
Arbuscular Mycorrhizal Fungus Alters Alfalfa (Medicago sativa) Defense Enzyme Activities and Volatile Organic Compound Contents in Response to Pea Aphid (Acyrthosiphon pisum) Infestation
Abstract
:1. Introduction
2. Materials and Methods
2.1. Alfalfa, AM Fungus, and Growth Medium
2.2. AM Fungus Inoculation and Aphid Infestation
2.3. AM Colonization, Shoot N, P Uptake, Dry Weight, Defense Enzyme Activity, and Hormone Content
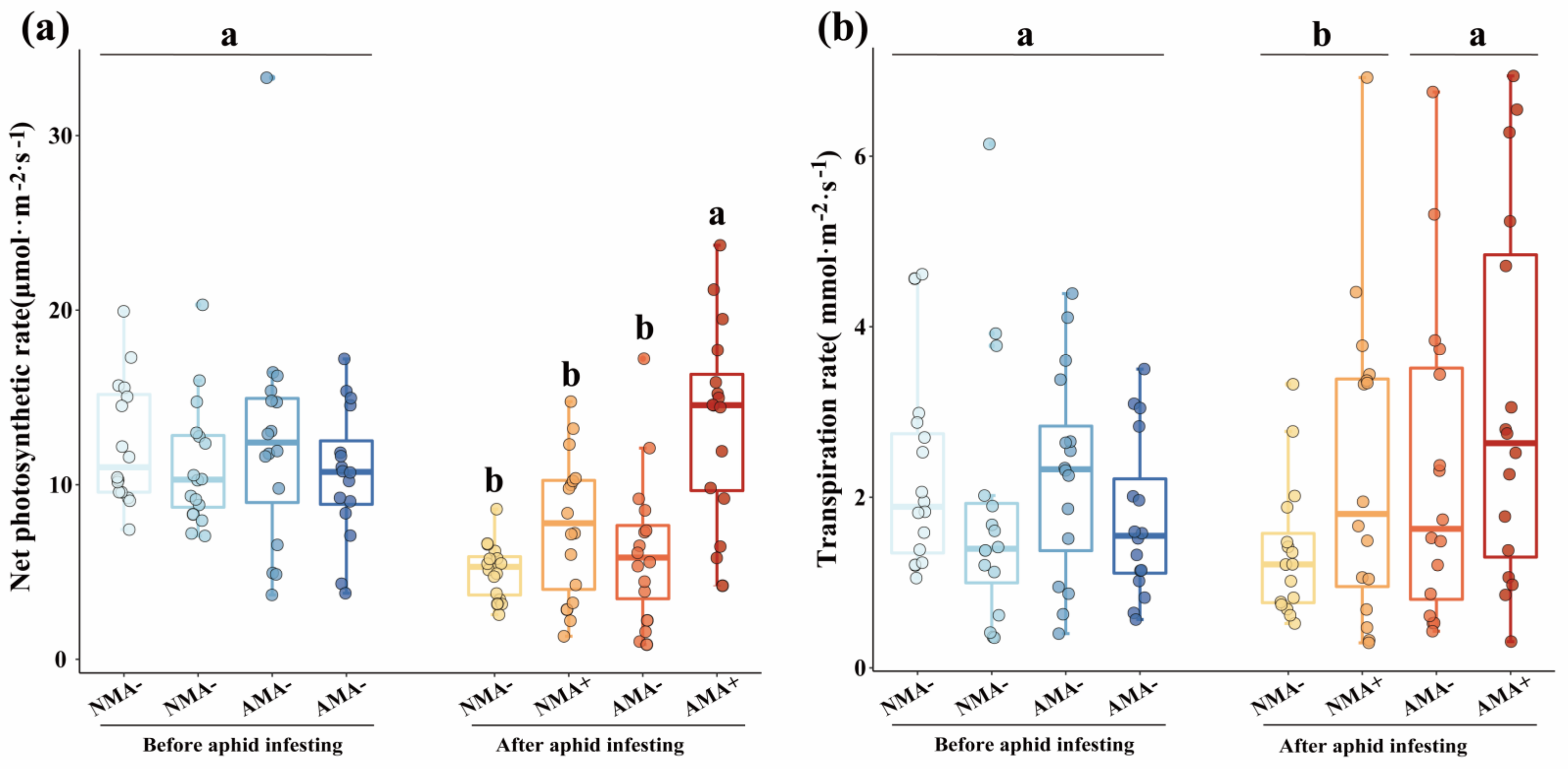
2.4. The Photosynthesis Indexes
2.5. The Transcriptome Analysis for Plant Leaves
2.6. Volatile Organic Components Analysis
2.7. Statistical Analysis
3. Results
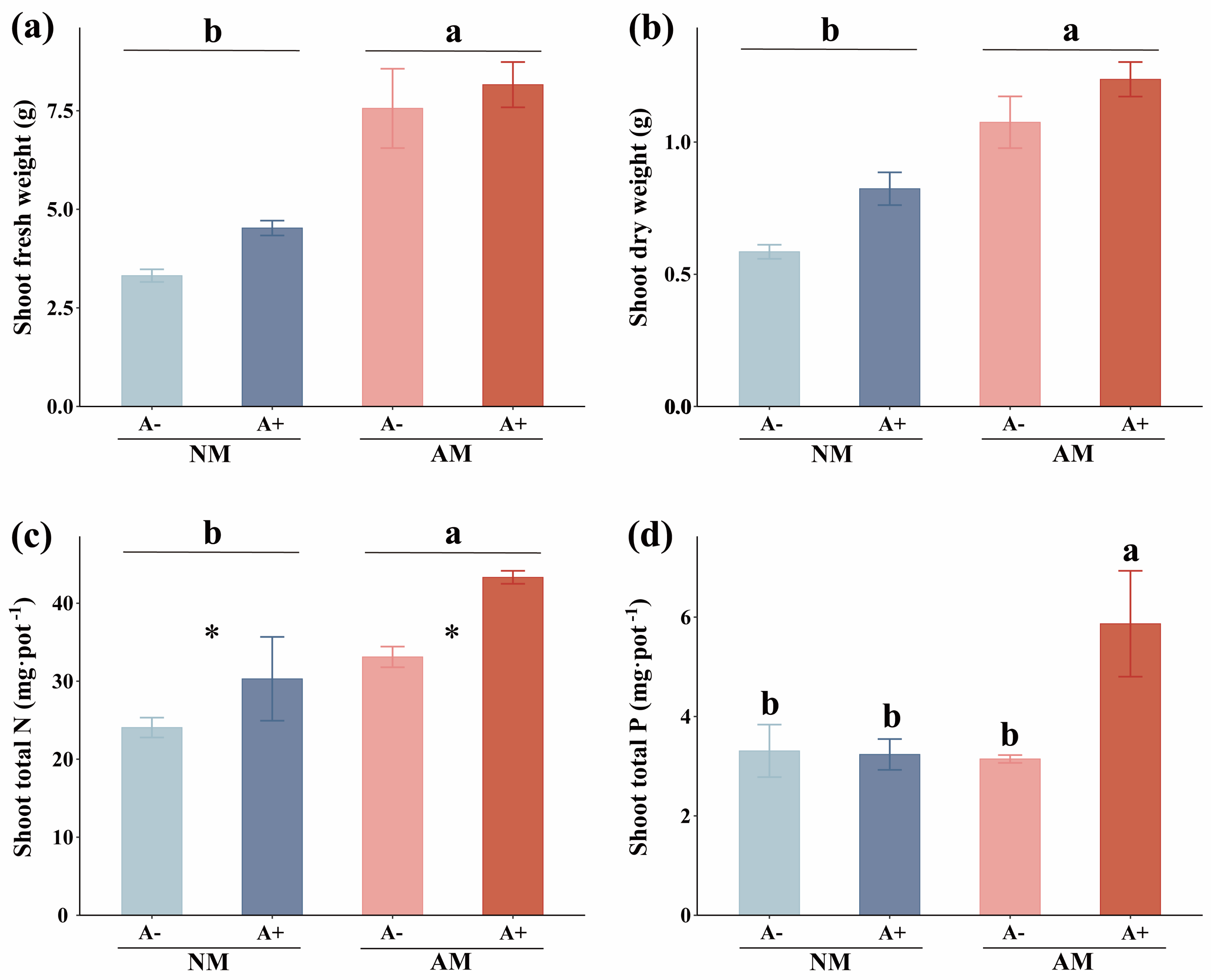
3.1. AM Fungal Colonization, Plant Growth, Shoot N and P Uptake
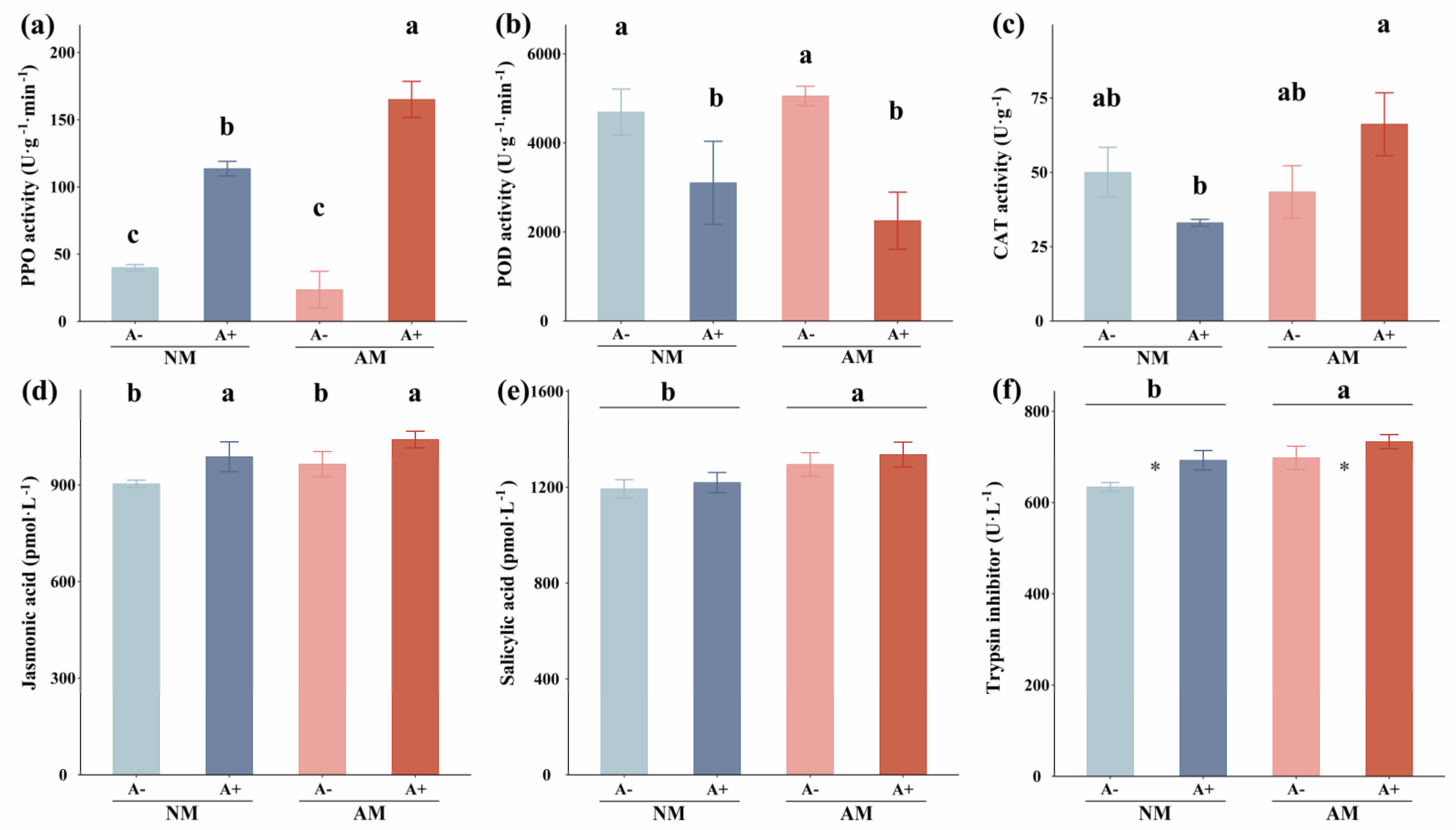
3.2. The Activities of Plant Defense Enzymes, Jasmonic Acid, SA, Trypsin Inhibitors, Total Phenolics, ABA, and NO Concentration in Alfalfa
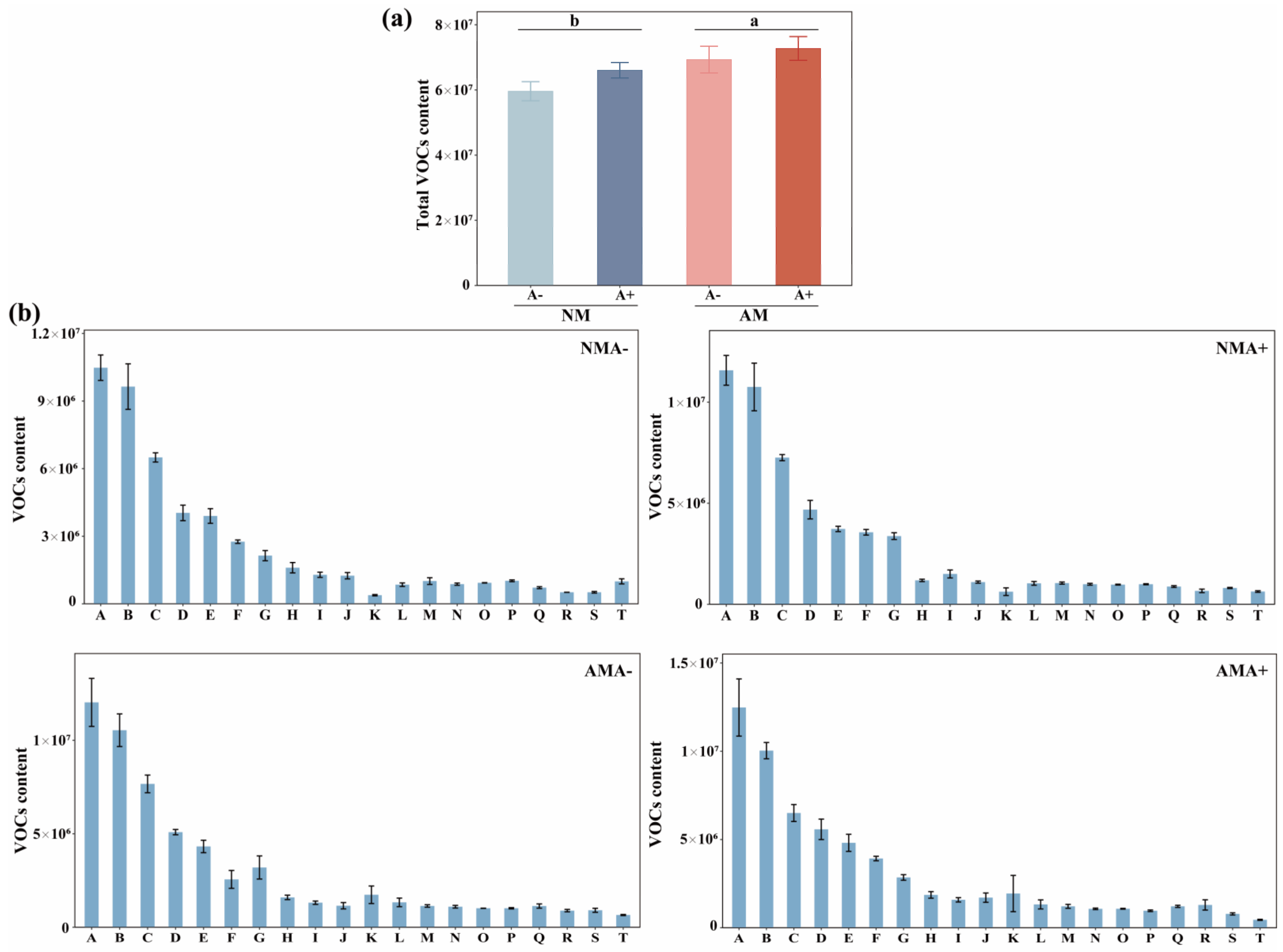
3.3. Volatile Organic Components in Alfalfa
3.4. Differentially Expressed Genes and VOCs Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dedryver, C.A.; Ralec, A.L.; Fabre, F. The conflicting relationships between aphids and men: A review of aphid damage and control strategies. Comptes Rendus Biol. 2010, 333, 539–553. [Google Scholar] [CrossRef] [PubMed]
- Ryalls, J.M.W.; Riegler, M.; Moore, B.D.; Johnson, S.N. Biology and trophic interactions of lucerne aphids. Agric. Meteorol. 2013, 15, 335–350. [Google Scholar] [CrossRef]
- Shi, S.; Nan, L.; Kevin, S. The Current Status, Problems, and Prospects of Alfalfa (Medicago sativa L.) Breeding in China. Agronomy 2017, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Hufbauer, R.A. Pea aphid-parasitoid interactions: Have parasitoids adapted to differential resistance? Ecol. 2001, 82, 717–725. [Google Scholar] [CrossRef]
- Gao, L.L.; Klingler, J.P.; Anderson, J.P.; Edwards, O.R.; Singh, K.B. Characterization of pea aphid resistance in Medicago truncatula. Plant Physiol. 2008, 146, 996–1009. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Fan, Y.; Zhang, W.; Tian, Z.; Zhao, K. Soybean aphid, aphis glycines (hemiptera: Aphididae), developmental and reproductive capacity on white clover, Trifolium repens (rosales: Leguminosae), in northeast china. Appl. Entomol. Zool. 2017, 52, 491–495. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Li, C.; Liu, R. Indirect interactions between arbuscular mycorrhizal fungi and Spodoptera exigua alter photosynthesis and plant endogenous hormones. Mycorrhiza 2017, 27, 525–535. [Google Scholar] [CrossRef]
- Charters, M.D.; Sait, S.M.; Field, K.J. Aphid herbivory drives asymmetry in carbon for nutrient exchange between plants and an arbuscular mycorrhizal fungus. Curr. Biol. 2020, 30, 1801–1808. [Google Scholar] [CrossRef]
- Moloi, M.J.; Westhuizen, A.J. The reactive oxygen species are involved in resistance responses of wheat to the Russian wheat aphid. J. Plant Physiol. 2006, 163, 1118–1125. [Google Scholar] [CrossRef]
- Ahmad, P.; Sarwat, M.; Sharma, S. Reactive oxygen species, antioxidants and signaling in plants. J. Plant Biol. 2008, 51, 167–173. [Google Scholar] [CrossRef]
- Sabljic, I.; Barneto, J.A.; Balestrasse, K.B.; Zavala, J.A.; Pagano, E.A. Role of reactive oxygen species and isoflavonoids in soybean resistance to the attack of the southern green stink bug. PeerJ 2020, 8, e9956. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Sun, Y.; Peng, X.; Wang, Q.; Marvin, H.; Ge, F. Up-regulation of abscisic acid signaling pathway facilitates aphid xylem absorption and osmoregulation under drought stress. J. Exp. Bot. 2016, 67, 681–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Nan, Z.; Duan, T. Rhizophagus intraradices promotes alfalfa (Medicago sativa) defense against pea aphids (Acyrthosiphon pisum) revealed by RNA-Seq analysis. Mycorrhiza 2019, 29, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Mumm, R.; Dicke, M. Variation in natural plant products and the attraction of bodyguards involved in indirect plant defense. Can. J. Zool. 2010, 88, 628–667. [Google Scholar] [CrossRef]
- Schwartzberg, E.G.; Boeroeczky, K.; Tumlinson, J.H. Pea Aphids, Acyrthosiphon pisum, suppress induced plant volatiles in broad bean, Vicia Faba. J. Chem. Ecol. 2011, 37, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Jaeger, D.M.; Runyon, J.B.; Richardson, B.A. Signals of speciation: Volatile organic compounds resolve closely related sagebrush taxa, suggesting their importance in evolution. New Phytol. 2016, 211, 1393–1401. [Google Scholar] [CrossRef] [Green Version]
- Loreto, F.; Schnitzler, J.P. Abiotic stresses and induced BVOCs. Trends Plant Sci. 2010, 15, 154–166. [Google Scholar] [CrossRef]
- Cascone, P.; Iodice, L.; Maffei, M.E.; Bossi, S.; Arimura, G.; Guerrieri, E. Tobacco overexpressing beta-ocimene induces direct and indirect responses against aphids in receiver tomato plants. J. Plant Physiol. 2015, 173, 28–32. [Google Scholar] [CrossRef]
- Du, Y.J.; Poppy, G.M.; Powell, W. Relative importance of semiochemicals from first and second trophic levels in host foraging behavior of Aphidius ervi. J. Chem. Ecol. 1996, 22, 1591–1605. [Google Scholar] [CrossRef]
- Yang, D.; Lei, Z.; Bedair, M.; Sumner, L.W. An Optimized SPME-GC-MS Method for Volatile Metabolite Profiling of Different Alfalfa (Medicago sativa L.) Tissues. Molecules 2021, 26, 6473. [Google Scholar] [CrossRef]
- Formenti, L.; Rasmann, S. Mycorrhizal fungi enhance resistance to herbivores in tomato plants with reduced jasmonic acid production. Agronomy 2019, 9, 131. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Wees, S.V.; Bakker, P. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, P.; Shi, Z.; Diao, F.; Hao, L.; Zhang, J.; Xu, J.; Wang, L.; Dang, Z.; Guo, W. Effects of arbuscular mycorrhizal fungi on growth and Na plus accumulation of suaeda glauca (bunge) grown in salinized wetland soils. Appl. Soil Ecol. 2021, 166, 104065. [Google Scholar] [CrossRef]
- Babikova, Z.; Gilbert, L.; Bruce, T.; Dewhirst, S.Y.; Pickett, J.A.; Johnson, D. Arbuscular mycorrhizal fungi and aphids interact by changing host plant quality and volatile emission. Funct. Ecol. 2014, 28, 375–385. [Google Scholar] [CrossRef] [Green Version]
- Meier, A.R.; Hunter, M.D. Mycorrhizae alter constitutive and herbivore-induced volatile emissions by milkweeds. J. Chem. Ecol. 2019, 45, 610–625. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Duan, T.; Nan, Z. Arbuscular mycorrhizal fungus alleviates alfalfa leaf spots caused by Phoma medicaginis revealed by RNA-seq analysis. J. Appl. Microbiol. 2021, 130, 547–560. [Google Scholar] [CrossRef]
- Lorenzo, C.D.; Pedro, G.; Antonietti, M.S.; Maximiliano, S.; Estefanía, M.; Dezar, C.A.; Vazquez, M.; Watson, G.; Yanovsky, M.J.; Cerdán, P.D. Improvement of alfalfa forage quality and management through the down-regulation of MsFTa1. Plant Biotechnol. J. 2020, 18, 944–954. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wang, M.; Li, X.; Wang, X.; Jia, C.; Yang, X.; Feng, R.; Yuan, M. A small set of differentially expressed genes was associated with two color morphs in natural populations of the pea aphid Acyrthosiphon pisum. Gene 2018, 651, 23–32. [Google Scholar] [CrossRef]
- Gao, P.; Li, Y.; Guo, Y.; Duan, T. Co-inoculation of lucerne (Medicago sativa) with an AM fungus and a rhizobium reduces occurrence of spring black stem and leaf spot caused by Phoma medicaginis. Crop Pasture Sci. 2018, 69, 933–943. [Google Scholar] [CrossRef]
- Gonda, I.; Bar, E.; Portnoy, V.; Lev, S.; Burger, J.; Schaffer, A.A.; Lewinsohn, E. Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L. fruit. J. Exp. Bot. 2010, 61, 1111–1123. [Google Scholar] [CrossRef]
- Tholl, D.; Boland, W.; Hansel, A.; Loreto, F.; Röse, U.S.; Schnitzler, J.P. Practical approaches to plant volatile analysis. Plant J. 2006, 45, 540–560. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; An, M.; Li, X.; Han, Z.; Li, S.; Li, B. Molybdenum-induced effects on nitrogen absorption and utilization under different nitrogen sources in Vitis vinifera. J. Plant Interact. 2022, 17, 756–765. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedure for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158. [Google Scholar] [CrossRef]
- Giovannetti, M.; Mosse, B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Gao, J.; Zhao, B.; Dong, S.; Liu, P.; Zhang, J. Response of summer maize photosynthate accumulation and distribution to shading stress assessed by using 13CO2 stable isotope tracer in the field. Front. Plant Sci. 2017, 8, 1821. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Liu, D.; Song, P.; Jiang, F.; Zhang, T. Exposure to hypoxia causes stress erythropoiesis and downregulates immune response genes in spleen of mice. BMC Genom. 2021, 22, 413. [Google Scholar] [CrossRef]
- Shi, Q.; Hu, B.; Yang, C.; Deng, S.; Cheng, X.; Wu, J.; Qi, N. ATF3 inhibits arsenic-induced malignant transformation of human bronchial epithelial cells by attenuating inflammation. Toxicology 2021, 460, 152890. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Guo, W.; Peng, T.; Zhang, F.; Hao, Q.; Wang, G. Integrative analyses of nontargeted volatile profiling and transcriptome data provide molecular insight into voc diversity in cucumber plants (Cucumis sativus). Plant Physiol. 2016, 172, 603–618. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Ponzio, C.; Weldegergis, B.T.; Dicke, M.; Gols, R. Compatible and incompatible pathogen-plant interactions differentially affect plant volatile emissions and the attraction of parasitoid wasps. Funct. Ecol. 2016, 30, 1779–1789. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, X.X.; Liu, Y.M.; Liu, D.Y.; Du, Y.F.; Chen, X.P.; Zou, C. The role of phosphorus supply in maximizing the leaf area, photosynthetic rate, coordinated to grain yield of summer maize. Field Crops Res. 2018, 219, 113–119. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Babikova, Z.; Gilbert, L.; Randall, K.C.; Bruce, T.J.A.; Pickett, J.A.; Johnson, D. Increasing phosphorus supply is not the mechanism by which arbuscular mycorrhiza increase attractiveness of bean (Vicia faba) to aphids. J. Exp. Bot. 2014, 65, 5231–5241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thipyapong, P.; Steffens, J.C. Tomato polyphenol oxidase—Differential response of the polyphenol oxidase F promoter to injuries and wound signals. Plant Physiol. 1997, 115, 409–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahanil, S.; Attajarusit, J.; Stout, M.J.; Thipyapong, P. Overexpression of tomato polyphenol oxidase increases resistance to common cutworm. Plant Sci. 2008, 174, 456–466. [Google Scholar] [CrossRef]
- Grant, J.J.; Loake, G.J. Role of reactive oxygen intermediates and cognate redox signaling in disease resistance. Plant Physiol. 2000, 124, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, S.; Bao, Q.; Chu, Y.; Sun, H.; Huang, Y. Jasmonic acid alleviates cadmium toxicity through regulating the antioxidant response and enhancing the chelation of cadmium in rice (Oryza sativa L.). Environ. Pollut. 2022, 304, 119178. [Google Scholar] [CrossRef]
- Goggin, F.L.; Fischer, H.D. Reactive oxygen species in plant interactions with aphids. Front. Plant Sci. 2022, 12, 811105. [Google Scholar] [CrossRef]
- Zhao, H.; Xia, S.; Ming, X.; Xiao, Z.; Li, Q.; Hong, J.K. Antioxidant enzyme responses induced by whiteflies in tobacco plants in defense against aphids: Catalase may play a dominant role. PLoS ONE 2016, 11, e0165454. [Google Scholar] [CrossRef] [Green Version]
- Ryu, H.; Cho, Y.G. Plant hormones in salt stress tolerance. J. Integr. Plant Biol. 2015, 58, 147–155. [Google Scholar] [CrossRef]
- Dempsey, D.A.; Klessig, D.F. How does the multifaceted plant hormone salicylic acid combat disease in plants and are similar mechanisms utilized in humans? BMC Biol. 2017, 15, 23. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Wang, X.; Zu, H.; Zeng, X.; Li, R. Molecular dissection of rice phytohormone signaling involved in resistance to a piercing-sucking herbivore. New Phytol. 2021, 230, 1639–1652. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.M.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Exogenous jasmonic acid modulates the physiology, antioxidant defense and glyoxalase systems in imparting drought stress tolerance in different Brassica species. Plant Biotechnol. Rep. 2014, 8, 279–293. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Ghassemi-Golezani, K. How can salicylic acid and jasmonic acid mitigate salt toxicity in soybean plants? Ecotoxicol. Environ. Saf. 2018, 147, 1010–1016. [Google Scholar] [CrossRef] [PubMed]
- Gunes, A.; Inal, A.; Alpaslan, M.; Cicek, N.; Guneri, E.; Eraslan, F.; Guzelordu, T. Effects of exogenously applied salicylic acid on the induction of multiple stress tolerance and mineral nutrition in maize (Zea mays L.). Arch. Agron. Soil Sci. 2005, 51, 687–695. [Google Scholar] [CrossRef]
- Stevens, J.; Senaratna, T.; Sivasithamparam, K. Salicylic acid induces salinity tolerance in tomato (Lycopersicon esculentum cv. Roma): Associated changes in gas exchange, water relations and membrane stabilisation. Plant Growth Regul. 2006, 49, 77–83. [Google Scholar]
- Ku, Y.S.; Sintaha, M.; Cheung, M.Y.; Lam, H.M. Plant hormone signaling crosstalks between biotic and abiotic stress responses. Int. J. Mol. Sci. 2018, 19, 3206. [Google Scholar] [CrossRef] [Green Version]
- Jang, G.; Yoon, Y.; Yang, D.C. Crosstalk with jasmonic acid integrates multiple responses in plant development. Int. J. Mol. Sci. 2020, 21, 305. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Hernandez, C.; Martinez-Gallardo, N.; Guerrero-Rangel, A.; Valdes-Rodriguez, S.; Delano-Frier, J. Trypsin and alpha-amylase inhibitors are differentially induced in leaves of amaranth (Amaranthus hypochondriacus) in response to biotic and abiotic stress. Plant Physiol. 2004, 122, 254–264. [Google Scholar] [CrossRef]
- Babikova, Z.; Gilbert, L.; Bruce, T.J.A.; Birkett, M.; Caulfield, J.C.; Woodcock, C.; Pickett, J.A.; Johnson, D. Underground signals carried through common mycelial networks warn neighbouring plants of aphid attack. Ecol. Lett. 2013, 16, 835–843. [Google Scholar] [CrossRef]
- Skoczek, A.; Piesik, D.; Wenda-Piesik, A.; Buszewski, B.; Bocianowski, J.; Wawrzyniak, M. Volatile organic compounds released by maize following herbivory or insect extract application and communication between plants. J. Appl. Entomol. 2016, 141, 630–643. [Google Scholar] [CrossRef]
- Effah, E.; Barrett, D.P.; Peterson, P.G.; Godfrey, A.J.R.; Potter, M.A.; Holopainen, J.K.; McCormick, A.C. Natural Variation in Volatile Emissions of the Invasive Weed Calluna vulgaris in New Zealand. Plants 2020, 9, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallini, A.; Janssen, A.; Sabelis, M.W. Odour-mediated responses of phytophagous mites to conspecific and heterospecific competitors. Oecologia 1997, 110, 179–185. [Google Scholar] [CrossRef] [PubMed]
- De Moraes, C.M.; Mescher, M.C.; Tumlinson, J.H. Caterpillar-induced nocturnal plant volatiles repel conspecific females. Nature 2001, 410, 577–580. [Google Scholar] [CrossRef] [PubMed]
- Mauck, K.E.; Moraes, C.M.D.; Mescher, M.C. Deceptive chemical signals induced by a plant virus attract insect vectors to inferior hosts. Proc. Natl. Acad. Sci. USA 2010, 107, 3600–3605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visser, J.H.; Piron, P.G.M. Olfactory antennal responses to plant volatiles in apterous virginoparae of the vetch aphid megoura viciae. Entomol. Exp. Appl. 1995, 77, 37–46. [Google Scholar] [CrossRef]
- Zhang, Z.; Bian, L.; Sun, X.; Luo, Z.; Xin, Z.; Luo, F.; Chen, Z. Electrophysiological and behavioural responses of the tea geometrid ectropis obliqua (lepidoptera: Geometridae) to volatiles from a non-host plant, rosemary, Rosmarinus officinalis (lamiaceae). Pest Manag. Sci. 2015, 71, 96–104. [Google Scholar] [CrossRef]
- Razmjou, J.; Davari, M.; Ebadollahi, A. Effect of two plant essential oils and the entomopathogenic fungus, lecanicillium muscarium (zare & gams) on the cotton aphid, aphis gossypii glover. Egypt. J. Biol. Pest Control 2016, 26, 775–779. [Google Scholar]
- Masimbula, R.; Oki, K.; Takahashi, K.; Matsuura, H. Metabolism of airborne methyl salicylate in adjacent plants. Biosci. Biotechnol. Biochem. 2020, 84, 1780–1787. [Google Scholar] [CrossRef]
- Park, S.W.; Kaimoyo, E.; Kumar, D.; Mosher, S.; Klessig, D.F. Methyl salicylate is a critical mobile signal for plant systemic acquired resistance. Science 2007, 318, 113–116. [Google Scholar] [CrossRef]
- Kalaivani, K.; Kalaiselvi, M.M.; Senthil-Nathan, S. Effect of methyl salicylate (MeSA), an elicitor on growth, physiology and pathology of resistant and susceptible rice varieties. Sci. Rep. 2016, 6, 34498. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Mcgillen, M.R.; Dale, V.; Casas, J.; Mellouki, A. The fate of methyl salicylate in the environment and its role as signal in multitrophic interactions. Sci. Total Environ. 2020, 749, 141406. [Google Scholar] [CrossRef] [PubMed]
- Williams, L.; Rodriguez-Saona, C.; Castle, S.C.; Zhu, S. EAG-active herbivore-induced plant volatiles modify behavioral responses and host attack by an egg parasitoid. J. Chem. Ecol. 2008, 34, 1190–1201. [Google Scholar] [CrossRef] [PubMed]
- Salamanca, J.; Souza, B.; Lundgren, J.G.; Rodriguez-Saona, C. From laboratory to field: Electro-antennographic and behavioral responsiveness of two insect predators to methyl salicylate. Chemoecology 2017, 27, 51–63. [Google Scholar] [CrossRef]
- Mallinger, R.E.; Hogg, D.B.; Claudio, G. Methyl salicylate attracts natural enemies and reduces populations of soybean aphids (Hemiptera: Aphididae) in soybean agroecosystems. J. Econ. Entomol. 2011, 104, 115–124. [Google Scholar] [CrossRef]
- Lee, J.C.; Flores, S.M.; Graham, K.; Skillman, V.P. Methyl salicylate can benefit ornamental pest control, and does not alter per capita predator consumption at close-range. Front. Ecol. Environ. 2022, 9, 788187. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
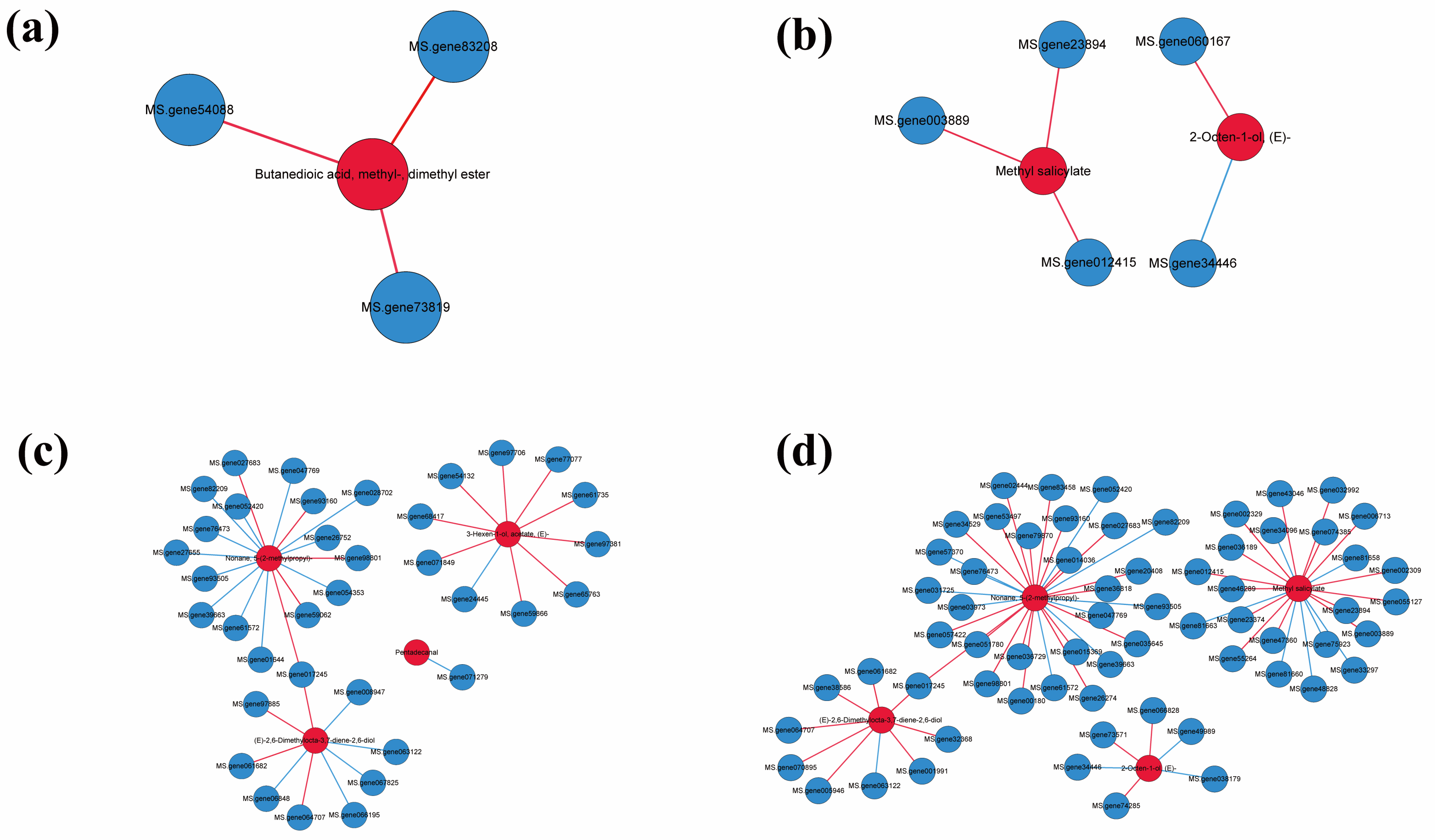{kind=link}
| Control | Treatment | Compounds | VIP | p_Value | Fold Change | Log2FC |
|---|---|---|---|---|---|---|
| NMA− | AMA- | Butanedioic acid, methyl-, dimethyl ester | 1.596 | 0.00034495 | 4.849 | 2.278 |
| NMA− | NMA+ | 2-Octen-1-ol, (E)− | 1.280 | 0.026913339 | 0.424 | 1.237 |
| NMA− | NMA+ | Methyl salicylate | 1.694 | 0.00390791 | 0.360 | 1.475 |
| AMA− | AMA+ | Pentadecanal | 1.618 | 0.048801556 | 2.064 | 1.045 |
| AMA− | AMA+ | (E)-2,6-Dimethylocta-3,7-diene-2,6-diol | 1.962 | 0.029184939 | 3.582 | 1.841 |
| AMA− | AMA+ | Nonane, 5-(2-methylpropyl)- | 2.166 | 0.000394176 | 3.454 | 1.788 |
| AMA− | AMA+ | 3-Hexen-1-ol, acetate, (E)− | 1.866 | 0.045902066 | 3.711 | 1.892 |
| NMA+ | AMA+ | 2-Octen-1-ol, (E)− | 1.433 | 0.031982472 | 3.122 | 1.642 |
| NMA+ | AMA+ | Methyl salicylate | 1.610 | 0.007071354 | 5.098 | 2.350 |
| NMA+ | AMA+ | (E)-2,6-Dimethylocta-3,7-diene-2,6-diol | 1.478 | 0.034961913 | 3.214 | 1.685 |
| NMA+ | AMA+ | Nonane, 5-(2-methylpropyl)- | 1.666 | 0.00029612 | 3.207 | 1.681 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Li, Y.; Tian, Z.; Duan, T. Arbuscular Mycorrhizal Fungus Alters Alfalfa (Medicago sativa) Defense Enzyme Activities and Volatile Organic Compound Contents in Response to Pea Aphid (Acyrthosiphon pisum) Infestation. J. Fungi 2022, 8, 1308. https://doi.org/10.3390/jof8121308
Wang Y, Li Y, Tian Z, Duan T. Arbuscular Mycorrhizal Fungus Alters Alfalfa (Medicago sativa) Defense Enzyme Activities and Volatile Organic Compound Contents in Response to Pea Aphid (Acyrthosiphon pisum) Infestation. Journal of Fungi. 2022; 8(12):1308. https://doi.org/10.3390/jof8121308
Chicago/Turabian StyleWang, Yajie, Yingde Li, Zhen Tian, and Tingyu Duan. 2022. "Arbuscular Mycorrhizal Fungus Alters Alfalfa (Medicago sativa) Defense Enzyme Activities and Volatile Organic Compound Contents in Response to Pea Aphid (Acyrthosiphon pisum) Infestation" Journal of Fungi 8, no. 12: 1308. https://doi.org/10.3390/jof8121308