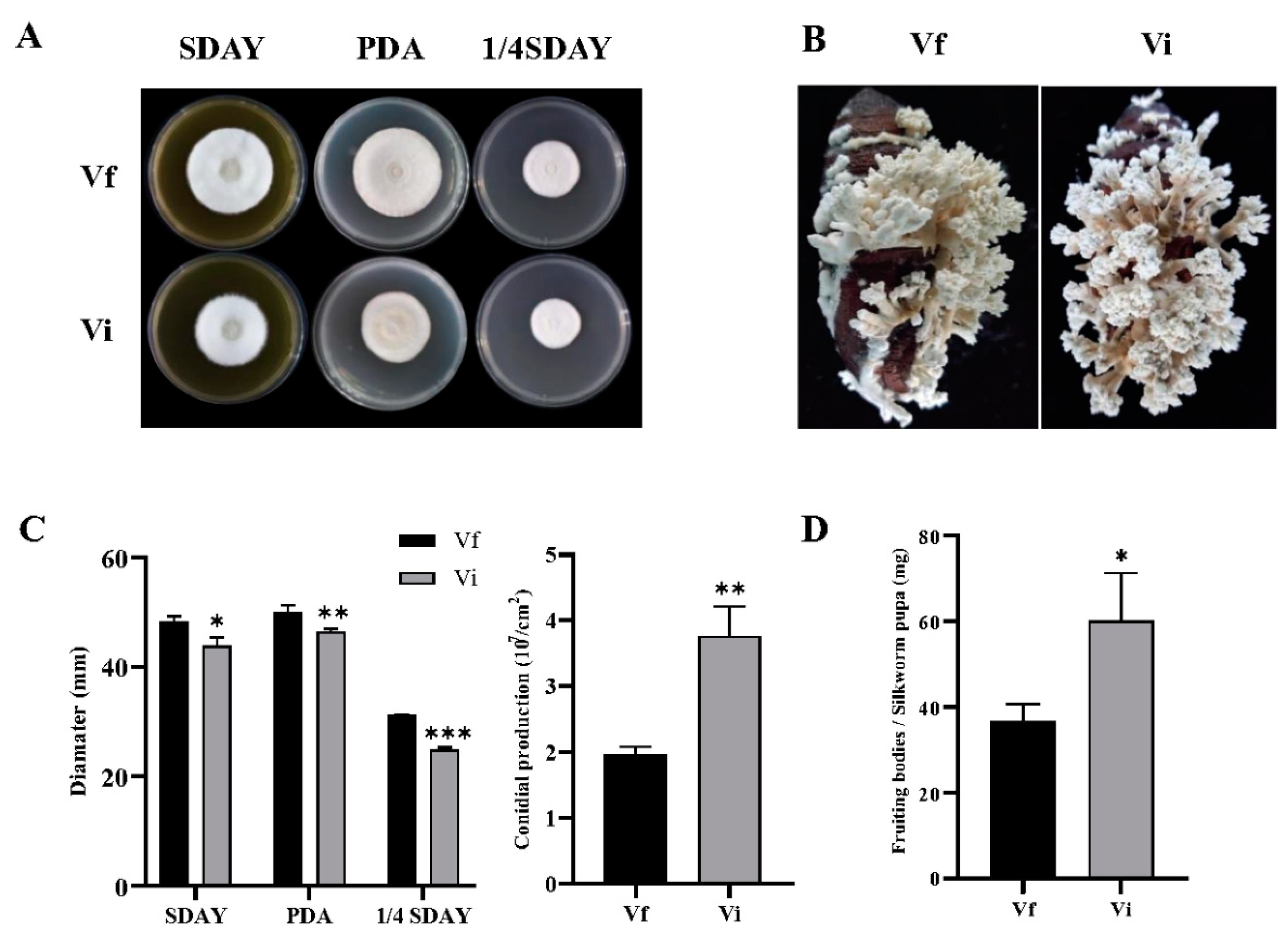
3.5. CchPV1 Infection Slows Down the Growth Rate of the Host, but Increases the Conidiation and Formation of Fruit Bodies of the Host
3.6. CchPV1 Infection Weakens Multi-Stress Tolerance of the Host
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, I.C.; Lin, S.; Tsai, Y.T.; Hsu, J.H.; Chen, Y.L.; Lin, W.H.; Chen, C.C. Cordyceps cicadae mycelia and its active compound HEA exert beneficial effects on blood glucose in type 2 diabetic db/db mice. J. Sci. Food Agric. 2019, 99, 606–612. [Google Scholar] [CrossRef]
- Li, Z.; Luan, F.; Hywel-Jones Nigel, L.; Zhang, S.; Chen, M.; Huang, B. Biodiversity of cordycipitoid fungi associated with Isaria cicadae Miquel II: Teleomorph discovery and nomenclature of chanhua, an important medicinal fungus in China. Mycosystema 2021, 40, 1–12. [Google Scholar]
- Nxumalo, W.; Elateeq, A.A.; Sun, Y. Can Cordyceps cicadae be used as an alternative to Cordyceps militaris and Cordyceps sinensis?—A review. J. Ethnopharmacol. 2020, 257, 112879. [Google Scholar] [CrossRef]
- Dong, C.; Li, W.; Li, Z.; Yan, W.; Li, T. Cordyceps industry in China: Current status, challenges and perspectives-Jinhu declaration for Cordyceps industry development. Mycosystema 2016, 35, 1–15. [Google Scholar]
- Li, C.R.; Wang, Y.Q.; Cheng, W.M.; Chen, Z.A.; Hywel-Jones, N.; Li, Z.Z. Review on research progress and prospects of cicada flower, Isaria cicadae (Ascomycetes). Int. J. Med. Mushrooms 2021, 23, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Ochoa, W.F.; Li, H.; Havens, W.M.; Nibert, M.L.; Ghabrial, S.A.; Baker, T.S. Structure of Fusarium poae virus 1 shows conserved and variable elements of partitivirus capsids and evolutionary relationships to picobirnavirus. J. Struct. Biol. 2010, 172, 363–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, J.H.; Dong, L.P.; Lin, L.; Ochoa, W.F.; Sinkovits, R.S.; Havens, W.M.; Nibert, M.L.; Baker, T.S.; Ghabrial, S.A.; Tao, Y.Z.J. Atomic structure reveals the unique capsid organization of a dsRNA virus. Proc. Natl. Acad. Sci. USA 2009, 106, 4225–4230. [Google Scholar] [CrossRef] [Green Version]
- Ochoa, W.F.; Havens, W.M.; Sinkovits, R.S.; Nibert, M.L.; Ghabrial, S.A.; Baker, T.S. Partitivirus structure reveals a 120-subunit, helix-rich capsid with distinctive surface arches formed by quasisymmetric coat-protein dimers. Structure 2008, 16, 776–786. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.Q.; Cheng, J.S.; Tang, J.H.; Fu, Y.P.; Jiang, D.H.; Baker, T.S.; Ghabrial, S.A.; Xie, J.T. A Novel partitivirus that confers hypovirulence on plant pathogenic fungi. J. Virol. 2014, 88, 10120–10133. [Google Scholar] [CrossRef] [Green Version]
- Vainio, E.J.; Chiba, S.; Ghabrial, S.A.; Maiss, E.; Roossinck, M.; Sabanadzovic, S.; Suzuki, N.; Xie, J.; Nibert, M. ICTV virus taxonomy profile: Partitiviridae. J. Gen. Virol. 2018, 99, 17–18. [Google Scholar] [CrossRef]
- Gilbert, K.B.; Holcomb, E.E.; Allscheid, R.L.; Carrington, J.C. Hiding in plain sight: New virus genomes discovered via a systematic analysis of fungal public transcriptomes. PLoS ONE 2019, 14, e0219207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nibert, M.L.; Ghabrial, S.A.; Maiss, E.; Lesker, T.; Vainio, E.J.; Jiang, D.; Suzuki, N. Taxonomic reorganization of family Partitiviridae and other recent progress in partitivirus research. Virus Res. 2014, 188, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Jian, B.; Wang, N.; Sun, Y.; de la Llera Moya, M.; Phillips, M.C.; Rothblat, G.H.; Swaney, J.B.; Tall, A.R. Scavenger receptor BI promotes high density lipoprotein-mediated cellular cholesterol efflux. J. Biol. Chem. 1997, 272, 20982–20985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitt, M.J.; Breinig, F. Yeast viral killer toxins: Lethality and self-protection. Nat. Rev. Microbiol. 2006, 4, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Potgieter, C.A.; Castillo, A.; Castro, M.; Cottet, L.; Morales, A. A wild-type Botrytis cinerea strain co-infected by double-stranded RNA mycoviruses presents hypovirulence-associated traits. Virol. J. 2013, 10, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrero, N.; Dueñas, E.; Quesada-Moraga, E.; Zabalgogeazcoa, I. Prevalence and diversity of viruses in the entomopathogenic fungus Beauveria bassiana. Appl. Environ. Microbiol. 2012, 78, 8523–8530. [Google Scholar] [CrossRef] [Green Version]
- Morris, T.; Dodds, J. Isolation and analysis of double-stranded RNA from virus-infected plant and fungal tissue. Phytopathology 1979, 69, 854–858. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coutts, R.H.; Livieratos, I. A rapid method for sequencing the 5′-and 3′-termini of double-stranded RNA viral templates using RLM-RACE. J. Phytopathol. 2003, 151, 525–527. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.-H.; Cheng, Z.; Yang, X.-L.; Li, S.; Ding, Z.-Q.; Zhou, T.-S.; Zhang, W.-J.; Chen, J.-K. Genetic diversity and structure of Cordyceps sinensis populations from extensive geographical regions in China as revealed by inter-simple sequence repeat markers. J. Microbiol. 2008, 46, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, J.; Hu, Y.; Ying, S.-H.; Feng, M.-G. Roles of six Hsp70 genes in virulence, cell wall integrity, antioxidant activity and multiple stress tolerance of Beauveria bassiana. Fungal Genet. Biol. 2020, 144, 103437. [Google Scholar] [CrossRef]
- Meng, H.; Wang, Z.; Wang, Y.; Zhu, H.; Huang, B. Dicer and Argonaute genes involved in RNA interference in the entomopathogenic fungus Metarhizium robertsii. Appl. Environ. Microbiol. 2017, 83, e03230-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.-X.; Zhou, X.-Z.; Meng, H.-M.; Liu, Y.-J.; Zhou, Q.; Huang, B. Comparative transcriptomic analysis of the heat stress response in the filamentous fungus Metarhizium anisopliae using RNA-Seq. Appl. Microbiol. Biot. 2014, 98, 5589–5597. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Shrestha, B.; Sung, G.-H.; Han, S.-K.; Sung, J.-M. Optimum conditions for artificial fruiting body formation of Cordyceps cardinalis. Mycobiology 2010, 38, 133–136. [Google Scholar] [CrossRef] [Green Version]
- Ying, S.-H.; Feng, M.-G. A conidial protein (CP15) of Beauveria bassiana contributes to the conidial tolerance of the entomopathogenic fungus to thermal and oxidative stresses. Appl. Microbiol. Biot. 2011, 90, 1711–1720. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.-L.; Ying, S.-H.; Feng, M.-G.; Hatting, J.L. In vitro and in vivo responses of fungal biocontrol agents to gradient doses of UV-B and UV-A irradiation. Biocontrol 2010, 55, 413–422. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, N.; Wang, P.; Zhu, Q.; Yang, G.; Huang, B. Molecular characterization of a novel alternavirus infecting the entomopathogenic fungus Cordyceps chanhua. Arch. Virol. 2022, 167, 1467–1470. [Google Scholar] [CrossRef]
- Cornejo, C.; Hisano, S.; Bragança, H.; Suzuki, N.; Rigling, D. A new double-stranded RNA mycovirus in Cryphonectria naterciae is able to cross the species barrier and is deleterious to a new host. J. Fungi 2021, 7, 861. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Q.; Shi, N.; Wang, P.; Zhang, Y.; Peng, F.; Yang, G.; Huang, B. A Novel Gammapartitivirus That Causes Changes in Fungal Development and Multi-Stress Tolerance to Important Medicinal Fungus Cordyceps chanhua. J. Fungi 2022, 8, 1309. https://doi.org/10.3390/jof8121309
Zhu Q, Shi N, Wang P, Zhang Y, Peng F, Yang G, Huang B. A Novel Gammapartitivirus That Causes Changes in Fungal Development and Multi-Stress Tolerance to Important Medicinal Fungus Cordyceps chanhua. Journal of Fungi. 2022; 8(12):1309. https://doi.org/10.3390/jof8121309
Chicago/Turabian StyleZhu, Qiuyan, Najie Shi, Ping Wang, Yuxiang Zhang, Fan Peng, Guogen Yang, and Bo Huang. 2022. "A Novel Gammapartitivirus That Causes Changes in Fungal Development and Multi-Stress Tolerance to Important Medicinal Fungus Cordyceps chanhua" Journal of Fungi 8, no. 12: 1309. https://doi.org/10.3390/jof8121309