
Two New Species and a New Record of Microdochium from Grasses in Yunnan Province, South-West China

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Isolation, and Identification
2.2. DNA Extraction, PCR Amplification, and DNA Sequencing
2.3. Phylogenetic Analyses
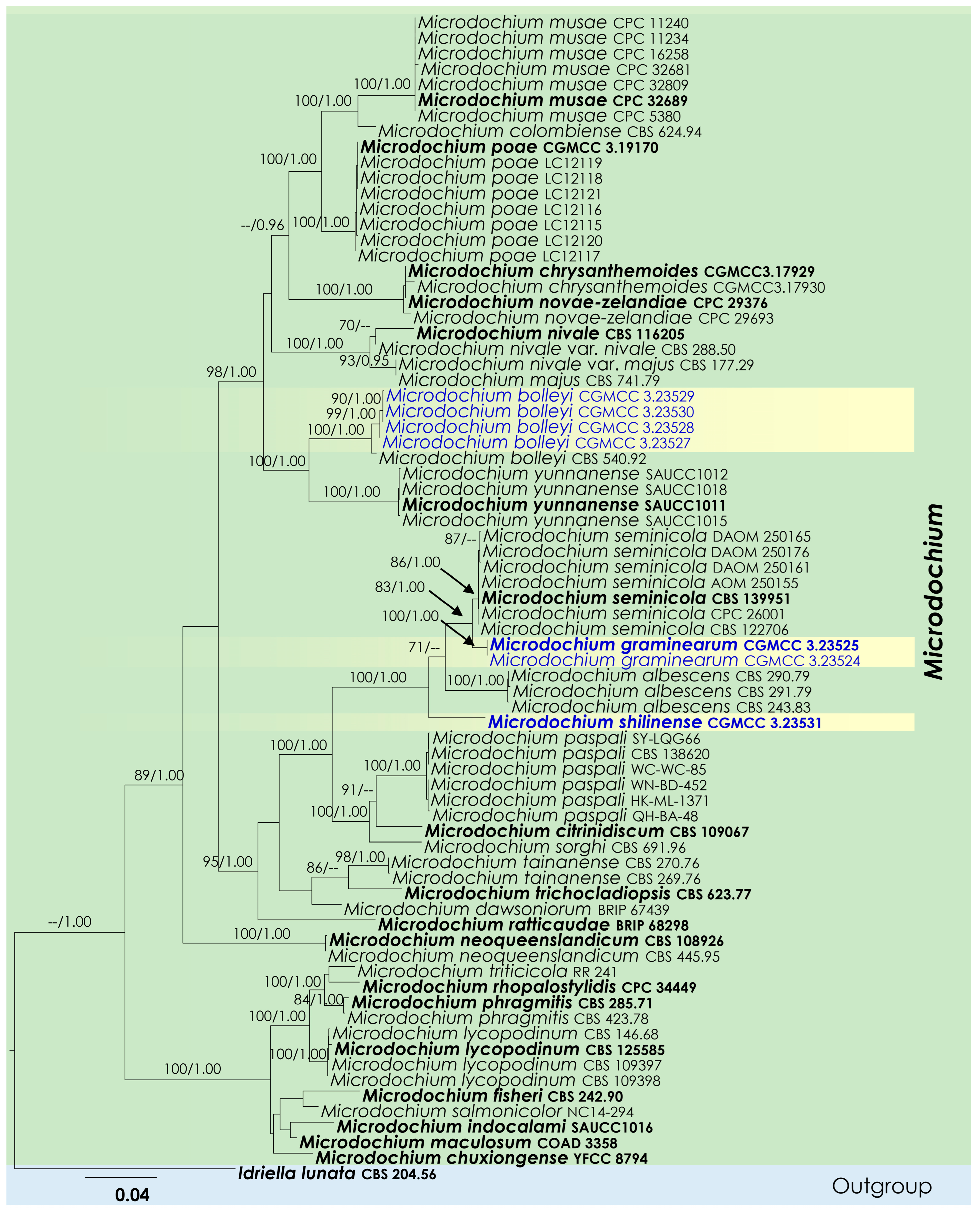
3. Results
3.1. Phylogenetic Analyses
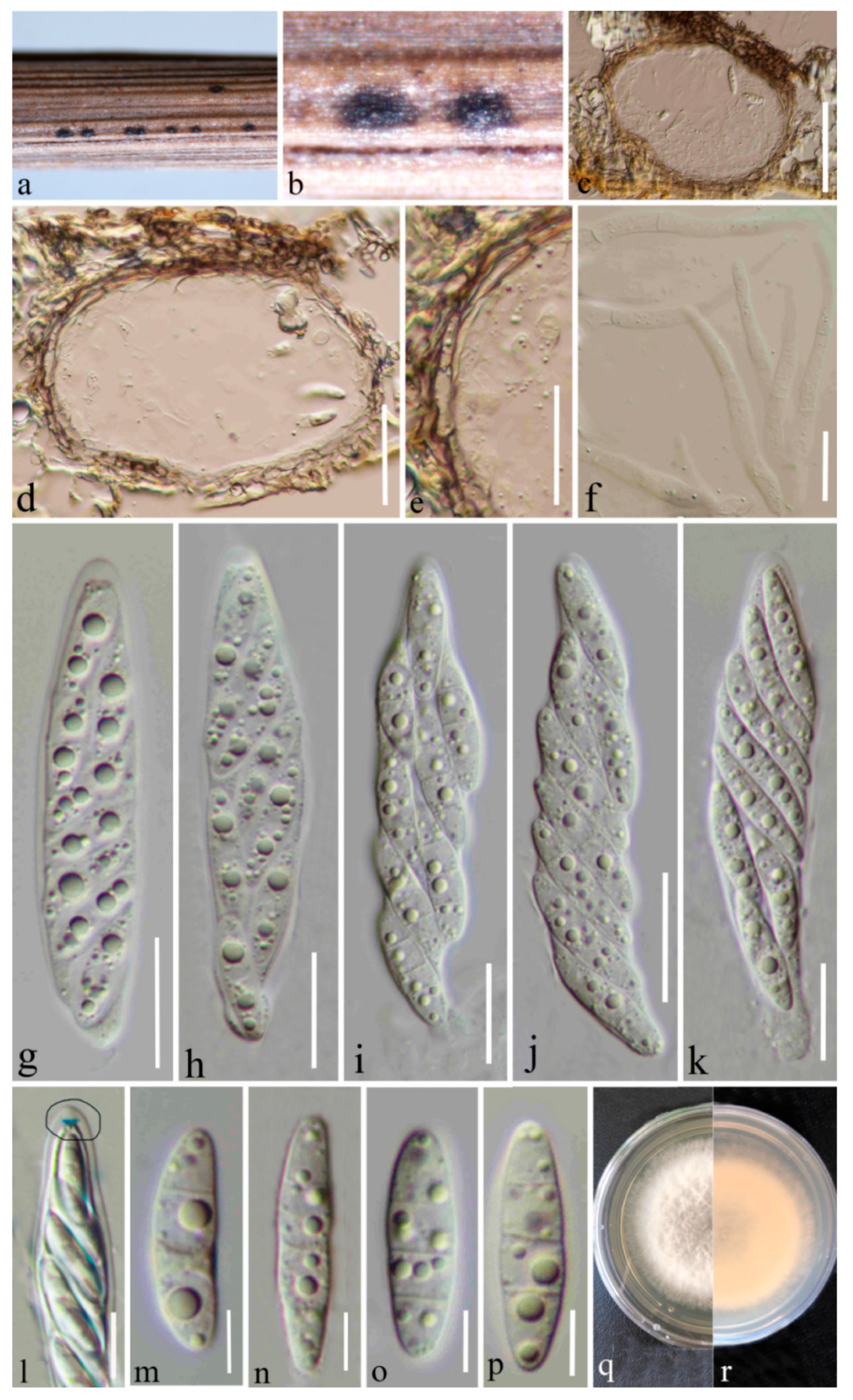
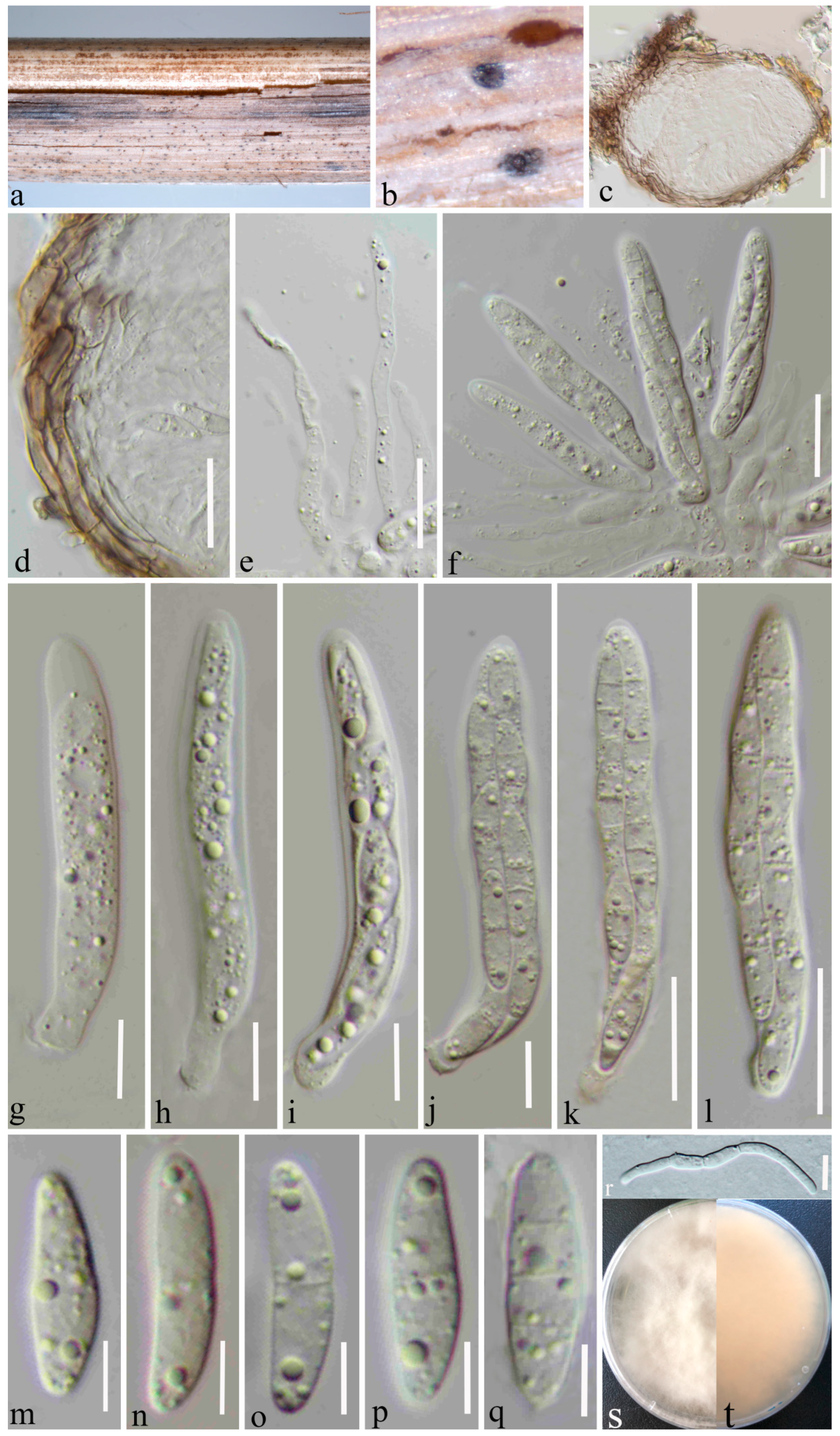
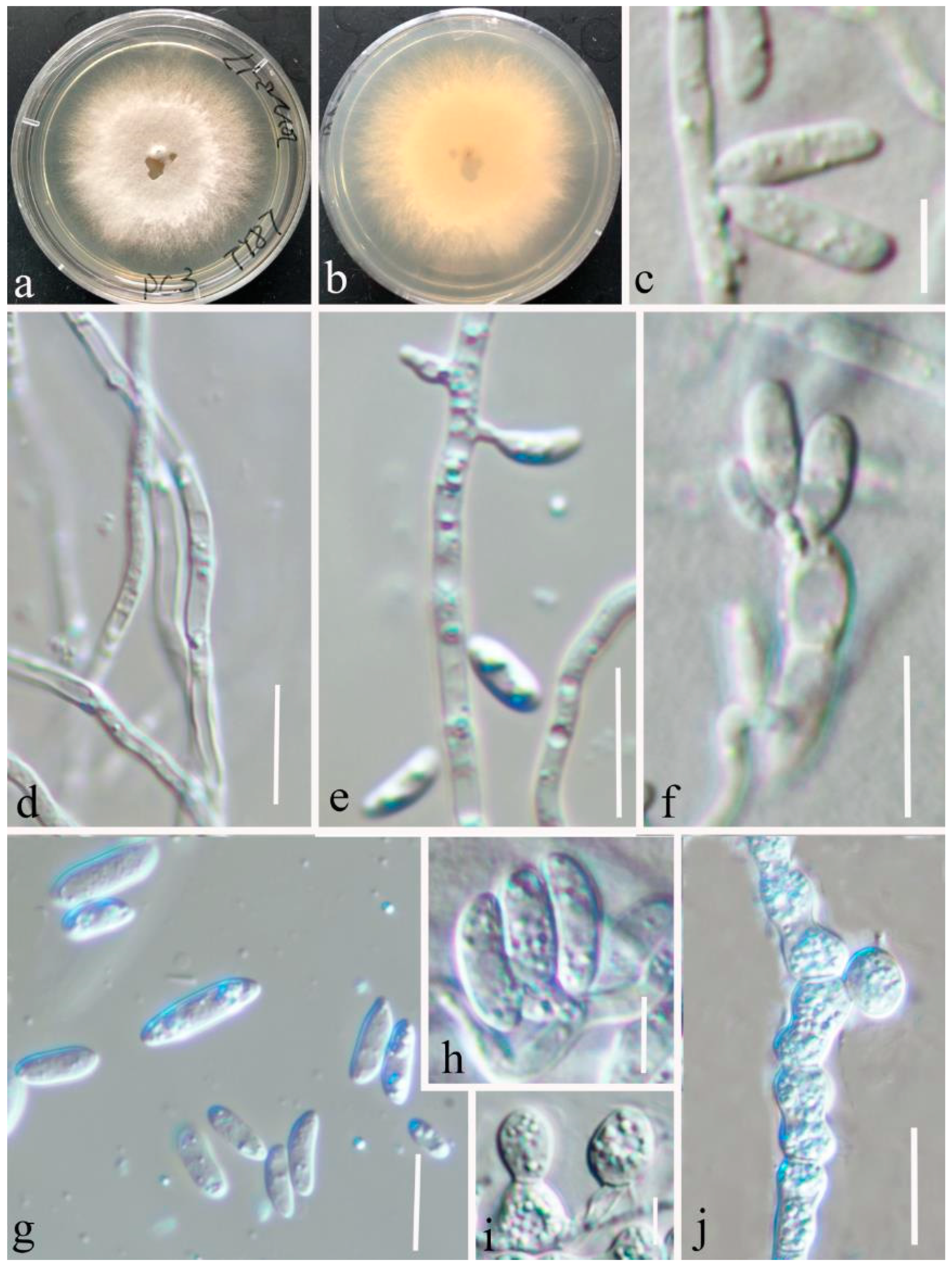
3.2. Taxonomy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T.; Saxena, R.K.; Erdoğdu, M.; Selçuk, F.; Rajeshkumar, K.C.; Aptroot, A.; et al. Outline of Fungi and fungus-like taxa–2021. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Sydow, H. Mycotheca Germanica Fasc. XLII-XLV (No. 2051–2250). Ann. Mycol. 1924, 22, 257–268. [Google Scholar]
- Hernández-Restrepo, M.; Groenewald, J.Z.; Crous, P.W. Taxonomic and phylogenetic re-evaluation of Microdochium, Monographella and Idriella. Pers. Mol. Phylogeny Evol. Fungi 2016, 36, 57–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.F.; Liu, F.; Zhou, X.; Liu, X.Z.; Liu, S.J.; Cai, L. Culturable mycobiota from Karst caves in China, with descriptions of 20 new species. Persoonia 2017, 39, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Schumacher, R.K.; Wingfield, M.J.; Akulov, A.; Denman, S.; Roux, J.; Braun, U.; Burgess, T.I.; Carnegie, A.J.; Váczy, K.Z.; et al. New and interesting fungi 1. Fungal Syst. Evol. 2018, 1, 169–215. [Google Scholar] [CrossRef]
- Crous, P.W.; Schumacher, R.K.; Akulov, A.; Thangavel, R.; Hernández-Restrepo, M.; Carnegie, A.J.; Cheewangkoon, R.; Wingfield, M.J.; Summerell, B.A.; Quaedvlieg, W.; et al. New and interesting fungi 2. Fungal Syst. Evol. 2019, 3, 57–134. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Chooi, Y.-H.; Gilchrist, C.L.M.; Lacey, E.; Pitt, J.I.; Roets, F.; Swart, W.J.; Cano-Lira, J.F.; Valenzuela-Lopez, N.; et al. Fungal Planet description sheets: 1042–1111. Persoonia 2020, 44, 301–459. [Google Scholar] [CrossRef]
- Crous, P.W.; Cowan, D.A.; Maggs-Kölling, G.; Yilmaz, N.; Thangavel, R.; Wingfield, M.J.; Noordeloos, M.E.; Dima, B.; Brandrud, T.E.; Jansen, G.M.; et al. Fungal Planet description sheets: 1182–1283. Persoonia 2021, 46, 313–528. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Osieck, E.R.; Jurjevi, Ž; Boers, J.; Van Iperen, A.L.; Starink-Willemse, M.; Dima, B.; Balashov, S.; Bulgakov, T.S.; Johnston, P.R.; et al. Fungal Planet description sheets: 1284–1382. Persoonia 2021, 47, 178–374. [Google Scholar] [CrossRef]
- Marin-Felix, Y.; Hernández-Restrepo, M.; Wingfield, M.J.; Akulov, A.; Carnegie, A.J.; Cheewangkoon, R.; Gramaje, D.; Groenewald, J.Z.; Guarnaccia, V.; Halleen, F.; et al. Genera of phytopathogenic fungi: GOPHY 2. Stud. Mycol. 2019, 92, 47–133. [Google Scholar] [CrossRef]
- Huang, S.T.; Xia, J.W.; Zhang, X.G.; Sun, W.X.; Li, Z. Two new species of Microdochium from Indocalamus longiauritus in south-western China. MycoKeys 2020, 72, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Index Fungorum. Available online: http://www.indexfungorum.org/Names/Names.asp (accessed on 9 September 2022).
- Das, K.; Lee, S.Y.; Jung, H.Y. Molecular and Morphological Characterization of Two Novel Species Collected from Soil in Korea. Mycobiology 2020, 48, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.X.; Zou, W.Q.; Wang, Y.M.; Huang, O.; Yu, H. Morphology and phylogeny of Microdochium chuxiongense sp. nov., a fungus from Southwest China. Phytotaxa 2022, 555, 147–158. [Google Scholar] [CrossRef]
- Von Arx, J.A. Plant pathogenic fungi. Nova Hedwig. 1987, 87, 1–288. [Google Scholar]
- Glynn, N.; Hare, M.; Parry, D.; Edwards, S. Phylogenetic analysis of EF-1 alpha gene sequences from isolates of Microdochium nivale leads to elevation of varieties majus and nivale to species status. Mycol. Res. 2005, 109, 872–880. [Google Scholar] [CrossRef]
- Jewell, L.E.; Hsiang, T. Multigene differences between Microdochium Nivale and Microdochium majus. Botany 2013, 91, 99–106. [Google Scholar] [CrossRef]
- Holubová-Jechová, V.; Mercado Sierra, A. Some new or interesting microfungi from Cuba. Mycotaxon 1982, 14, 309–315. [Google Scholar]
- Samuels, G.J.; Müller, E.; Petrini, O. Studies on the Amphisphaeriaceae (sensu lato) 3. New species of Monographella and Pestalosphaeria and two new genera. Mycotaxon 1987, 28, 473–499. [Google Scholar]
- Sutton, B.C.; Pirozynski, K.A.; Deighton, F.C. Microdochium Syd. Can. J. Bot. 1972, 50, 1899–1907. [Google Scholar] [CrossRef]
- Sutton, B.C.; Hodges, C.S. Eucalyptus microfungi: Microdochium and Phaeoisaria species from Brazil. Nova Hedwig. 1976, 27, 215–222. [Google Scholar]
- Hoog, G.S.; Hermanides-Nijhof, E.J. The black yeasts and allied Hyphomycetes. Stud. Mycol. 1977, 15, 1–222. [Google Scholar]
- Ernst, M.; Neubert, K.; Mendgen, K.W.; Wirsel, S.G.R. Niche differentiation of two sympatric species of Microdochium colonizing the roots of common reed. BMC Microbiol. 2011, 11, 242–254. [Google Scholar] [CrossRef] [Green Version]
- Jaklitsch, W.M.; Voglmayr, H. Phylogenetic relationships of five genera of Xylariales and Rosasphaeria gen. nov. (Hypocreales). Fungal Divers. 2012, 52, 75–98. [Google Scholar] [CrossRef]
- Liu, Y.; Zachow, C.; Raaijmakers, J.M.; de Bruijn, I. Elucidating the diversity of aquatic Microdochium and Trichoderma species and their activity against the fish pathogen Saprolegnia diclina. Int. J. Mol. Sci. 2016, 17, 140–154. [Google Scholar] [CrossRef] [Green Version]
- Berg, G.; Zachow, C.; Lottmann, J.; Gotz, M.; Costa, R.; Smalla, K. Impact of plant species and site on rhizosphere-associated fungi antagonistic to Verticillium dahliae Kleb. Appl. Environ. Microbiol. 2005, 71, 4203–4213. [Google Scholar] [CrossRef]
- Bhosale, S.H.; Patil, K.B.; Parameswaran, P.S.; Naik, C.G.; Jagtap, T.G. Active pharmaceutical ingredient (api) from an estuarine fungus, Microdochium nivale (Fr.). J. Environ. Biol. 2011, 32, 653–658. [Google Scholar]
- Santiago, I.F.; Alves, T.; Rabello, A.; Sales Junior, P.A.; Romanha, A.J.; Zani, C.L. Leishmanicidal and antitumoral activities of endophytic fungi associated with the Antarctic angiosperms Deschampsia antarctica Desv. and Colobanthus quitensis (Kunth) Bartl. Extremophiles 2012, 16, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.; Baghela, A.; Singh, S.K. Morphology and phylogeny of Microdochium fisheri, a new record from India. Plant Pathol. Quar. 2017, 7, 191–200. [Google Scholar] [CrossRef]
- Bhunjun, C.S.; Niskanen, T.; Suwannarach, N.; Wannathes, N.; Chen, Y.J.; McKenzie, E.H.; Maharachchikumbura, S.S.; Buyck, B.; Zhao, C.L.; Fan, Y.G.; et al. The numbers of fungi: Are the most speciose genera truly diverse? Fungal Divers. 2022, 114, 387–462. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Rathnayaka, A.R.; Marasinghe, D.S.; Calabon, M.S.; Gentekaki, E.; Lee, H.B.; Hurdeal, V.G.; Pem, D.; Dissanayake, L.S.; Wijesinghe, S.N.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Gao, Y.H.; Sun, W.; Su, Y.Y.; Cai, L. Three new species of Phomopsis in Gutianshan Nature Reserve in China. Mycol. Prog. 2014, 13, 111–121. [Google Scholar] [CrossRef]
- Cai, L.; Hyde, K.D.; Taylor, P.W.J.; Weir, B.S.; Waller, J.; Abang, M.M.; Zhang, J.Z.; Yang, Y.L.; Phoulivong, S.; Liu, Z.Y.; et al. A polyphasic approach for studying Colletotrichum. Fungal Divers. 2009, 39, 183–204. [Google Scholar]
- Ren, G.C.; Wanasinghe, D.N.; Jeewon, R.; Monkai, J.; Mortimer, P.E.; Hyde, K.D.; Xu, J.C.; Gui, H. Taxonomy and phylogeny of the novel rhytidhysteron-like collections in the Greater Mekong Subregion. MycoKeys 2022, 86, 65–85. [Google Scholar] [CrossRef] [PubMed]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, D.J.; Buyck, B.; Cai, L.; Dai, Y.C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The Faces of Fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Chaiwan, N.; Gomdola, D.; Wang, S.; Monkai, J.; Tibpromma, S.; Doilom, M.; Wanasinghe, D.N.; Mortimer, P.E.; Lumyong, S.; Hyde, K.D. https://gmsmicrofungi.org:: An online database providing updated information of microfungi in the Greater Mekong Subregion. Mycosphere 2021, 12, 1513–1526. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Jones, E.B.G.; Dissanayake, A.J.; Hyde, K.D. Saprobic Dothideomycetes in Thailand: Vaginatispora appendiculata sp. nov. (Lophiostomataceae) introduced based on morphological and molecular data. Stud. Fungi 2016, 1, 56–68. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic Relationships among Ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Sung, G.H.; Sung, J.M.; Hywel-Jones, N.L.; Spatafora, J.W. A multigene phylogeny of Clavicipitaceae (Ascomycota, Fungi): Identification of localized incongruence using a combinational bootstrap approach. Mol. Phylogenetics Evol. 2007, 44, 1204–1223. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author. Evolutionary Biology Centre, Uppsala University 2004. Available online: https://github.com/nylander/MrModeltest2/releases (accessed on 16 September 2020).
- Rambaut, A. FigTree v1.4.2, a Graphical Viewer of Phylogenetic Trees. 2014. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 5 December 2020).
- Goonasekara, I.D.; Jayawardene, R.S.; Saichana, N.; Hyde, K.D. Checklist of microfungi on grasses in Thailand (excluding bambusicolous fungi). Asian J. Mycol. 2018, 1, 88–105. [Google Scholar] [CrossRef]
- Thambugala, K.M.; Wanasinghe, D.N.; Phillips, A.J.L.; Camporesi, E.; Bulgakov, T.S.; Phukhamsakda, C.; Ariyawansa, H.A.; Goonasekara, I.D.; Phookamsak, R.; Dissanayake, A.; et al. Mycosphere notes 1–50: Grass (Poaceae) inhabiting. Mycosphere 2017, 8, 697–796. [Google Scholar] [CrossRef]
- Braun, U. A Monograph of Cercosporella, Ramularia, and Allied Genera (Phytopathogenic Hyphomycetes); Ihw-Verlag: Eching, Germany, 1995; Volume 1. [Google Scholar]
- Hong, S.K.; Kim, W.G.; Choi, H.W. Identification of Microdochium bolleyi associated with basal rot of creeping bent grass in Korea. Mycobiology 2008, 36, 77–80. [Google Scholar] [CrossRef] [Green Version]
- Savulescu, T. Einige neue Micromyceten. Bull. Sect. Sci. Acad. Roum. 1942, 25, 22–30. [Google Scholar]
- Müller, E.; Samuels, G.J. Monographella maydis sp. nov. and its connection to the tar-spot disease of Zea mays. Nova Hedwig. 1984, 40, 112–120. [Google Scholar]
- Samuels, G.J.; Hallett, I.C. Microdochium stoveri and Monographella stoveri, new combinations for Fusarium stoveri and Micronectriella stoveri. Trans. Br. Mycol. Soc. 1983, 81, 473–483. [Google Scholar] [CrossRef]
- von Arx, J.A. Notes on Monographella and Microdochium. Trans. Br. Mycol. Soc. 1984, 83, 373–374. [Google Scholar] [CrossRef]
- Galea, V.J.; Price, T.V.; Sutton, B.C. Taxonomy and biology of the lettuce anthracnose fungus. Trans. Br. Mycol. Soc. 1986, 86, 619–628. [Google Scholar] [CrossRef]
- Zhang, W.; Nan, Z.B.; Tian, P.; Hu, M.J.; Gao, Z.Y.; Li, M.; Liu, G.D. Microdochium paspali, a new species causing seashore paspalum disease in southern China. Mycologia 2015, 107, 80–89. [Google Scholar] [CrossRef]
- Liang, J.M.; Li, G.S.; Zhao, M.Q.; Cai, L. A new leaf blight disease of turfgrasses caused by Microdochium poae, sp. nov. Mycologia 2019, 111, 265–273. [Google Scholar] [CrossRef]
- Braun, U. Studies on Ramularia and allied genera (VI). Nova Hedwig. 1993, 56, 423–454. [Google Scholar]
- Matsushima, T. Matsushima Mycological Memoirs No. 6; Matsushima Fungus Collection: Kobe, Japan, 1989; Volume 6, pp. 1–100. [Google Scholar]
- Mouchacca, J.; Samson, R.A. Deux nouvelles espèces du genre Microdochium Sydow. Rev. Mycol. 1973, 37, 267–275. [Google Scholar]
- Petrak, F. Mykologische Notizen. IX. Ann. Mycol. 1927, 25, 193–343. [Google Scholar]
- Kwasna, H.; Bateman, G.L. Microdochium Triticicola sp. nov. from roots of Triticum aestivum in the United Kingdom. Mycologia 2007, 99, 765–776. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes/Loci | PCR Primers (Forward/Reverse) | PCR Annealing Thermal Conditions | References |
|---|---|---|---|
| ITS | ITS5/ITS4 | 55 °C for 15 s | [39] |
| LSU | LR0R/LR5 | [40] | |
| tub2 | Btub526F and Btub1332R | 55 °C for 30 s | [18] |
| rpb2 | fRPB2-5F2/fRPB2-7cR | 57 °C for 50 s | [41,42] |
| Species Name | Strain Numbers | GenBank Accession Numbers | |||
|---|---|---|---|---|---|
| LSU | ITS | tub2 | rpb2 | ||
| Idriella lunata | CBS 204.56 * | KP858981 | KP859044 | NA | NA |
| Microdochium albescens | CBS 290.79 | KP858950 | KP859014 | KP859078 | KP859123 |
| M. albescens | CBS 291.79 | KP858932 | KP858996 | KP859059 | KP859105 |
| M. albescens | CBS 243.83 | KP858930 | KP858994 | KP859057 | KP859103 |
| M. bolleyi | CBS 540.92 | KP858946 | KP859010 | KP859073 | KP859119 |
| M. bolleyi | CGMCC 3.23527 | OP104018 | OP103968 | OP242830 | NA |
| M. bolleyi | CGMCC 3.23528 | OP104019 | OP103969 | OP242831 | NA |
| M. bolleyi | CGMCC 3.23529 | OP104020 | OP103970 | OP242832 | OP184897 |
| M. bolleyi | CGMCC 3.23530 | OP104021 | OP103971 | OP242833 | OP184898 |
| M. chrysanthemoides | LC 5363 * | KU746736 | KU746690 | NA | NA |
| M. chrysanthemoides | LC 5466 | KU746735 | KU746689 | NA | NA |
| M. citrinidiscum | CBS 109067 * | KP858939 | KP859003 | KP859066 | KP859112 |
| M. colombiense | CBS 624.94 * | KP858935 | KP858999 | KP859062 | KP859108 |
| M. chuxiongense | YFCC 8794 * | OK586160 | OK586161 | OK556901 | OK584019 |
| M. dawsoniorum | BRIP 67439 | NA | MN492650 | NA | NA |
| M. fisheri | CBS 242.90 * | KP858951 | KP859015 | KP859079 | KP859124 |
| M. graminearum | CGMCC 3.23524 | OP104015 | OP103965 | OP242835 | OP236026 |
| M. graminearum | CGMCC 3.23525 * | OP104016 | OP103966 | OP236029 | OP236027 |
| M. indocalami | SAUCC 1016 * | MT199878 | MT199884 | MT435653 | MT510550 |
| M. lycopodinum | CBS 125585 * | KP858952 | KP859016 | KP859080 | KP859125 |
| M. lycopodinum | CBS 146.68 | KP858929 | KP858993 | KP859056 | KP859102 |
| M. lycopodinum | CBS 109397 | KP858940 | KP859004 | KP859067 | KP859113 |
| M. lycopodinum | CBS 109398 | KP858941 | KP859005 | KP859068 | KP859114 |
| M. maculosum | COAD 3358 * | OK966953 | OK966954 | NA | NA |
| M. majus | CBS 741.79 | KP858937 | KP859001 | KP859064 | KP859110 |
| M. musae | CBS 111018 | NA | AY293061 | NA | NA |
| M. musae | CBS 143499 | MH107941 | MH107894 | NA | NA |
| M. musae | CBS 143500 * | MH107942 | MH107895 | NA | MH108003 |
| M. musae | CPC:11234 | MH107943 | MH107896 | NA | NA |
| M. musae | CPC:11240 | MH107944 | MH107897 | NA | NA |
| M. musae | CPC:16258 | MH107945 | MH107898 | NA | NA |
| M. musae | CPC:32681 | MH107946 | MH107899 | NA | NA |
| M. neoqueenslandicum | CBS 445.95 | KP858933 | KP858997 | KP859060 | KP859106 |
| M. neoqueenslandicum | CBS 108926 * | KP858938 | KP859002 | KP859065 | KP859111 |
| M. nivale | CBS 116205 * | KP858944 | KP859008 | KP859071 | KP859117 |
| M. nivale var. majus | CBS 177.29 | MH866500 | MH855031 | NA | NA |
| M. nivale var. nivale | CBS 288.50 | MH868135 | MH856626 | NA | NA |
| M. novae-zelandiae | CPC:29376 * | NG_066396 | NR_172274 | LT990608 | LT990641 |
| M. novae-zelandiae | CPC:29693 | LT990628 | LT990656 | LT990609 | LT990642 |
| M. paspali | CBS 138620 * | NA | NR_158810 | NA | NA |
| M. paspali | CBS138620 | NA | KJ569509 | KJ569514 | NA |
| M. paspali | QH-BA-48 | NA | KJ569510 | KJ569515 | NA |
| M. paspali | SY-LQG66 | NA | KJ569511 | KJ569516 | NA |
| M. paspali | WC-WC-85 | NA | KJ569512 | KJ569517 | NA |
| M. paspali | WN-BD-452 | NA | KJ569513 | KJ569518 | NA |
| M. phragmitis | CBS 285.71 * | KP858949 | KP859013 | KP859077 | KP859122 |
| M. phragmitis | CBS 423.78 | KP858948 | KP859012 | KP859076 | KP859121 |
| M. poae | CGMCC 3.19170 * | NA | MH740898 | MH740914 | MH740906 |
| M. poae | LC 12115 | NA | MH740901 | MH740917 | MH740909 |
| M. poae | LC 12116 | NA | MH740902 | MH740918 | MH740910 |
| M. poae | LC 12117 | NA | MH740903 | MH740919 | MH740911 |
| M. poae | LC 12118 | NA | MH740897 | MH740913 | MH740905 |
| M. poae | LC 12119 | NA | MH740899 | MH740915 | MH740907 |
| M. poae | LC 12120 | NA | MH740904 | MH740920 | MH740912 |
| M. poae | LC 12121 | NA | MH740900 | MH740916 | MH740908 |
| M. ratticaudae | BRIP 68298 * | MW481666 | MW481661 | NA | MW626890 |
| M. rhopalostylidis | CPC:34449 * | MK442532 | MK442592 | NA | MK442667 |
| M. salmonicolor | NC14-294 | MK836108 | MK836110 | NA | NA |
| M. seminicola | KAS 3576 * | KP858974 | KP859038 | KP859101 | KP859147 |
| M. seminicola | KAS 1516 | KP858961 | KP859025 | KP859088 | KP859134 |
| M. seminicola | KAS 3574 | KP858973 | KP859037 | KP859100 | KP859146 |
| M. seminicola | KAS 3158 | KP858970 | KP859034 | KP859097 | KP859143 |
| M. seminicola | KAS 1527 | KP858966 | KP859030 | KP859093 | KP859139 |
| M. seminicola | KAS 1473 | KP858955 | KP859019 | KP859082 | KP859128 |
| M. seminicola | CBS 122706 | KP858943 | KP859007 | KP859070 | KP859116 |
| M. shilinense | CGMCC 3.23531 * | OP104022 | OP103972 | OP242834 | NA |
| M. sorghi | CBS 691.96 | KP858936 | KP859000 | KP859063 | KP859109 |
| M. tainanense | CBS 269.76 * | KP858945 | KP859009 | KP859072 | KP859118 |
| M. tainanense | CBS 270.76 | KP858931 | KP858995 | KP859058 | KP859104 |
| M. trichocladiopsis | CBS 623.77 * | KP858934 | KP858998 | KP859061 | KP859107 |
| M. triticicola | RR 241 | NA | AJ748691 | NA | NA |
| M. yunnanense | SAUCC 1011 * | MT199875 | MT199881 | MT435650 | MT510547 |
| M. yunnanense | SAUCC 1012 | MT199876 | MT199882 | NA | MT510548 |
| M. yunnanense | SAUCC 1015 | MT199877 | MT199883 | MT435652 | MT510549 |
| M. yunnanense | SAUCC 1018 | MT199880 | MT199886 | MT435655 | NA |
| Name of Taxon | Host | Place | Life-Mode | References |
|---|---|---|---|---|
| Microdochium albescens | Oryza sativa | Ivory Coast | Plant pathogen | [4] |
| M. bolleyi | Gramineae, wood, Setaria parviflora | North Dakota, U.S.A.; Syria, Canada; China | Plant pathogen, endophyte | [23,52,53], this study |
| M. caespitosum | Dead leaves | Tanzania | Saprophyte | [21] |
| M. chrysanthemoides | Air of a karst cave | China | ― | [5] |
| M. citrinidiscum | Leaf of Eichhornia crassipes | Peru | Pathogen | [4] |
| M. colombiense | Musa sapientum | Colombia | ― | [4] |
| M. chuxiongense | On pileus of Bondarzewia sp. | China | ― | [15] |
| M. consociatum | ― | San Jorge Province (Ecuador) | ― | [4] |
| M. cylindricum | Dead leaves of Eucalyptus | Brazil | Saprophyte | [22] |
| M. dawsoniorum | Leaves of Sporobolus natalensis | Australia | ― | [8] |
| M. fisheri | Stem of Oryzae sativa, Rhizospheric paddy soil | U.K.; India | Endophyte | [4,30] |
| M. fusariisporum | Dead straw of Panicum virgatum | Kansas, U.S.A. | Saprophyte | [4] |
| M. graminearum | Gramineae | China | Saprophyte | This study |
| M. griseum | Dead leaves of Sapium ellipticum | Tanzania | Saprophyte | [21] |
| M. indocalami | Leaves of Indocalamus longiauritus | China | Plant pathogen | [12] |
| M. intermedium | Soi1 | Papua New Guinea | ― | [23] |
| M. linariae | Stem | Italy | ― | [54] |
| M. lycopodinum | Lycopodium annotinum, Phragmites australis, air, salmon eggs | Austria; Germany; Netherlands | Non-pathogenic | [4,25,26] |
| M. maculosum | Leaves of Digitaria insularis | Brazil | Plant pathogen | [10] |
| M. majus | On Triticum aestivum | Germany | Plant pathogen | [4,17] |
| M. maydis | Leaves of Zea mays | Mexico | Plant pathogen | [4,55] |
| M. musae | Leaves of Musa sp. | China (Taiwan) | Plant pathogen | [6] |
| M. neoqueenslandicum | Juncus effusus, Agrostis sp. | Waihi, New Zealand; Netherlands | Plant pathogen | [4] |
| M. nivale | Roots of Triticum aestivum; Porteresia coarctata | UK | Plant pathogen | [28,56] |
| M. novae-zelandiae | Leaves of Poaceae | New Zealand | Plant pathogen | [11] |
| M. opuntiae | Dead leaves of Oputia | Louisiana, U.S.A.; Langlois | Plant pathogen | [4,57] |
| M. oryzae | Oryzae sativa | Japan | Plant pathogen | [56] |
| M. palmicola | Dead petiole of Roystonea regia | Cuba | Saprophyte | [19] |
| M. panattonianum | Leaves of Lactuca sativa | Denmark | Plant pathogen | [58] |
| M. paspali | Paspalum vaginatum | China (Hainan) | Pathogen | [59] |
| M. passiflorae | Dead stem of Passiflora edulis | New Zealand | Saprophyte | [20] |
| M. phragmitis | Phragmitis communis, Phragmites australis, salmon eggs, angiosperms | Germany; Poland; Antarctic | Endophyte | [3,26,29] |
| M. phyllanthi | Leaves of Phyllanthus discoideus | Germany; Poland | Plant pathogen | [21] |
| M. poae | Leaves of Poa pratensis and Agrostis stolonifera | China | Plant pathogen | [60] |
| M. punctum | Stem of Sisyrinchii campestris | U.S.A. | ― | [61] |
| M. queenslandicum | Forest soil | Australia | ― | [62] |
| M. ratticaudae | Stem of Sporobolus natalensis (Poaceae) | Australia | ― | [9] |
| M. rhopalostylidis | Leaves of Rhopalostylis sapida | New Zealand | Plant pathogen | [7] |
| M. salmonicolor | Soil | Korea | ― | [14] |
| M. sclerotiorum | Culture contaminant | Netherlands | ― | [63] |
| M. seminicola | Grain seeds, barley, Triticum aestivum | Canada; Switzerland | Plant pathogen | [4] |
| M. shilinense | Gramineae | China | Saprophyte | This study |
| M. sorghi | Leaves of Sorghum vulgaris | Louisiana, U.S.A.; Cuba | Pathogen | [12,16,52] |
| M. stevensonii | Panicum hemitomon | Florida, U.S.A. | ― | [4,64] |
| M. stoveri | Musa sp. | Honduras, Central America | Plant pathogen | [56] |
| M. tainanense | Root of Saccharum officinarum | Japan; China (Taiwan) | Rhizosphere fungus | [4,23] |
| M. trichocladiopsis | Rhizosphere of Triticum aestivum | Unknown country | Rhizosphere fungus | [4] |
| M. triticicola | Roots of Triticum aestivum | UK | Plant pathogen | [65] |
| M. yunnanense | Leaves of Indocalamus longiauritus | China | Plant pathogen | [12] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Ren, G.-C.; Wanasinghe, D.N.; Xu, J.-C.; Gomes de Farias, A.R.; Gui, H. Two New Species and a New Record of Microdochium from Grasses in Yunnan Province, South-West China. J. Fungi 2022, 8, 1297. https://doi.org/10.3390/jof8121297
Gao Y, Ren G-C, Wanasinghe DN, Xu J-C, Gomes de Farias AR, Gui H. Two New Species and a New Record of Microdochium from Grasses in Yunnan Province, South-West China. Journal of Fungi. 2022; 8(12):1297. https://doi.org/10.3390/jof8121297
Chicago/Turabian StyleGao, Ying, Guang-Cong Ren, Dhanushka N. Wanasinghe, Jian-Chu Xu, Antonio Roberto Gomes de Farias, and Heng Gui. 2022. "Two New Species and a New Record of Microdochium from Grasses in Yunnan Province, South-West China" Journal of Fungi 8, no. 12: 1297. https://doi.org/10.3390/jof8121297