
Diversity of Mycoviruses Present in Strains of Binucleate Rhizoctonia and Multinucleate Rhizoctonia, Causal Agents for Potato Stem Canker or Black Scurf

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains
2.2. Extraction of Total RNA
2.3. Metatranscriptome Sequencing
2.4. Genome Organization and Phylogenetic Analysis
2.5. Virus Names
3. Results
3.1. Comparison of Mycoviral Diversity in BNR and MNR
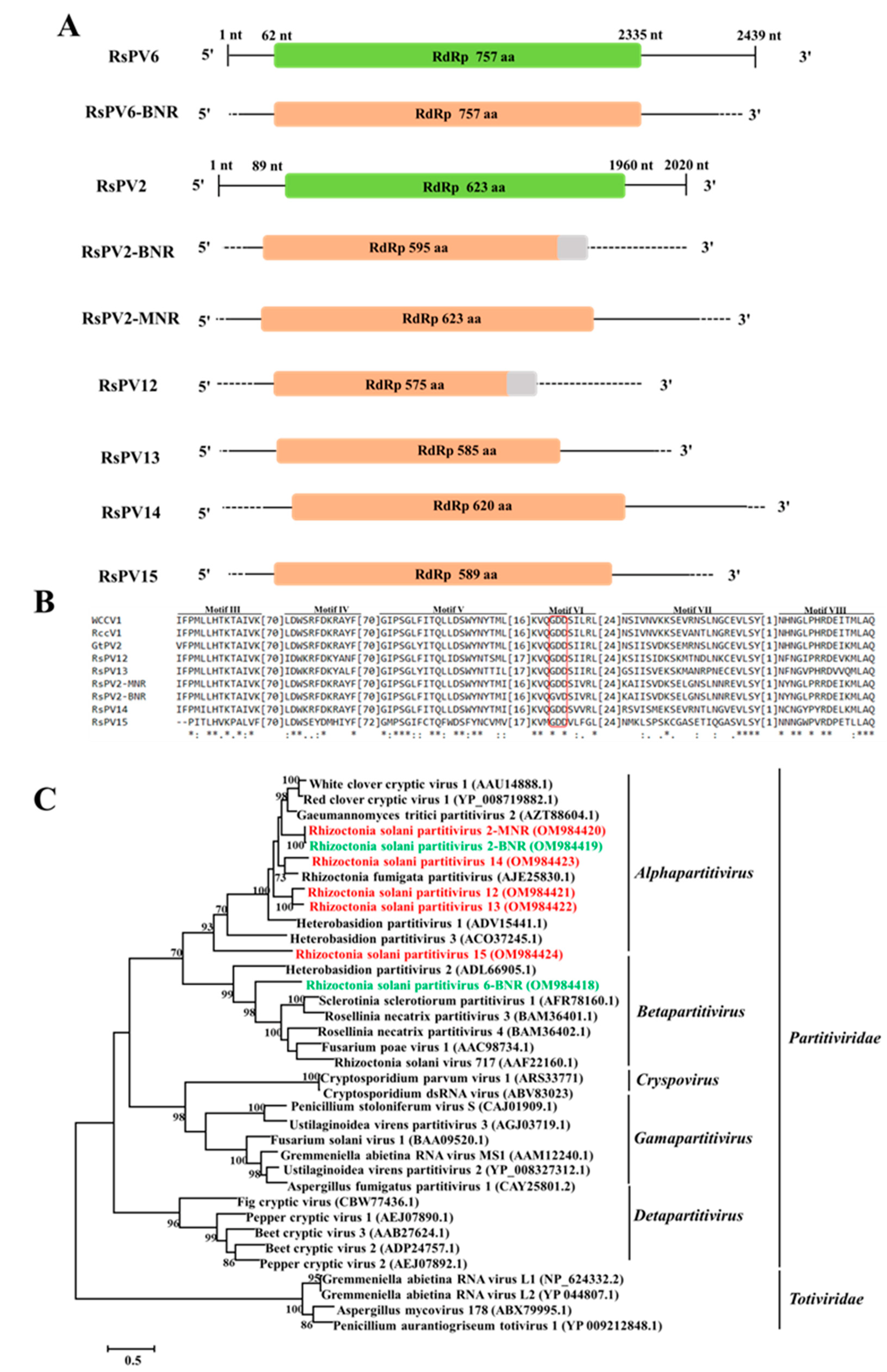
3.2. Genome Organization and Phylogenetic Analysis of Putative Members of the Family Partitiviridae
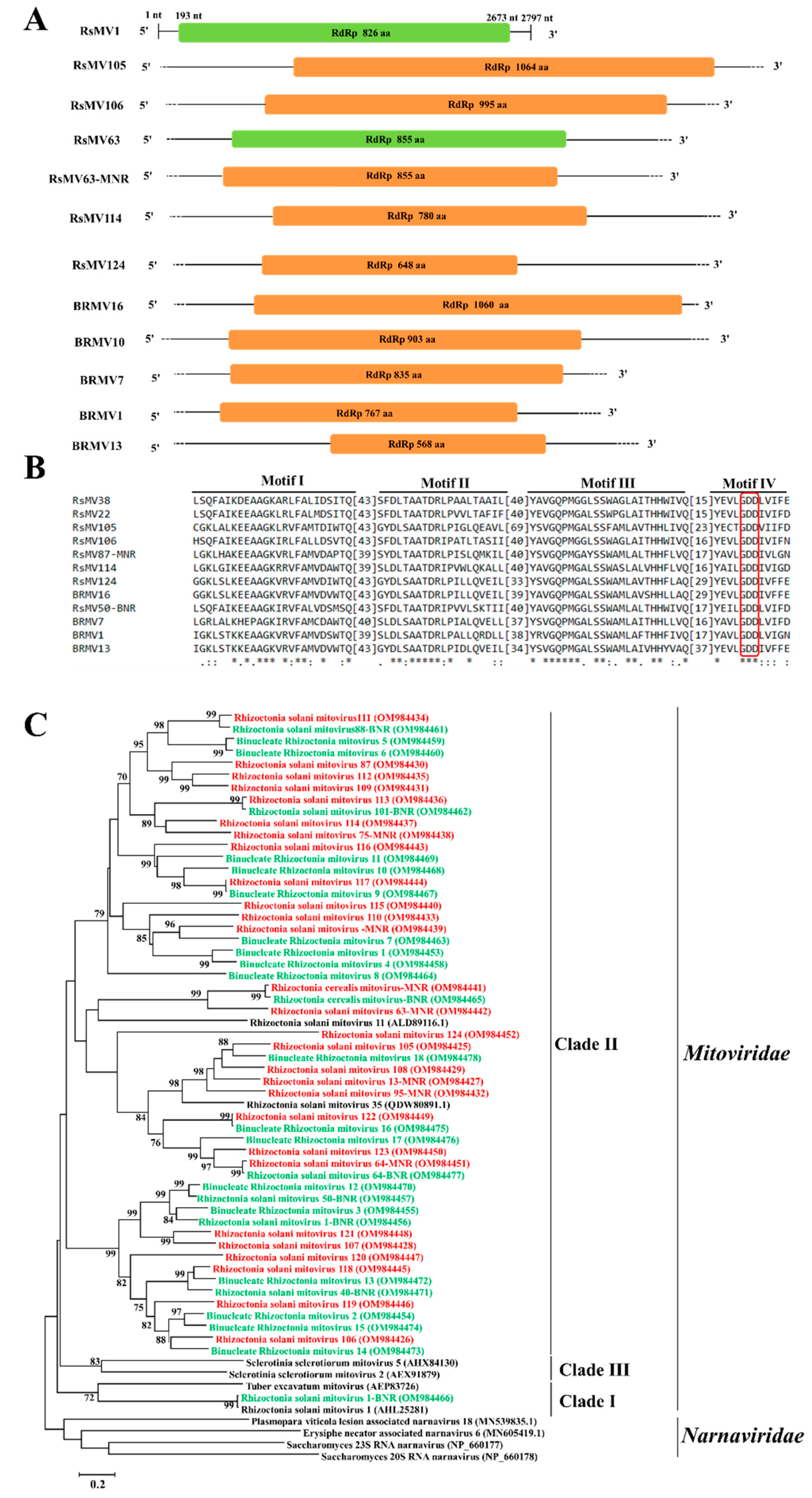
3.3. Genome Organization and Phylogenetic Analysis of Putative Members of the Family Mitoviridae
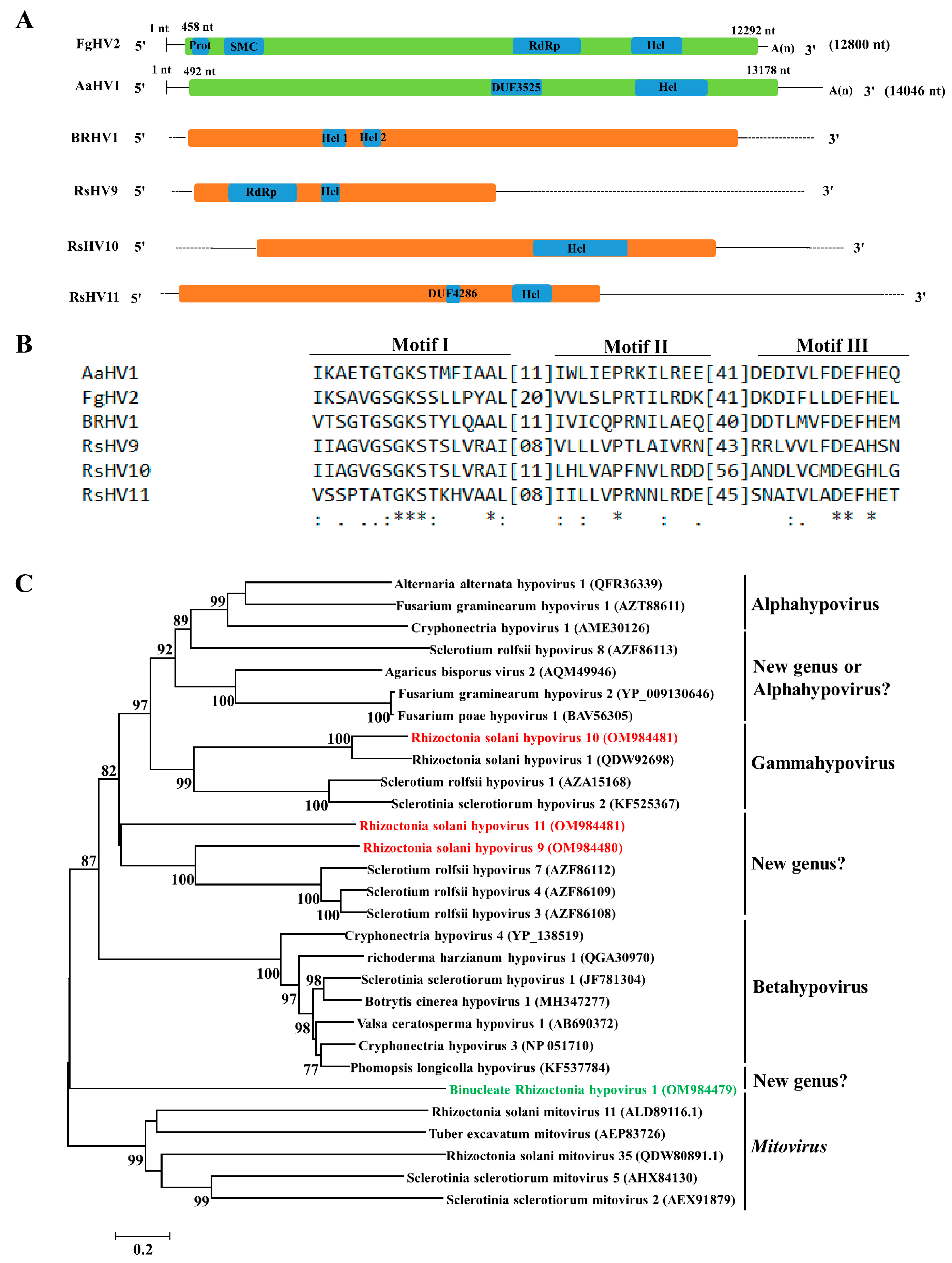
3.4. Genome Organization and Phylogenetic Analysis of Putative Members of the Family Hypoviridae
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gandy, D.G.A. Transmissible disease of cultivated mushrooms (‘watery stipe’). Ann. Appl. Biol. 1960, 48, 427–430. [Google Scholar] [CrossRef]
- Ghabrial, S.A.; Suzuki, N. Viruses of plant pathogenic fungi. Annu. Rev. Phytopathol. 2009, 47, 353–384. [Google Scholar] [CrossRef] [PubMed]
- Ghabrial, S.A.; Caston, J.R.; Jiang, D.H.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Adboulaye, A.H.; Foda, M.F.; Kotta-Loizou, I. Viruses infecting the plant pathogenic fungus Rhizoctonia solani. Viruses 2019, 11, 1113–1137. [Google Scholar]
- Xie, J.T.; Jiang, D.H. New insights into mycoviruses and exploration for the biological control of crop fungal diseases. Annu. Rev. Phytopathol. 2014, 52, 45–68. [Google Scholar] [CrossRef]
- Kotta-Loizou, I. Mycoviruses and their role in fungal pathogenesis. Curr. Opin. Microbiol. 2021, 63, 10–18. [Google Scholar] [CrossRef]
- Tran, T.T.; Li, H.; Nguyen, D.Q.; Jones, M.G.K.; Wylie, S.T. Co-infection with three mycoviruses stimulates growth of a Monilinia fructicola isolate on nutrient medium, but does not induce hypervirulence in a natural host. Viruses 2019, 11, 89. [Google Scholar] [CrossRef]
- Cai, G.H.; Hillman, B.I. Phytophthora viruses. Adv. Virus Res. 2013, 86, 327–350. [Google Scholar]
- Olivé, M.; Campo, S. The dsRNA mycovirus ChNRV1 causes mild hypervirulence in the fungal phytopathogen Colletotrichum higginsianum. Arch. Microbiol. 2021, 203, 241–249. [Google Scholar] [CrossRef]
- Okada, R.; Ichinose, S.; Takeshita, K.; Urayama, S.; Fukuhara, T.; Komatsu, K.; Arie, T.; Ishihara, A.; Egusa, M.; Kodama, M.; et al. Molecular characterization of a novel mycovirus in Alternaria alternata manifesting two-sided effects: Down-regulation of host growth and up-regulation of host plant pathogenicity. Virology 2018, 519, 23–32. [Google Scholar] [CrossRef]
- Sutela, S.; Poimala, A.; Vainio, E.J. Viruses of fungi and oomycetes in the soil environment. FEMS Microbiol. Ecol. 2019, 95, fiz119. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Kim, K.H. Exploration of the interactions between mycoviruses and Fusarium graminearum. Adv. Virus Res. 2010, 106, 123–144. [Google Scholar]
- Milgroom, M.G.; Cortesi, P. Biological control of chestnut blight with hypovirulence: A critical analysis. Annu. Rev. Phytopathol. 2004, 42, 311–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.X.; Xie, J.T.; Fu, Y.P.; Cheng, J.S.; Qu, Z.; Zhao, Z.Z.; Cheng, S.F.; Chen, T.; Li, B.; Wang, Q.Q.; et al. A 2-kb mycovirus converts a pathogenic fungus into a beneficial endophyte for Brassica protection and yield enhancement. Mol. Plant 2020, 13, 1420–1433. [Google Scholar] [CrossRef]
- Abdoulaye, A.H.; Cheng, J.S.; Fu, Y.P.; Jiang, D.H.; Xie, J.T. Complete genome sequence of a novel mitovirus from the phytopathogenic fungus Rhizoctonia oryzae-sativae. Arch. Virol. 2017, 162, 1409–1412. [Google Scholar] [CrossRef]
- Bartholomäus, A.; Wibberg, D.; Winkler, A.; Puhler, A.; Schluter, A.; Varrelmann, M. Deep sequencing analysis reveals the mycoviral diversity of the virome of an avirulent isolate of Rhizoctonia solani AG-2-2 IV. PLoS ONE 2016, 11, e0165965. [Google Scholar] [CrossRef]
- Andika, I.B.; Wei, S.; Cao, C.M.; Salaipeth, L.; Kondo, H.; Sun, L.Y. Phytopathogenic fungus hosts a plant virus: A naturally occurring cross-kingdom viral infection. Proc. Natl. Acad. Sci. USA 2017, 114, 12267–12272. [Google Scholar] [CrossRef]
- Das, S.; Falloon, R.E.; Stewart, A.; Pitman, A.R. Novel mitoviruses in Rhizoctonia solani AG-3PT infecting potato. Fungal Biol. 2016, 120, 338–350. [Google Scholar] [CrossRef]
- Li, Y.T.; Li, S.W.; Liang, Z.J.; Cai, Q.N.; Zhou, T.; Zhao, C.; Wu, X.H. RNA-seq analysis of Rhizoctonia solani AG-4HGI strain BJ-1H infected by a new viral strain of Rhizoctonia solani partitivirus 2 reveals a potential mechanism for hypovirulence. Phytopathology 2022, 112, 1373–1385. [Google Scholar] [CrossRef]
- Li, Y.T.; Li, S.W.; Zhao, Y.M.; Zhou, T.; Wu, X.H.; Zhao, C. Six novel mycoviruses containing positive single-stranded RNA and double-stranded RNA genomes co-infect a single strain of the Rhizoctonia solani AG-3 PT. Viruses 2022, 14, 813. [Google Scholar] [CrossRef]
- Li, P.F.; Zhang, H.L.; Chen, X.G.; Qiu, D.W.; Guo, L.H. Molecular characterization of a novel hypovirus from the plant pathogenic fungus Fusarium graminearum. Virology 2015, 481, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Li, W.; Chen, H.G.; Yu, H.S. Full genome sequence of a putative novel mitovirus isolated from Rhizoctonia cerealis. Arch. Virol. 2015, 160, 1815–1818. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, T.; Sun, H.Y.; Deng, Y.Y.; Zhang, A.X.; Chen, H.G.; Wang, K.R. Complete genome sequence of a novel endornavirus in the wheat sharp eyespot pathogen Rhizoctonia cerealis. Arch. Virol. 2014, 159, 1213–1216. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Q.; Sun, Y.; Xu, P.; Zhang, L.F.; Chen, W.R.; Ren, Z.; Yang, G.H.; Mo, X.H. Complete nucleotide sequence of a novel mycovirus infecting Rhizoctonia fumigata AG-Ba isolate C-314 Baishi. Arch. Virol. 2022, 167, 959–963. [Google Scholar] [CrossRef]
- Li, Y.Q.; Sun, Y.; Xu, P.; Zhang, L.F.; Chen, Z.B.; Niu, Y.F.; Yang, G.H.; Mo, X.H. A novel alphapartitivirus from binucleate Rhizoctonia fumigata AG-Ba isolate C-314 Baishi. Arch. Virol. 2022, 167, 255–259. [Google Scholar] [CrossRef]
- Neang, S.; Bincader, S.; Rangsuwan, S.; Keawmanee, P.; Rin, S.; Salaipeth, L.; Das, S.; Kondo, H.; Suzuki, N.; Sato, I.; et al. Omnipresence of partitiviruses in rice aggregate sheath spot symptom-associated fungal isolates from paddies in Thailand. Viruses 2021, 13, 2269. [Google Scholar] [CrossRef]
- Roossinck, M.J. Life styles of plant viruses. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 1899–1905. [Google Scholar] [CrossRef]
- Roossinck, M.J. Plant virus metagenomics: Biodiversity and ecology. Annu. Rev. Genet. 2012, 46, 357–367. [Google Scholar] [CrossRef]
- Roossinck, M.J. Metagenomics of plant and fungal viruses reveals an abundance of persistent lifestyles. Front. Microbiol. 2015, 5, 767. [Google Scholar] [CrossRef]
- Feldman, T.S.; Morsy, M.R.; Roossinck, M.J. Are communities of microbial symbionts more diverse than communities of microbial hosts? Fungal Biol. 2012, 116, 465–477. [Google Scholar] [CrossRef]
- Marzano, S.Y.L.; Nelson, B.D.; Ajayi-Oyetunde, O.; Bradley, C.A.; Hughes, T.J.; Hartman, G.L.; Eastburn, D.M.; Domier, L.L. Identification of diverse mycoviruses through metatranscriptomics characterization of the viromes of five major fungal plant pathogens. J. Virol. 2016, 90, 6846–6863. [Google Scholar] [CrossRef] [PubMed]
- Picarelli, M.A.S.; Forgia, M.; Rivas, E.B.; Nerva, L.; Chiapello, M.; Turina, M.; Colariccio, A. Extreme diversity of mycoviruses present in isolates of Rhizoctonia solani AG2-2 LP from Zoysia japonica from Brazil. Front. Cell Infect. Microbiol. 2019, 9, 224. [Google Scholar] [CrossRef]
- He, Z.R.; Huang, X.T.; Fan, Y.; Yang, M.; Zhou, E.X. Metatranscriptomic analysis reveals rich mycoviral diversity in three major fungal pathogens of rice. Int. J. Mol. Sci. 2022, 23, 9192. [Google Scholar] [CrossRef] [PubMed]
- Banville, G.J. Yield losses and damage to potato plants caused by Rhizoctonia solani Kühn. Am. Potato J. 1989, 66, 821–834. [Google Scholar] [CrossRef]
- Balali, G.R.; Neate, S.M.; Scott, E.S.; Whisson, D.L.; Wicks, T.J. Anastomosis group and pathogenicity of isolates of Rhizoctonia solani from potato crops in South Australia. Plant Pathol. 1995, 44, 1050–1057. [Google Scholar] [CrossRef]
- Woodhall, J.W.; Lee, A.K.; Edwards, S.G.; Jenkinson, P. Infection of potato by Rhizoctonia solani: Effect of anastomosis group. Plant Pathol. 2008, 57, 897–905. [Google Scholar] [CrossRef]
- Fiers, M.; Edel-Hermann, V.; Héraud, C.; Gautheron, N.; Chatot, C.; Le-Hingrat, Y.; Bouchek-Mechiche, K.; Steinberg, C. Genetic diversity of Rhizoctonia solani associated with potato tubers in France. Mycologia 2011, 103, 1230–1244. [Google Scholar] [CrossRef]
- Woodhall, J.W.; Adams, I.P.; Peters, J.C.; Harper, G.; Boonham, N. A new quantitative real-time PCR assay for Rhizoctonia solani AG3-PT and the detection of AGs of Rhizoctonia solani associated with potato in soil and tuber samples in Great Britain. Eur. J. Plant Pathol. 2013, 136, 273–280. [Google Scholar] [CrossRef]
- Yang, Y.G.; Zhao, C.; Guo, Z.J.; Wu, X.H. Anastomosis group and pathogenictity of Rhizoctonia solani associated with stem canker and black scurf of potato in China. Eur. J. Plant Pathol. 2015, 143, 99–111. [Google Scholar] [CrossRef]
- Gush, S.; Muzhinji, N.; Turter, M.; van der Waals, J.E. First report of Rhizoctonia solani AG-2-2IIIB causing elephant hide on potato tubers in South Africa. Plant Dis. 2019, 103, 1776. [Google Scholar] [CrossRef]
- Murdock, M.R.; Woodhall, J.W.; Maggard, R.; Keith, S.; Harrington, M.; Oropeza, A.; Stokes, B.; Duellman, K.M. First report of Rhizoctonia solani AG 2-1 causing stem canker of potato (Solanum tuberosum) in Idaho. Plant Dis. 2019, 103, 2130. [Google Scholar] [CrossRef]
- Yang, S.; Kong, Y.; Min, F.; Zhang, J.; Wang, L.; Hu, L.; Wang, X. First report of Rhizoctonia solani AG-1-IB infecting potato with stem canker in Heilongjiang province, China. Plant Dis. 2019, 103, 2126. [Google Scholar] [CrossRef]
- Carling, D.E.; Brainard, K.A.; Virgen-Calleros, G.; Olalde-Portugal, V. First report of Rhizoctonia solani AG-7 on potato in Mexico. Plant Dis. 1998, 82, 127. [Google Scholar] [CrossRef] [PubMed]
- Truter, M.; Wehner, F.C. Anastomosis grouping of Rhizoctonia solani associated with black scurf and stem canker of potato in South Africa. Plant Dis. 2004, 88, 83. [Google Scholar] [CrossRef] [PubMed]
- Yanar, Y.; Yilmaz, G.; Cesmeli, I.; Coskun, S. Characterization of Rhizoctonia solani isolates from potatoes in Turkey and screening potato cultivars for resistance to AG-3 isolates. Phytoparasitica 2005, 33, 370–376. [Google Scholar] [CrossRef]
- Harris, A.R. Solid formulations of binucleate Rhizoctonia isolates suppress Rhizoctonia solani and Pythium ultimum in potting medium. Microbiol. Res. 2000, 154, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Herr, L.J. Relationship of binucleate Rhizoctonia isolates used for biocontrol of Rhizoctonia crown rot of sugar beet to anastomosis systems. Can. J. Microbiol. 1991, 37, 339–344. [Google Scholar] [CrossRef]
- Yang, Y.G.; Zhao, C.; Guo, Z.J.; Wu, X.H. Anastomosis groups and pathogenicity of binucleate Rhizoctonia isolates associated with stem canker of potato in China. Eur. J. Plant Pathol. 2014, 139, 535–544. [Google Scholar] [CrossRef]
- Yang, Y.G.; Wu, X.H. First report of potato stem canker caused by binucleate Rhizoctonia AG-A in Jilin province, China. Plant Dis. 2013, 97, 1246. [Google Scholar] [CrossRef]
- Muzhinji, N.; Truter, M.; Woodhall, J.W.; van der Waals, J.E. Anastomosis group and pathogenicity of Rhizoctonia solani and binucleate Rhizoctonia from potato in South Africa. Plant Dis. 2015, 99, 1790–1802. [Google Scholar] [CrossRef]
- Escande, A.R.; Echandi, E. Protection of potato from Rhizoctonia canker with binucleate Rhizoctonia fungi. Plant Pathol. 1991, 40, 197–202. [Google Scholar] [CrossRef]
- Yang, Y.G.; Zhao, C.; Guo, Z.J.; Wu, X.H. Potato stem canker caused by binucleate Rhizoctonia AG-G in China. J. Gen. Plant Pathol. 2015, 81, 287–290. [Google Scholar] [CrossRef]
- Yang, Y.G.; Zhao, C.; Guo, Z.J.; Wu, X.H. Characterization of a new anastomosis group (AG-W) of binucleate Rhizoctonia, causal agent for potato stem canker. Plant Dis. 2015, 99, 1757–1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Zheng, L.; Zhang, M.L.; Chen, Q.G.; Zhu, M.H.; Zhou, E.X. A novel mycovirus closely related to viruses in the genus Alphapartitivirus confers hypovirulence in the phytopathogenic fungus Rhizoctonia solani. Virology 2014, 456, 220–226. [Google Scholar] [CrossRef]
- Chiapello, M.; Rodriguez-Romero, J.; Ayllon, M.A.; Turina, M. Analysis of the virome associated to grapevine downy mildew lesions reveals new mycovirus lineages. Virus Evol. 2020, 6, veaa058. [Google Scholar] [CrossRef]
- Chen, Y.; Gai, X.T.; Chen, R.X.; Li, C.X.; Zhao, G.K.; Xia, Z.Y.; Zou, C.M.; Zhong, J. Characterization of three novel betapartitiviruses co-infecting the phytopathogenic fungus Rhizoctonia solani. Virus Res. 2019, 270, 197649. [Google Scholar] [CrossRef]
- Ong, J.W.L.; Li, H.; Sivasithamparam, K.; Dixon, K.W.; Jones, M.G.K.; Wylie, S.J. The challenges of using high-throughput sequencing to track multiple bipartite mycoviruses of wild orchid-fungus partnerships over consecutive years. Virology 2017, 510, 297–304. [Google Scholar] [CrossRef]
- Lyu, R.L.; Zhang, Y.; Tang, Q.; Li, Y.Y.; Cheng, J.S.; Fu, Y.P.; Chen, T.; Jiang, D.H.; Xie, J.T. Two alphapartitiviruses co-infecting a single isolate of the plant pathogenic fungus Rhizoctonia solani. Arch Virol. 2018, 163, 515–520. [Google Scholar] [CrossRef]
- Shi, M.; Lin, X.D.; Tian, J.H.; Chen, L.J.; Chen, X.; Li, C.X.; Qin, X.C.; Li, J.; Cao, J.P.; Eden, J.S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Bian, R.L.; Liu, Q.; Yang, L.; Pang, T.X.; Salaipeth, L.; Andika, I.B.; Kondo, H.; Sun, L.Y. Identification of a novel hypovirulence-inducing hypovirus from Alternaria alternata. Front. Microbiol. 2019, 10, 1076. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.Z.; Zhu, H.J.; Gao, B.D.; Zhou, Q.; Zhong, J. Diverse, novel mycoviruses from the virome of a hypovirulent Sclerotium rolfsii strain. Front. Plant Sci. 2018, 9, 1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vainio, E.J.; Chiba, S.; Ghabrial, S.A.; Maiss, E.; Roossinck, M.; Sabanadzovic, S.; Suzuki, N.; Xie, J.T.; Nibert, M. ICTV virus taxonomy profile: Partitiviridae. J. Gen. Virol. 2018, 99, 17–18. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Chen, D.; Lei, X.H.; Zhu, H.J.; Zhu, J.Z.; Gao, B.D. Detection and characterization of a novel gammapartitivirus in the phytopathogenic fungus Colletotrichum acutatum strain HNZJ001. Virus Res. 2014, 190, 104–109. [Google Scholar] [CrossRef]
- Jiang, D.H.; Ayllon, M.A.; Marzano, S.Y.L.; Lefkowitz, E.J.; Davison, A.J.; Siddell, S.G.; Sabanadzovic, S.; Simmonds, P.; Smith, D.B.; Orton, R.J. ICTV virus taxonomy profile: Mymonaviridae. J. Gen. Virol. 2019, 100, 1343–1344. [Google Scholar] [CrossRef]
- Zoll, J.; Verweij, P.E.; Melchers, W.J.G. Discovery and characterization of novel Aspergillus fumigatus mycoviruses. PLoS ONE 2018, 13, e0200511. [Google Scholar] [CrossRef]
- Xavier, A.D.; de-Barros, A.P.O.; Godinho, M.T.; Zerbini, F.M.; Souza, F.D.; Bruckner, F.P.; Alfenas-Zerbini, P. A novel mycovirus associated to Alternaria alternata comprises a distinct lineage in Partitiviridae. Virus Res. 2018, 244, 21–26. [Google Scholar] [CrossRef]
- Gilbert, K.B.; Holcomb, E.E.; Allscheid, R.L.; Carrington, J.C. Hiding in plain sight: New virus genomes discovered via a systematic analysis of fungal public transcriptomes. PLoS ONE 2019, 14, e0219207. [Google Scholar] [CrossRef]
- Li, S.W.; Li, Y.T.; Hu, C.H.; Han, C.G.; Zhou, T.; Zhao, C.; Wu, X.H. Full genome sequence of a new mitovirus from the phytopathogenic fungus Rhizoctonia solani. Arch. Virol. 2020, 165, 1719–1723. [Google Scholar] [CrossRef]
- Mizutani, Y.; Uesaka, K.; Ota, A.; Calassanzio, M.; Ratti, C.; Suzuki, T.; Fujimori, F.; Chiba, S. De novo sequencing of novel mycoviruses from Fusarium sambucinum: An attempt on direct RNA sequencing of viral dsRNAs. Front. Microbiol. 2021, 12, 641484. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.Y.; Wu, S.S.; Liu, L.J.; Cheng, J.S.; Fu, Y.P.; Jiang, D.H.; Xie, J.T. A mitovirus related to plant mitochondrial gene confers hypovirulence on the phytopathogenic fungus Sclerotinia sclerotiorum. Virus Res. 2015, 197, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.J.; Wu, S.S.; Cheng, J.S.; Fu, Y.P.; Jiang, D.H.; Xie, J.T. Molecular characterization of two positive-strand RNA viruses co-infecting a hypovirulent strain of Sclerotinia sclerotiorum. Virology 2014, 464, 450–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalifa, M.E.; Pearson, M.N. Characterisation of a novel hypovirus from Sclerotinia sclerotiorum potentially representing a new genus within the Hypoviridae. Virology 2014, 464, 441–449. [Google Scholar] [CrossRef]
- Osaki, H.; Sasaki, A.; Nomiyama, K.; Tomioka, K. Multiple virus infection in a single strain of Fusarium poae shown by deep sequencing. Virus Genes 2016, 52, 835–847. [Google Scholar] [CrossRef]
- Nuss, D.L. Hypovirulence: Mycoviruses at the fungal-plant interface. Nat. Rev. Microbiol. 2005, 3, 632–642. [Google Scholar] [CrossRef]
- Hillman, B.I.; Halpern, B.T.; Brown, M.P. A viral dsRNA element of the chestnut blight fungus with a distinct genetic organization. Virology 1994, 201, 241–250. [Google Scholar] [CrossRef]
- Hao, F.M.; Ding, T.; Wu, M.D.; Zhang, J.; Yang, L.; Chen, W.D.; Li, G.Q. Two novel hypovirulence-associated mycoviruses in the phytopathogenic fungus Botrytis cinerea: Molecular characterization and suppression of infection cushion formation. Viruses 2018, 10, 254. [Google Scholar] [CrossRef]
- Smart, C.D.; Yuan, W.; Foglia, R.; Nuss, D.L.; Fulbright, D.W.; Hillman, B.I. Cryphonectria hypovirus 3, a virus species in the family Hypoviridae with a single open reading frame. Virology 1999, 265, 66–73. [Google Scholar] [CrossRef]
- Yaegashi, H.; Kanematsu, S.; Ito, T. Molecular characterization of a new hypovirus infecting a phytopathogenic fungus, Valsa ceratosperma. Virus Res. 2012, 165, 143–150. [Google Scholar] [CrossRef]
- Linder-Basso, D.; Dynek, J.N.; Hillman, B.I. Genome analysis of Cryphonectria hypovirus 4, the most common hypovirus species in North America. Virology 2005, 337, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.C.; Kondo, H.; Liu, L.; Guo, L.H.; Qiu, D.W. A novel virus in the family Hypoviridae from the plant pathogenic fungus Fusarium graminearum. Virus Res. 2013, 174, 69–77. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Library Sample | Number of Contigs | Average Length (Base Pair) | Annotation | ||
|---|---|---|---|---|---|
| Host Fungi | Virus | Not Found | |||
| BNR | 99,951 | 473.87 | 82,400 (82.44%) | 225 (0.23%) | 17,326 (17.33%) |
| MNR | 112,939 | 477.17 | 94,251 (83.45%) | 571 (0.51%) | 18,117 (16.04%) |
| Type of Nucleic Acid | Virus Family | Number of Mycovirus Contigs | |
|---|---|---|---|
| BNR | MNR | ||
| Positive single-stranded RNA (+ssRNA) | Benyviridae | 14 | 15 |
| Botourmiaviridae | 8 | 11 | |
| Endornaviridae | 21 | 71 | |
| Fusariviridae | 13 | 67 | |
| Hypoviridae | 5 | 20 | |
| Mitoviridae | 75 | 149 | |
| Narnaviridae | 4 | 10 | |
| Betaflexiviridae | - a | 1 | |
| Deltaflexiviridae | - a | 1 | |
| Gammaflexiviridae | - a | 1 | |
| Tymoviridae | - a | 13 | |
| Togaviridae | - a | 3 | |
| Tombusviridae | - a | 1 | |
| Virgaviridae | 1 | - b | |
| unclassified | 1 | 5 | |
| Double-stranded RNA (dsRNA) | Bipartitiviridae | 11 | 8 |
| Partitiviridae | 10 | 64 | |
| Totiviridae | 3 | 11 | |
| Megabirnaviridae | - a | 2 | |
| Negative single-stranded RNA (-ssRNA) | Mymonaviridae | 4 | 1 |
| Bunyaviridae | - a | 3 | |
| Rhabdoviridae | 1 | - b | |
| unclassified | 2 | 28 | |
| Total | 173 | 485 | |
| Contig | Size (Amino Acid) | Name | Origin | Best Match | Identity | Query Cover | E-Value | Accession Number |
|---|---|---|---|---|---|---|---|---|
| Contig5377 | 757 | Rhizoctonia solani partitivirus 6-BNR | BNR | Rhizoctonia solani partitivirus 6 | 99% | 93% | 0 | OM984418 |
| Contig47566 | 595 | Rhizoctonia solani Partitivirus 2-BNR | BNR | Rhizoctonia solani partitivirus 2 | 99% | 96% | 0 | OM984419 |
| Contig7211 | 623 | Rhizoctonia solani Partitivirus 2-MNR | MNR | Rhizoctonia solani partitivirus 2 | 99% | 95% | 0 | OM984420 |
| Contig6469 a | 575 | Rhizoctonia solani Partitivirus 12 | MNR | Ceratobasidium partitivirus | 56% | 95% | 0 | OM984421 |
| Contig8296 a | 585 | Rhizoctonia solani Partitivirus 13 | MNR | Rhizoctonia solani partitivirus 3 | 53% | 92% | 0 | OM984422 |
| Contig25890 a | 620 | Rhizoctonia solani Partitivirus 14 | MNR | Rhizoctonia fumigata partitivirus | 62% | 93% | 0 | OM984423 |
| Contig59663 a | 589 | Rhizoctonia solani Partitivirus 15 | MNR | Raphanus sativus cryptic virus 1 | 56% | 92% | 0 | OM984424 |
| Contig | Size (Amino Acid) | Name | Origin | Best Match | Identity | Query Cover | E-Value | Accession Number |
|---|---|---|---|---|---|---|---|---|
| First_Contig100 a,b | 767 | Binucleate Rhizoctonia mitovirus 1 | BNR | Rhizoctonia solani mitovirus 15 | 43% | 71% | 0 | OM984453 |
| Contig385 a | 857 | Binucleate Rhizoctonia mitovirus 2 | BNR | Rhizoctonia solani mitovirus 78 | 52% | 67% | 0 | OM984454 |
| Contig462 a | 959 | Binucleate Rhizoctonia mitovirus 3 | BNR | Rhizoctonia solani mitovirus 51 | 72% | 73% | 0 | OM984455 |
| Contig463 | 966 | Rhizoctonia solani mitovirus 51-BNR | BNR | Rhizoctonia solani mitovirus 51 | 97% | 73% | 0 | OM984456 |
| Contig824 | 946 | Rhizoctonia solani mitovirus 50-BNR | BNR | Rhizoctonia solani mitovirus 50 | 96% | 92% | 0 | OM984457 |
| Contig2147 a | 779 | Binucleate Rhizoctonia mitovirus 4 | BNR | Rhizoctonia solani mitovirus 15 | 42% | 72% | 0 | OM984458 |
| First_Contig193 a | 788 | Binucleate Rhizoctonia mitovirus 5 | BNR | Grapevine-associated mitovirus 10 | 71% | 70% | 0 | OM984459 |
| Contig1318 a | 792 | Binucleate Rhizoctonia mitovirus 6 | BNR | Grapevine-associated mitovirus 10 | 71% | 66% | 0 | OM984460 |
| Contig1296 | 833 | Rhizoctonia solani mitovirus 88-BNR | BNR | Rhizoctonia solani mitovirus 88 | 92% | 66% | 0 | OM984461 |
| First_Contig3 | 785 | Rhizoctonia solani mitovirus 101-BNR | BNR | Rhizoctonia solani mitovirus 101 | 98% | 77% | 0 | OM984462 |
| Contig2865 a,b | 835 | Binucleate Rhizoctonia mitovirus 7 | BNR | Rhizoctonia solani mitovirus 15 | 47% | 75% | 0 | OM984463 |
| Contig109 a | 927 | Binucleate Rhizoctonia mitovirus 8 | BNR | Rhizoctonia solani mitovirus 42 | 83% | 74% | 0 | OM984464 |
| Contig377 | 844 | Rhizoctonia cerealis mitovirus-BNR | BNR | Rhizoctonia cerealis mitovirus | 93% | 79% | 0 | OM984465 |
| Contig2511 | 780 | Rhizoctonia solani mitovirus 1-BNR | BNR | Rhizoctonia solani mitovirus 1 | 100% | 92% | 0 | OM984466 |
| Contig139 a | 900 | Binucleate Rhizoctonia mitovirus 9 | BNR | Rhizoctonia solani mitovirus 31 | 61% | 74% | 0 | OM984467 |
| First_Contig8 a,b | 903 | Binucleate Rhizoctonia mitovirus 10 | BNR | Rhizoctonia solani mitovirus 41 | 84% | 74% | 0 | OM984468 |
| Contig461 a | 795 | Binucleate Rhizoctonia mitovirus 11 | BNR | Rhizoctonia solani mitovirus 41 | 56% | 73% | 0 | OM984469 |
| Contig301 a | 811 | Binucleate Rhizoctonia mitovirus 12 | BNR | Rhizoctonia solani mitovirus 50 | 89% | 93% | 0 | OM984470 |
| Second_Contig95 | 842 | Rhizoctonia solani mitovirus 40-BNR | BNR | Rhizoctonia solani mitovirus 40 | 97% | 57% | 0 | OM984471 |
| Contig643 a,b | 568 | Binucleate Rhizoctonia mitovirus 13 | BNR | Epicoccum nigrum mitovirus 1 | 72% | 68% | 0 | OM984472 |
| Contig1166 a | 852 | Binucleate Rhizoctonia mitovirus 14 | BNR | Rhizoctonia solani mitovirus 78 | 62% | 68% | 0 | OM984473 |
| Contig2240 a | 856 | Binucleate Rhizoctonia mitovirus 15 | BNR | Rhizoctonia solani mitovirus 78 | 55% | 65% | 0 | OM984474 |
| Contig333 a,b | 1060 | Binucleate Rhizoctonia mitovirus 16 | BNR | Rhizoctonia solani mitovirus 43 | 73% | 79% | 0 | OM984475 |
| Contig728 a | 1054 | Binucleate Rhizoctonia mitovirus 17 | BNR | Rhizoctonia solani mitovirus 64 | 52% | 73% | 0 | OM984476 |
| Contig2765 | 1047 | Rhizoctonia solani mitovirus 64-BNR | BNR | Rhizoctonia solani mitovirus 64 | 91% | 73% | 0 | OM984477 |
| Contig132 a | 1079 | Binucleate Rhizoctonia mitovirus 18 | BNR | Rhizoctonia solani mitovirus 48 | 53% | 64% | 0 | OM984478 |
| Contig7624 a,b | 1064 | Rhizoctonia solani mitovirus 105 | MNR | Rhizoctonia solani mitovirus 48 | 50% | 67% | 0 | OM984425 |
| Contig787 a,b | 995 | Rhizoctonia solani mitovirus 106 | MNR | Rhizoctonia solani mitovirus 78 | 54% | 60% | 0 | OM984426 |
| Contig722 | 917 | Rhizoctonia solani mitovirus 13-MNR | MNR | Rhizoctonia solani mitovirus 13 | 91% | 67% | 0 | OM984427 |
| Contig944 a | 879 | Rhizoctonia solani mitovirus 107 | MNR | Rhizoctonia solani mitovirus 54 | 64% | 70% | 0 | OM984428 |
| First_Contig217 a | 1027 | Rhizoctonia solani mitovirus 108 | MNR | Rhizoctonia solani mitovirus 48 | 86% | 83% | 0 | OM984429 |
| First_Contig20 | 864 | Rhizoctonia solani mitovirus 87-MNR | MNR | Rhizoctonia solani mitovirus 87 | 97% | 71% | 0 | OM984430 |
| Contig665 a | 865 | Rhizoctonia solani mitovirus 109 | MNR | Rhizoctonia solani mitovirus 65 | 61% | 66% | 0 | OM984431 |
| Contig931 | 1029 | Rhizoctonia solani mitovirus 95-MNR | MNR | Rhizoctonia solani mitovirus 95 | 96% | 54% | 0 | OM984432 |
| Contig201 a | 858 | Rhizoctonia solani mitovirus 110 | MNR | Rhizoctonia solani mitovirus 15 | 46% | 52% | 9 × 10−165 | OM984433 |
| Contig493 a | 836 | Rhizoctonia solani mitovirus 111 | MNR | Rhizoctonia solani mitovirus 88 | 85% | 69% | 0 | OM984434 |
| First_Contig34 a | 862 | Rhizoctonia solani mitovirus 112 | MNR | Rhizoctonia solani mitovirus 65 | 59% | 65% | 0 | OM984435 |
| Contig9722 a | 1028 | Rhizoctonia solani mitovirus 113 | MNR | Macrophomina phaseolina mitovirus 3 | 86% | 74% | 0 | OM984436 |
| First_Contig4017 a,b | 780 | Rhizoctonia solani mitovirus 114 | MNR | Rhizoctonia solani mitovirus 73 | 50% | 51% | 0 | OM984437 |
| First_Contig23 | 782 | Rhizoctonia solani mitovirus 75-MNR | MNR | Rhizoctonia solani mitovirus 75 | 96% | 60% | 0 | OM984438 |
| Contig451 | 961 | Rhizoctonia solani mitovirus 15-MNR | MNR | Rhizoctonia solani mitovirus 15 | 99% | 73% | 0 | OM984439 |
| Contig999 a | 939 | Rhizoctonia solani mitovirus 115 | MNR | Clitocybe odora virus | 39% | 69% | 7 × 10−160 | OM984440 |
| Contig1235 | 810 | Rhizoctonia cerealis mitovirus-MNR | MNR | Rhizoctonia cerealis mitovirus | 92% | 81% | 0 | OM984441 |
| Contig3325 b | 855 | Rhizoctonia solani mitovirus 63-MNR | MNR | Rhizoctonia solani mitovirus 63 | 90% | 78% | 0 | OM984442 |
| Contig2267 a | 841 | Rhizoctonia solani mitovirus 116 | MNR | Rhizoctonia solani mitovirus 31 | 45% | 49% | 2 × 10−175 | OM984443 |
| Contig1401 a | 900 | Rhizoctonia solani mitovirus 117 | MNR | Rhizoctonia solani mitovirus 31 | 60% | 74% | 0 | OM984444 |
| First_Contig722 a | 845 | Rhizoctonia solani mitovirus 118 | MNR | Rhizoctonia solani mitovirus 60 | 81% | 75% | 0 | OM984445 |
| Contig424 a | 843 | Rhizoctonia solani mitovirus 119 | MNR | Rhizoctonia solani mitovirus 78 | 80% | 74% | 0 | OM984446 |
| Contig2296 a | 964 | Rhizoctonia solani mitovirus 120 | MNR | Rhizoctonia solani mitovirus 93 | 64% | 46% | 0 | OM984447 |
| Contig5349 a | 882 | Rhizoctonia solani mitovirus 121 | MNR | Rhizoctonia solani mitovirus 84 | 88% | 72% | 0 | OM984448 |
| Second_Contig262 a | 421 | Rhizoctonia solani mitovirus 122 | MNR | Rhizoctonia solani mitovirus 43 | 74% | 68% | 0 | OM984449 |
| Contig1260 a | 1000 | Rhizoctonia solani mitovirus 123 | MNR | Rhizoctonia solani mitovirus 64 | 61% | 95% | 0 | OM984450 |
| First_Contig63 | 624 | Rhizoctonia solani mitovirus 64-MNR | MNR | Rhizoctonia solani mitovirus 64 | 91% | 91% | 0 | OM984451 |
| Contig2497 a,b | 648 | Rhizoctonia solani mitovirus 124 | MNR | Rhizoctonia solani mitovirus 43 | 69% | 77% | 0 | OM984452 |
| Contig | Size (Amino Acid) | Name | Origin | Best Match | Identity | Query Cover | E-Value | Accession Number |
|---|---|---|---|---|---|---|---|---|
| Contig11643 a | 2650 | Binucleate Rhizoctonia hypovirus 1 | BNR | Sclerotium rolfsii hypovirus 8 | 37% | 67% | 0 | OM984479 |
| Second_Contig451 a | 1776 | Rhizoctonia solani hypovirus 9 | MNR | Lentinula edodes hypovirus 1 | 31% | 62% | 1 × 10−168 | OM984480 |
| First_Contig678 a | 3611 | Rhizoctonia solani hypovirus 10 | MNR | Rhizoctonia solani hypovirus 1 | 50% | 81% | 0 | OM984481 |
| Contig2151 a | 2974 | Rhizoctonia solani hypovirus 11 | MNR | Mycosphaerella hypovirus A | 30% | 59% | 7 × 10−117 | OM984482 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Yang, N.; Mu, T.; Wu, X.; Zhao, C. Diversity of Mycoviruses Present in Strains of Binucleate Rhizoctonia and Multinucleate Rhizoctonia, Causal Agents for Potato Stem Canker or Black Scurf. J. Fungi 2023, 9, 214. https://doi.org/10.3390/jof9020214
Li Y, Yang N, Mu T, Wu X, Zhao C. Diversity of Mycoviruses Present in Strains of Binucleate Rhizoctonia and Multinucleate Rhizoctonia, Causal Agents for Potato Stem Canker or Black Scurf. Journal of Fungi. 2023; 9(2):214. https://doi.org/10.3390/jof9020214
Chicago/Turabian StyleLi, Yuting, Naibo Yang, Tongyu Mu, Xuehong Wu, and Can Zhao. 2023. "Diversity of Mycoviruses Present in Strains of Binucleate Rhizoctonia and Multinucleate Rhizoctonia, Causal Agents for Potato Stem Canker or Black Scurf" Journal of Fungi 9, no. 2: 214. https://doi.org/10.3390/jof9020214