
Thiamine Supplementation Improves Survival and Body Condition of Hatchery-Reared Steelhead (Oncorhynchus mykiss) in Oregon

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
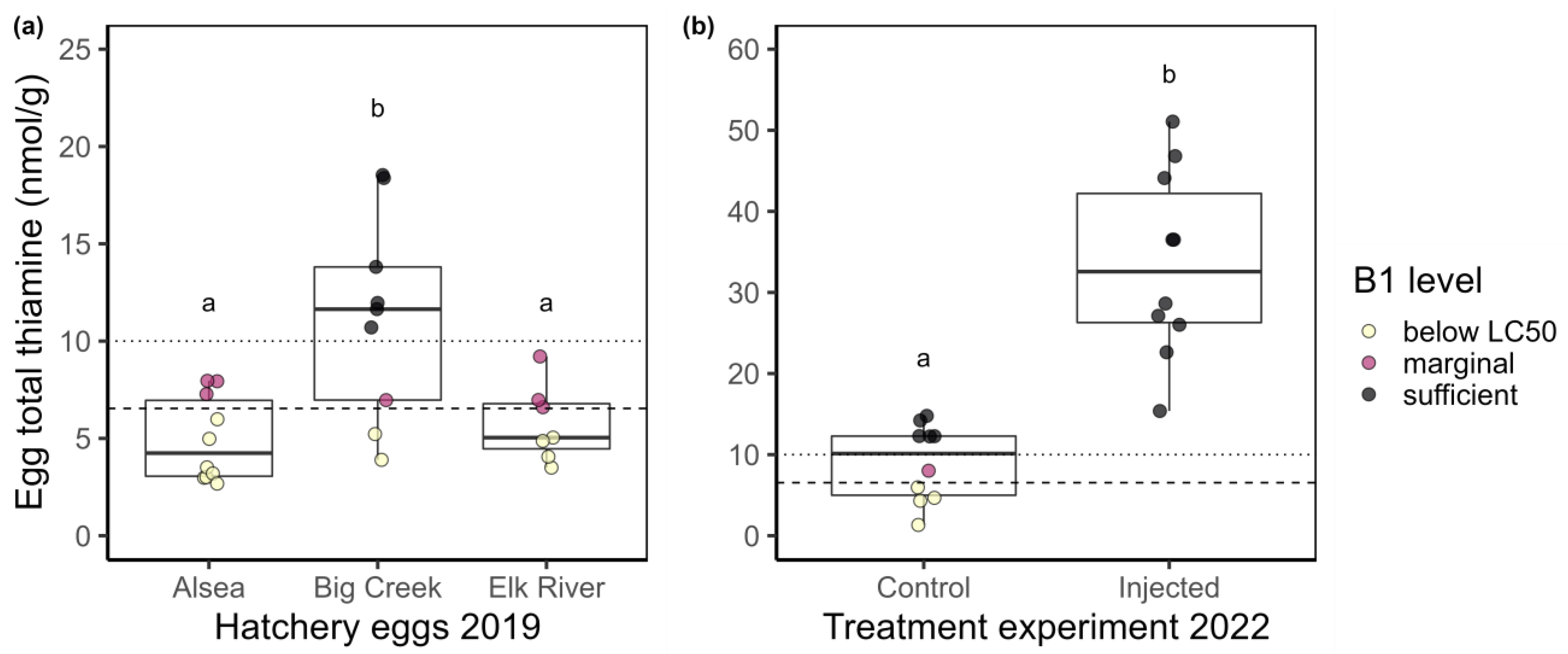
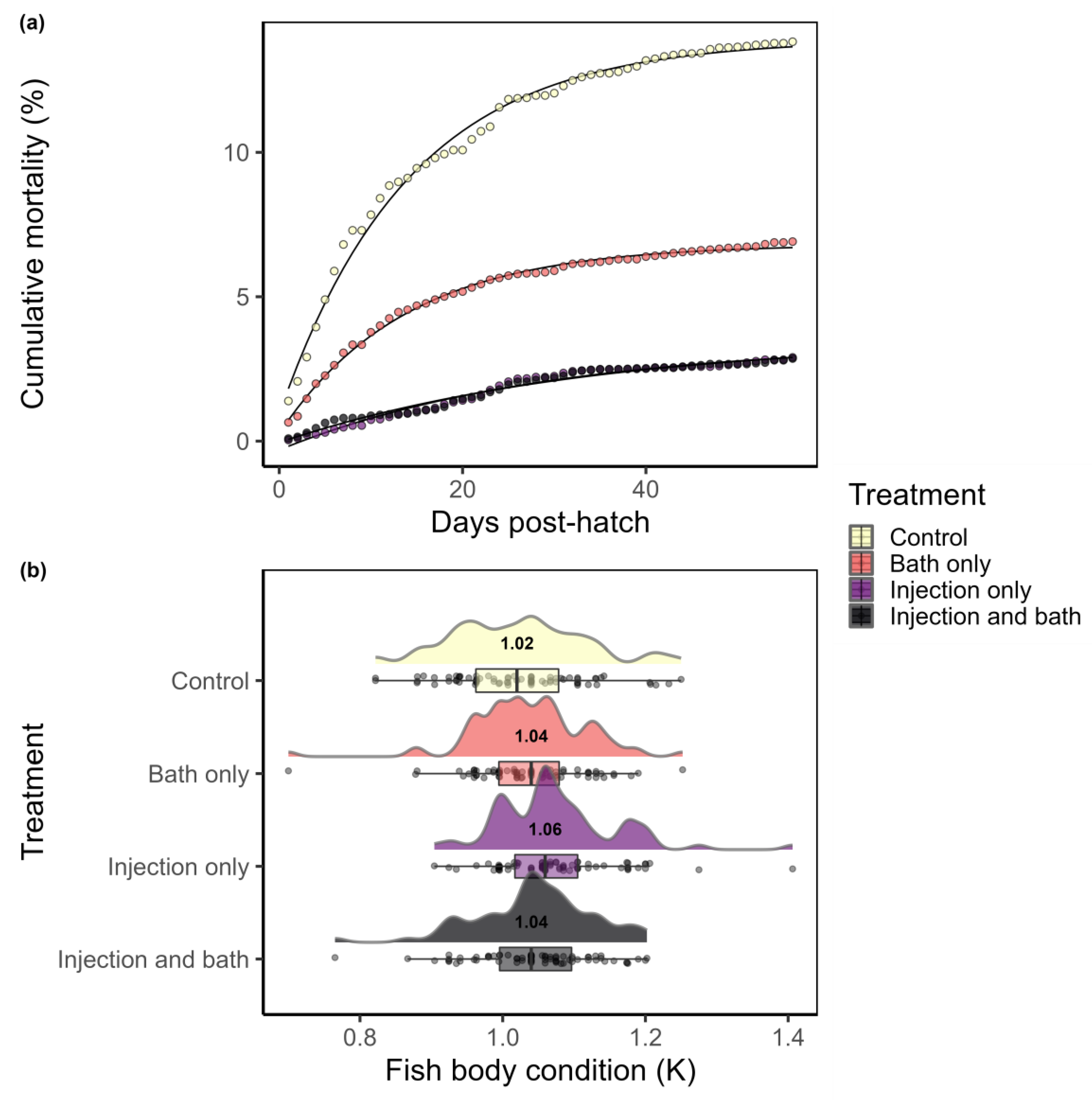
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Whitfield, K.C.; Bourassa, M.W.; Adamolekun, B.; Bergeron, G.; Bettendorff, L.; Brown, K.H.; Cox, L.; Fattal-Valevski, A.; Fischer, P.R.; Frank, E.L.; et al. Thiamine Deficiency Disorders: Diagnosis, Prevalence, and a Roadmap for Global Control Programs. Ann. N. Y. Acad. Sci. 2018, 1430, 3–43. [Google Scholar] [CrossRef] [PubMed]
- Vuorinen, P.J.; Rokka, M.; Ritvanen, T.; Käkelä, R.; Nikonen, S.; Pakarinen, T.; Keinänen, M. Changes in Thiamine Concentrations, Fatty Acid Composition, and Some Other Lipid-Related Biochemical Indices in Baltic Sea Atlantic Salmon (Salmo Salar) during the Spawning Run and Pre-Spawning Fasting. Helgol. Mar. Res. 2020, 74, 10. [Google Scholar] [CrossRef]
- Kraft, C.E.; Angert, E.R. Competition for Vitamin B1 (Thiamin) Structures Numerous Ecological Interactions. Q. Rev. Biol. 2017, 92, 151–168. [Google Scholar] [CrossRef] [PubMed]
- Harder, A.M.; Ardren, W.R.; Evans, A.N.; Futia, M.H.; Kraft, C.E.; Marsden, J.E.; Richter, C.A.; Rinchard, J.; Tillitt, D.E.; Christie, M.R. Thiamine Deficiency in Fishes: Causes, Consequences, and Potential Solutions. Rev. Fish Biol. Fish. 2018, 28, 865–886. [Google Scholar] [CrossRef]
- Honeyfield, D.C.; Hinterkopf, J.P.; Fitzsimons, J.D.; Tillitt, D.E.; Zajicek, J.L.; Brown, S.B. Development of Thiamine Deficiencies and Early Mortality Syndrome in Lake Trout by Feeding Experimental and Feral Fish Diets Containing Thiaminase. J. Aquat. Anim. Health 2005, 17, 4–12. [Google Scholar] [CrossRef]
- Honeyfield, D.C.; Ross, J.P.; Carbonneau, D.A.; Terrell, S.P.; Woodward, A.R.; Schoeb, T.R.; Perceval, H.F.; Hinterkopf, J.P. Pathology, Physiologic Parameters, Tissue Contaminants, And Tissue Thiamine In Morbid And Healthy Central Florida Adult American Alligators (Alligator mississippiensis). J. Wildl. Dis. 2008, 44, 280–294. [Google Scholar] [CrossRef]
- Balk, L.; Hägerroth, P.-Å.; Gustavsson, H.; Sigg, L.; Åkerman, G.; Ruiz Muñoz, Y.; Honeyfield, D.C.; Tjärnlund, U.; Oliveira, K.; Ström, K.; et al. Widespread Episodic Thiamine Deficiency in Northern Hemisphere Wildlife. Sci. Rep. 2016, 6, 38821. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.B.; Fitzsimons, J.D.; Honeyfield, D.C.; Tillitt, D.E. Implications of Thiamine Deficiency in Great Lakes Salmonines. J. Aquat. Anim. Health 2005, 17, 113–124. [Google Scholar] [CrossRef]
- Riley, S.C.; Rinchard, J.; Honeyfield, D.C.; Evans, A.N.; Begnoche, L. Increasing Thiamine Concentrations in Lake Trout Eggs from Lakes Huron and Michigan Coincide with Low Alewife Abundance. North Am. J. Fish. Manag. 2011, 31, 1052–1064. [Google Scholar] [CrossRef]
- Fitzsimons, J.D.; Williston, B.; Williston, G.; Brown, L.; El-Shaarawi, A.; Vandenbyllaardt, L.; Honeyfeld, D.; Tillitt, D.; Wolgamood, M.; Brown, S.B. Egg Thiamine Status of Lake Ontario Salmonines 1995–2004 with Emphasis on Lake Trout. J. Gt. Lakes Res. 2007, 33, 93–103. [Google Scholar] [CrossRef]
- Amcoff, P.; Börjeson, H.; Lindeberg, J.; Norrgren, L. Thiamine Concentrations in Feral Baltic Salmon Exhibiting the M74 Syndrome. Am. Fish. Soc. Symp. 1998, 21, 82–89. [Google Scholar] [CrossRef]
- Engelhardt, J.; Frisell, O.; Gustavsson, H.; Hansson, T.; Sjöberg, R.; Collier, T.K.; Balk, L. Severe Thiamine Deficiency in Eastern Baltic Cod (Gadus morhua). PLoS ONE 2020, 15, e0227201. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, V.; Staykov, Y.; Tzanova, M.; Valkova, E.; Krastev, B.; Dimitrov, Z.; Staykov, Y.; Tzanova, M.; Valkova, E.; Krastev, B.; et al. Reproductive Process in Bulgarian Trout Farms in Relation to the Prevention of M74 Syndrome. Bulg. J. Agric. Sci. 2017, 23, 147–153. [Google Scholar]
- Ketola, H.G.; Chiotti, T.L.; Rathman, R.S.; Fitzsimons, J.D.; Honeyfield, D.C.; Van Dusen, P.J.; Lewis, G.E. Thiamine Status of Cayuga Lake Rainbow Trout and Its Influence on Spawning Migration. North Am. J. Fish. Manag. 2005, 25, 1281–1287. [Google Scholar] [CrossRef]
- Mantua, N.; Johnson, R.; Field, J.; Lindley, S.; Williams, T.; Todgham, A.; Jeffres, C.; Bell, H.; Cocherell, D.; Rinchard, J.; et al. Mechanisms, Impacts, and Mitigation for Thiamine Deficiency and Early Life Stage Mortality in California’s Central Valley Chinook Salmon. North Pac. Anadromous Fish Comm. Tech. Rep. 2021, 17, 92–93. [Google Scholar] [CrossRef]
- Fitzsimons, J.D.; Williston, B.; Amcoff, P.; Balk, L.; Pecor, C.; Ketola, H.G.; Hinterkopf, J.P.; Honeyfield, D.C. The Effect of Thiamine Injection on Upstream Migration, Survival, and Thiamine Status of Putative Thiamine-Deficient Coho Salmon. J. Aquat. Anim. Health 2005, 17, 48–58. [Google Scholar] [CrossRef]
- Fisher, J.P.; Spitsbergen, J.M.; Iamonte, T. Pathological and Behavioral Manifestations of the “Cayuga Syndrome,” a Thiamine Deficiency in Larval Landlocked Atlantic Salmon. J. Aquat. Anim. Health 1995, 7, 269–283. [Google Scholar] [CrossRef]
- Fitzsimons, J.D.; Ketola, G.; Wooster, G.W.; Brown, S.B. Use of a Thiamine Antagonist to Induce Cayuga-Syndrome-like Mortalities in Larval Atlantic Salmon. J. Aquat. Anim. Health 2001, 13, 151–157. [Google Scholar] [CrossRef]
- Lundström, J.; Börjeson, H.; Norrgren, L. Histopathological Studies of Yolk-Sac Fry of Baltic Salmon (Salmo Salar) with the M74 Syndrome. Ambio 1999, 28, 16–23. [Google Scholar]
- Carvalho, P.S.M.; Tillitt, D.E.; Zajicek, J.L.; Claunch, R.A.; Honeyfield, D.C.; Fitzsimons, J.D.; Brown, S.B. Thiamine Deficiency Effects on the Vision and Foraging Ability of Lake Trout Fry. J. Aquat. Anim. Health 2009, 21, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.B.; Brown, L.R.; Brown, M.; Moore, K.; Villella, M.; Fitzsimons, J.D.; Williston, B.; Honeyfield, D.C.; Hinterkopf, J.P.; Tillitt, D.E.; et al. Effectiveness of Egg Immersion in Aqueous Solutions of Thiamine and Thiamine Analogs for Reducing Early Mortality Syndrome. J. Aquat. Anim. Health 2005, 17, 106–112. [Google Scholar] [CrossRef]
- Futia, M.H.; Hallenbeck, S.; Noyes, A.D.; Honeyfield, D.C.; Eckerlin, G.E.; Rinchard, J. Thiamine Deficiency and the Effectiveness of Thiamine Treatments through Broodstock Injections and Egg Immersion on Lake Ontario Steelhead Trout. J. Gt. Lakes Res. 2017, 43, 352–358. [Google Scholar] [CrossRef]
- Robards, M.D.; Quinn, T.P. The Migratory Timing of Adult Summer-Run Steelhead in the Columbia River over Six Decades of Environmental Change. Trans. Am. Fish. Soc. 2002, 131, 523–536. [Google Scholar] [CrossRef]
- Oregon Department of Fish and Wildlife. Fish Propagation Annual Report; Oregon Department of Fish and Wildlife: Salem, OR, USA, 2021; p. 189.
- National Oceanic and Atmospheric Administration. Endangered and Threatened Species; 5-Year Reviews for 28 Listed Species of Pacific Salmon, Steelhead, and Eulachon; Federal Register. 2016; Volume 81. Available online: https://www.federalregister.gov/ (accessed on 1 December 2022).
- Shoemaker, C.; Xu, D.-H.; LaFrentz, B.; LaPatra, S. Overview of Fish Immune System and Infectious Diseases. In Dietary Nutrients, Additives, and Fish Health; Lee, C.-S., Lim, C., Gatlin, D.M., Webster, C.D., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 1–24. ISBN 978-1-119-00556-8. [Google Scholar]
- Brown, S.B.; Fitzsimons, J.D.; Palace, V.P.; Vandenbyllaardt, L. Thiamine and Early Mortality Syndrome in Lake Trout. Am. Fish. Soc. Symp. 1998, 21, 18–25. [Google Scholar]
- Weerarathne, I.A. Sample-Size Requirements for Accurate Length-Frequency Distributions of Mesophotic Reef Fishes from Baited Remote Underwater Stereo Video. Ecol. Indic. 2021, 122, 107262. [Google Scholar] [CrossRef]
- Bagenal, T. Methods for Assessment of Fish Production in Fresh Waters, 3rd ed.; Blackwell Scientific Pub.: Oxford, UK, 1978. [Google Scholar]
- Zajicek, J.L.; Tillitt, D.E.; Brown, S.B.; Brown, L.R.; Honeyfield, D.C.; Fitzsimons, J.D. A Rapid Solid-Phase Extraction Fluorometric Method for Thiamine and Riboflavin in Salmonid Eggs. J. Aquat. Anim. Health 2005, 17, 95–105. [Google Scholar] [CrossRef]
- Tang, X.; Cronin, D.A.; Brunton, N.P. A Simplified Approach to the Determination of Thiamine and Riboflavin in Meats Using Reverse Phase HPLC. J. Food Compos. Anal. 2006, 19, 831–837. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing. 2021. Available online: https://www.R-project.org/ (accessed on 1 December 2022).
- Hothorn, T.; Hornik, K.; van de Wiel, M.; Zeileis, A. A Lego System for Conditional Inference. Am. Stat. 2006, 60, 257–263. [Google Scholar] [CrossRef]
- Mangiafico, S. Rcompanion: Functions to Support Extension Education Program Evaluation. 2023. Available online: https://CRAN.R-project.org/package=rcompanion (accessed on 1 December 2022).
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, 3rd ed.; Sage Publications: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- de Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. 2021. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 1 December 2022).
- Futia, M.H.; Rinchard, J. Evaluation of Adult and Offspring Thiamine Deficiency in Salmonine Species from Lake Ontario. J. Gt. Lakes Res. 2019, 45, 811–820. [Google Scholar] [CrossRef]
- Ivan, L.N.; Schmitt, B.R.; Rose, K.A.; Riley, S.C.; Rose, J.B.; Murphy, C.A. Evaluation of the Thiamine Dose-Response Relationship for Lake Trout (Salvelinus namaycush) Fry Using an Individual Based Model. J. Gt. Lakes Res. 2018, 44, 1393–1404. [Google Scholar] [CrossRef]
- Reed, A.; Rowland, F.; Krajcik, J.; Tillitt, D. Egg Thiamine Content, Length, Weight and Survival of Steelhead (Oncorhynchus Mykiss) Exposed to Three Methods of Experimental Thiamine Supplementation in 2022 and Egg Thiamine Content from Three Oregon Hatcheries in 2019: U.S. Geological Survey Data Release; U.S. Geological Survey: Reston, VA, USA, 2023. [CrossRef]
- Fitzsimons, J.D.; Brown, S.B.; Williston, B.; Williston, G.; Brown, L.R.; Moore, K.; Honeyfield, D.C.; Tillitt, D.E. Influence of Thiamine Deficiency on Lake Trout Larval Growth, Foraging, and Predator Avoidance. J. Aquat. Anim. Health 2009, 21, 302–314. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.P.; Brown, S.B.; Wooster, G.W.; Bowser, P.R. Maternal Blood, Egg and Larval Thiamin Levels Correlate with Larval Survival in Landlocked Atlantic Salmon (Salmo Salar). J. Nutr. 1998, 128, 2456–2466. [Google Scholar] [CrossRef] [PubMed]
- Van Doornik, D.M.; Kuligowski, D.R.; Morgan, C.A.; Burke, B.J.; Seamons, T.R. Insights, from Genetic Analyses, into Stock-Specific Distribution of Juvenile Steelhead (Oncorhynchus mykiss) from the Columbia River during Early Marine Migration. Fish. Bull. 2019, 117, 97–106. [Google Scholar] [CrossRef]
- Daly, E.A.; Scheurer, J.A.; Brodeur, R.D.; Weitkamp, L.A.; Beckman, B.R.; Miller, J.A. Juvenile Steelhead Distribution, Migration, Feeding, and Growth in the Columbia River Estuary, Plume, and Coastal Waters. Mar. Coast. Fish. 2014, 6, 62–80. [Google Scholar] [CrossRef]
- Thalmann, H.L.; Daly, E.A.; Brodeur, R.D. Two Anomalously Warm Years in the Northern California Current: Impacts on Early Marine Steelhead Diet Composition, Morphology, and Potential Survival. Trans. Am. Fish. Soc. 2020, 149, 369–382. [Google Scholar] [CrossRef]
- Myers, K.W. Ocean Ecology of Steelhead. In The Ocean Ecology of Salmon and Trout; American Fisheries Society: Bethesda, MA, USA, 2018; p. 126. ISBN 978-1-934874-45-5. [Google Scholar]
- Gåsnes, S.K.; Oliveira, V.H.S.; Gismervik, K.; Ahimbisibwe, A.; Tørud, B.; Jensen, B.B. Mortality Patterns during the Freshwater Production Phase of Salmonids in Norway. J. Fish Dis. 2021, 44, 2083–2096. [Google Scholar] [CrossRef]
- Ottinger, C.A.; Honeyfield, D.C.; Densmore, C.L.; Iwanowicz, L.R. Impact of Thiamine Deficiency on T-Cell Dependent and T-Cell Independent Antibody Production in Lake Trout. J. Aquat. Anim. Health 2012, 24, 258–273. [Google Scholar] [CrossRef]
- Ottinger, C.A.; Honeyfield, D.C.; Densmore, C.L.; Iwanowicz, L.R. In Vitro Immune Functions in Thiamine-Replete and -Depleted Lake Trout (Salvelinus Namaycush). Fish Shellfish Immunol. 2014, 38, 211–220. [Google Scholar] [CrossRef]
- Myers, K.; Irvine, J.; Logerwell, E.; Urawa, S.; Naydenko, S.; Zavolokin, A.; Davis, N. Pacific Salmon and Steelhead: Life in a Changing Winter Ocean. North Pac. Anadromous Fish Comm. Bull. 2016, 6, 113–138. [Google Scholar] [CrossRef]
- Crozier, L.G.; McClure, M.M.; Beechie, T.; Bograd, S.J.; Boughton, D.A.; Carr, M.; Cooney, T.D.; Dunham, J.B.; Greene, C.M.; Haltuch, M.A.; et al. Climate Vulnerability Assessment for Pacific Salmon and Steelhead in the California Current Large Marine Ecosystem. PLoS ONE 2019, 14, e0217711. [Google Scholar] [CrossRef]
- Soulsbury, C.D.; Gray, H.E.; Smith, L.M.; Braithwaite, V.; Cotter, S.C.; Elwood, R.W.; Wilkinson, A.; Collins, L.M. The Welfare and Ethics of Research Involving Wild Animals: A Primer. Methods Ecol. Evol. 2020, 11, 1164–1181. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Parameter | Estimate | SE | t-Value | p-Value | Residual Standard Error |
|---|---|---|---|---|---|
| Control | 0.0031 | ||||
| yf | 13.907 | 0.105 | 132.59 | <0.00001 | |
| y0 | 0.935 | 0.198 | 4.72 | 0.00002 | |
| ln(rate) | −2.653 | 0.034 | −77.73 | <0.00001 | |
| rate | 0.070 | ||||
| Bath only | 0.0011 | ||||
| yf | 6.814 | 0.036 | 188.87 | <0.00001 | |
| y0 | 0.269 | 0.072 | 3.76 | 0.00042 | |
| ln(rate) | −2.622 | 0.024 | −109.71 | <0.00001 | |
| rate | 0.073 | ||||
| Injection only | 0.0013 | ||||
| yf | 3.434 | 0.140 | 24.49 | <0.00001 | |
| y0 | −0.305 | 0.070 | −4.36 | 0.00006 | |
| ln(rate) | −3.360 | 0.088 | −38.01 | <0.00001 | |
| rate | 0.035 | ||||
| Injection and bath | 0.0013 | ||||
| yf | 3.604 | 0.190 | 18.93 | <0.00001 | |
| y0 | −0.046 | 0.068 | −0.68 | 0.502 | |
| ln(rate) | −3.528 | 0.110 | −32.16 | <0.00001 | |
| rate | 0.029 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reed, A.N.; Rowland, F.E.; Krajcik, J.A.; Tillitt, D.E. Thiamine Supplementation Improves Survival and Body Condition of Hatchery-Reared Steelhead (Oncorhynchus mykiss) in Oregon. Vet. Sci. 2023, 10, 156. https://doi.org/10.3390/vetsci10020156
Reed AN, Rowland FE, Krajcik JA, Tillitt DE. Thiamine Supplementation Improves Survival and Body Condition of Hatchery-Reared Steelhead (Oncorhynchus mykiss) in Oregon. Veterinary Sciences. 2023; 10(2):156. https://doi.org/10.3390/vetsci10020156
Chicago/Turabian StyleReed, Aimee N., Freya E. Rowland, Jennifer A. Krajcik, and Donald E. Tillitt. 2023. "Thiamine Supplementation Improves Survival and Body Condition of Hatchery-Reared Steelhead (Oncorhynchus mykiss) in Oregon" Veterinary Sciences 10, no. 2: 156. https://doi.org/10.3390/vetsci10020156